カイコの性は雌に特異的な単一のpiRNAが決定する
木内隆史・勝間 進
(東京大学大学院農学生命科学研究科 生産・環境生物学専攻昆虫遺伝研究室)
email:木内隆史,勝間 進
DOI: 10.7875/first.author.2014.073
A single female-specific piRNA is the primary determiner of sex in the silkworm.
Takashi Kiuchi, Hikaru Koga, Munetaka Kawamoto, Keisuke Shoji, Hiroki Sakai, Yuji Arai, Genki Ishihara, Shinpei Kawaoka, Sumio Sugano, Toru Shimada, Yutaka Suzuki, Masataka G. Suzuki, Susumu Katsuma
Nature, 509, 633-636 (2014)
カイコの性染色体はWZ型であり,1933年,W染色体の存在が雌を決定することが明らかになっている.そのため,W染色体には雌性決定遺伝子が存在すると考えられてきたが,その実体は80年以上も不明であった.今回,筆者らは,初期胚において雌雄のトランスクリプトームを比較することによりW染色体から産生される雌に特異的な転写産物を同定し,それがpiRNAの前駆体であることをつきとめた.また,この転写産物から産生される雌に特異的なpiRNAの標的は,Z染色体に存在する雄化遺伝子であることが示された.雄ではこの遺伝子のはたらきにより雄化が誘導されるのに対し,雌では雄化遺伝子が雌に特異的なpiRNAとカイコのPIWIタンパク質であるSiwiとの複合体のはたらきにより切断されることで雌化が誘導されていた.さらに,この雄化遺伝子は雄性の決定に関与するだけでなく,長年にわたりチョウ目の昆虫において存否が議論されていた遺伝子量の補正にもかかわることが明らかになった.この研究の成果により,80年以上も追い求められてきたカイコの雌性決定遺伝子の実体は,たった29塩基の小分子RNAをコードする遺伝子であり,カイコはこれまで発見された性決定機構とは一線を画すユニークな機構を採用していることがわかった.
生物の性決定機構は多様である.ヒトを含む哺乳類やショウジョウバエにおける性決定機構についてはそれに関与する因子やカスケードの多くが明らかにされてきたが,カイコを含むチョウ目の昆虫においては転写因子をコードするdoublesex(dsx)遺伝子が性決定カスケードの最下流で機能することがわかっているだけで,その上流についてはほとんど明らかにされていなかった.カイコの性染色体はWZ型であり,1933年,W染色体の存在が雌性を決定することが報告されている1).このことから,W染色体には仮想の雌性決定遺伝子であるFeminizer(Fem)遺伝子が存在すると考えられ,長年にわたりその実体が探し求められてきた.しかし,カイコのW染色体はトランスポゾンが入れ子状に配置された構造をしており2),全領域にわたる塩基配列の決定がむずかしいこと,また,雌では組換えが起こらないため順遺伝学的なアプローチが行えないことから,その同定は困難をきわめていた.2011年,筆者らは,W染色体から産生される転写産物としてはじめて,小分子RNAのひとつであるpiRNA(PIWI-interacting RNA)を同定した3).そして,当時から,この雌に多く存在するpiRNAが性の決定に関与するという仮説をたてていた.
カイコの性は発生の初期に決定されると考えられている.雌雄が分化する発生ステージを明瞭にするため,産卵ののちの卵を経時的にサンプリングし,昆虫の性決定において主要な役割を担うdsx遺伝子のカイコにおけるオルソログのスプライシングパターンの変化を追跡した.カイコのdsx遺伝子(Bmdsx遺伝子)もほかの昆虫と同様に,雌雄で異なる選択的スプライシングをうけることにより個体の雌雄を分化させる4-6).初期胚の雌雄を外見から区別することは不可能であるため,W染色体に特異的な3つのRAPDマーカー7) を用いることにより卵ひとつひとつの雌雄を鑑別する方法を開発し,雌雄別にRNAを調製したうえでRT-PCR法により解析した.Bmdsx mRNAは産卵ののち15時間では雌雄ともに雌に特異的なスプライシングを示したが,産卵ののち21時間から24時間にかけて雄では雄に特異的なスプライシングを示しはじめた.すなわち,W染色体からの雌化シグナルは産卵ののち21時間よりまえにすでに伝達されていると推測された.
そこで,産卵ののち15時間から24時間の雌雄別のRNAをRNA-Seq法により解析し,雌雄の胚におけるトランスクリプトームを詳細に比較した.その結果,調べたどの時間帯においてもつねに雌において発現量の高い転写産物を1つ見い出した.この転写産物の配列は全ゲノムが解読されている雄のカイコのゲノムには存在しないこと,雌のゲノムDNAあるいはcDNAを鋳型に用いた場合のみPCR法により増幅産物が確認されたことから,この転写産物はW染色体から産生されると考えられた.しかし,この雌に特異的な転写産物は既知の配列との相同性がまったくなく,さらに,機能をもつタンパク質をコードしているとも考えられなかった.そこで,この転写産物に対し,以前に構築した初期胚や卵巣のpiRNAライブラリー3,8,9) をマッピングしたところ,この転写産物から29塩基の単一のpiRNAが産生されていることがわかった.興味深いことに,この雌に特異的なpiRNAは産卵ののち15時間から蓄積しはじめ18時間から21時間にかけて急激に増加しており,性分化の生じる時期との強い相関がみられた.
雌に特異的な転写産物から産生されるpiRNAの性決定への関与について調べるため,このpiRNAに相補な配列をもつインヒビターRNAを合成し初期胚に注入した.その結果,インヒビターRNAを注入した雌の胚においてBmdsx mRNAの雄に特異的なスプライシングが確認され,雌に特異的なpiRNAの性決定への関与が裏づけられた.piRNAはその名のとおりPIWIタンパク質と複合体を形成することにより機能する.カイコのPIWIをコードするSiwi遺伝子10,11) をRNAi法によりノックダウンしたところ,雌の胚においてBmdsx mRNAの雄に特異的なスプライシングが確認された.さらに,この雌に特異的なpiRNAはSiwiと複合体を形成したことから,この複合体が雌化に寄与していると考えられた.以上の結果から,このW染色体から産生される転写産物をコードする遺伝子こそが,仮想の雌性決定遺伝子であるFem遺伝子であると結論し,その転写産物から産生される29塩基の雌に特異的なpiRNAをFem piRNAと命名した.
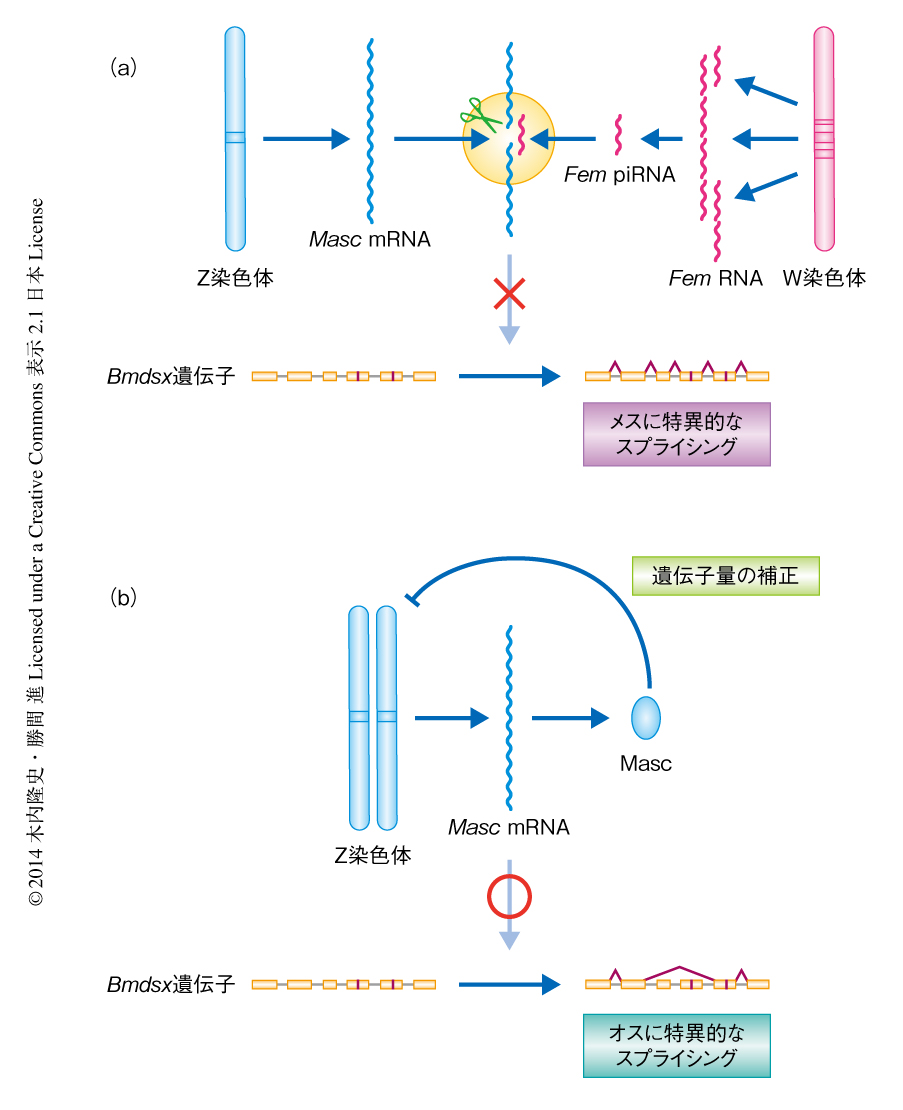
Fem piRNAは雌化をどのように誘導しているのだろうか.piRNAはその配列を利用して1本鎖のRNAを切断する活性をもつPIWIを標的となるRNAへと誘導し,標的であるトランスポゾンなどを切断する8,12,13).Fem piRNAの相補配列をカイコのゲノムにおいて探索したところ,唯一,Z染色体に存在するタンパク質をコードする遺伝子のエキソン部分との相同性が確認された.この遺伝子はCCCHドメインをタンデムに2つもつジンクフィンガータンパク質をコードしていた.系統解析の結果,このタンパク質はチョウ目の昆虫でのみオルソログの存在が認められた.piRNAとPIWIとの複合体は標的となる配列をpiRNAの5’末端から10番目と11番目の塩基のあいだで切断することが知られている12,13).改良RACE法14) を用いた切断部位の同定により,Fem piRNAがこのZ染色体に存在する遺伝子から転写されるmRNAを予想された部位で切断していることが明らかになった.このことから,このZ染色体の遺伝子は雄化遺伝子であり,雌においてはW染色体から産生されるFem piRNAがSiwiと複合体を形成し,この雄化遺伝子の転写産物を切断することにより雌化を誘導していることが示唆された(図1).この遺伝子はMasculinizer(Masc)遺伝子と命名された.
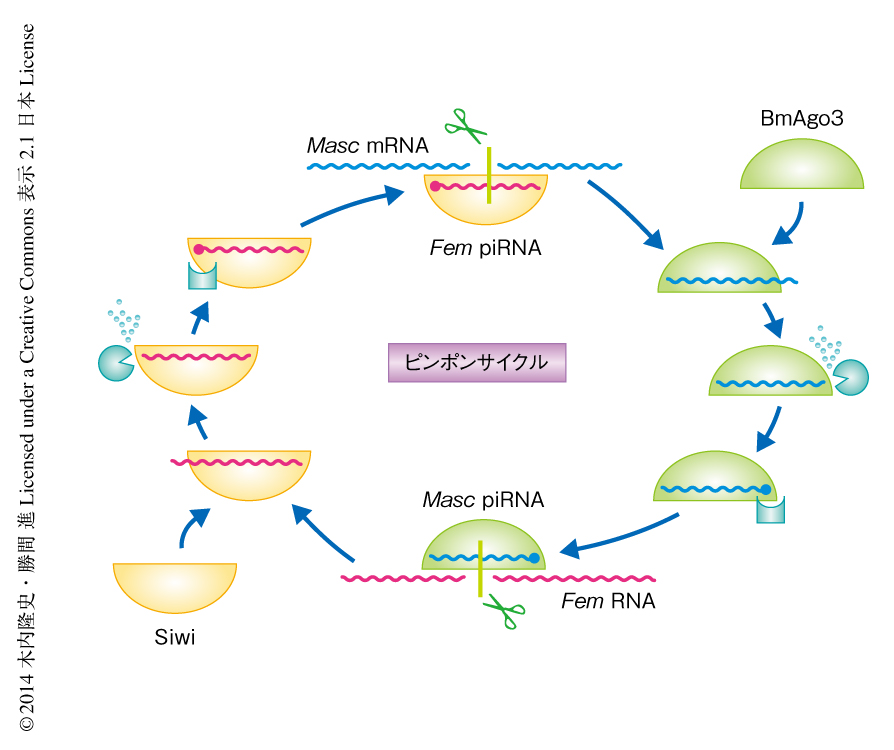
カイコではSiwiとBmAgo3という2つの異なるPIWIを介したピンポンサイクルをへてpiRNAが生合成されるため10-13),Fem piRNAの産生にはFem piRNAと10塩基のオーバーラップをもつpiRNAの存在が必要である.以前に構築したpiRNAライブラリー3,8,9) のなかから,Fem piRNAと10塩基のオーバーラップをもつpiRNAが発見され,それがMasc遺伝子から産生されることがわかった.このMasc遺伝子から産生されるFem piRNAのパートナーをMasc piRNAと命名した.Fem piRNAはSiwiに多く結合していたが,Masc piRNAはBmAgo3に多く結合していたことから,これらpiRNAの生合成のモデルが想定された(図2).これは,非コードRNAとタンパク質をコードする遺伝子とのあいだにピンポンサイクルが存在することを実験的に示したはじめての例であった.
Mascの雄化能を証明するため,初期胚においてRNAi法によるノックダウンを行った.Masc遺伝子に対するsiRNAを注入した雄の胚では雌の胚と同じ程度までMasc遺伝子の発現が抑制されており,Bmdsx遺伝子の雌に特異的なスプライシングが確認された.さらに,Masc遺伝子のsiRNAの注入は雄の胚でのみ孵化を阻害した.Masc遺伝子のノックダウンによる影響を調べるため,Masc遺伝子のsiRNAを注入した胚をRNA-Seq法により解析した結果,雄の胚においてMasc遺伝子のノックダウンにより発現の変化する転写産物の多くはZ染色体にコードされており,発現が亢進するものがほとんどであった.すなわち,Mascは雄が2本もつZ染色体における遺伝子の発現をグローバルに負に制御しており,遺伝子量の補正を担っていると考えられた(図1).チョウ目の昆虫における遺伝子量の補正については長期にわたりその存否が議論されてきたが,少なくとも,初期胚においてはじめてその存在が証明された.
今回,Fem piRNAの発見を契機に,その標的として雄化遺伝子であるMasc遺伝子が同定され,さらに,結論があいまいであったチョウ目の昆虫における遺伝子量の補正の存在も示された.チョウ目昆虫のモデルであるカイコの性決定カスケードの最上流が明らかにされたことにより,その下流の因子やほかのチョウ目昆虫における性決定カスケードもそう遠くない将来に明らかにされるだろう.とくに,W染色体を欠くZ0/ZZ型のチョウ目昆虫の性決定様式の解明は,W染色体の進化を語るうえでも興味深いテーマであると考えている.また,Mascからの性決定カスケードの遮断がひき起こす雄に特異的な胚の致死は,チョウ目昆虫において観察される共生細菌Wolbachiaによる“雄殺し”と類似した現象であり15),共生細菌による性操作の標的がMasc遺伝子やその下流の因子であることを示唆している.今後,これらの因子を標的としたアプローチにより“雄殺し”の分子機構が解明されるかもしれない.
チョウ目の昆虫は主要な農業害虫を多く含んでいる.また,カイコは遺伝子組換え技術を利用した有用物質の生産が期待される昆虫である.この研究の成果が,性操作を利用したチョウ目害虫の防除法の開発や,カイコは雄の生産性のほうが高いことから限性系統の作出につながることを期待している.
80年以上もその実体がつかめなかったW染色体の雌性決定遺伝子は,たった29塩基の小さなRNAをコードするものであった.長年にわたり雌性決定遺伝子が発見できなかった理由も合点がいく.このトランスポゾンに埋もれた“宝石”と,生物らしい“美しい”性決定機構を発見できたのは,筆者らの研究チームにさまざまな人材がバランスよくそろっていたからだと感じている.それにくわえ,カイコの性決定機構の解明に挑んできた多くの研究者たちの知見の蓄積が必要不可欠であったことも,まちがいない事実であろう.
略歴:2008年 東京大学大学院新領域創成科学研究科博士課程 修了,同年 同 博士研究員,2009年 東京大学大学院農学生命科学研究科 博士研究員を経て,2011年より同 特任助教(現 助教).
抱負:昆虫の生命現象を分子レベルで理解し,応用研究に生かしたい.
勝間 進(Susumu Katsuma)
東京大学大学院農学生命科学研究科 准教授.
© 2014 木内隆史・勝間 進 Licensed under CC 表示 2.1 日本
(東京大学大学院農学生命科学研究科 生産・環境生物学専攻昆虫遺伝研究室)
email:木内隆史,勝間 進
DOI: 10.7875/first.author.2014.073
A single female-specific piRNA is the primary determiner of sex in the silkworm.
Takashi Kiuchi, Hikaru Koga, Munetaka Kawamoto, Keisuke Shoji, Hiroki Sakai, Yuji Arai, Genki Ishihara, Shinpei Kawaoka, Sumio Sugano, Toru Shimada, Yutaka Suzuki, Masataka G. Suzuki, Susumu Katsuma
Nature, 509, 633-636 (2014)
要 約
カイコの性染色体はWZ型であり,1933年,W染色体の存在が雌を決定することが明らかになっている.そのため,W染色体には雌性決定遺伝子が存在すると考えられてきたが,その実体は80年以上も不明であった.今回,筆者らは,初期胚において雌雄のトランスクリプトームを比較することによりW染色体から産生される雌に特異的な転写産物を同定し,それがpiRNAの前駆体であることをつきとめた.また,この転写産物から産生される雌に特異的なpiRNAの標的は,Z染色体に存在する雄化遺伝子であることが示された.雄ではこの遺伝子のはたらきにより雄化が誘導されるのに対し,雌では雄化遺伝子が雌に特異的なpiRNAとカイコのPIWIタンパク質であるSiwiとの複合体のはたらきにより切断されることで雌化が誘導されていた.さらに,この雄化遺伝子は雄性の決定に関与するだけでなく,長年にわたりチョウ目の昆虫において存否が議論されていた遺伝子量の補正にもかかわることが明らかになった.この研究の成果により,80年以上も追い求められてきたカイコの雌性決定遺伝子の実体は,たった29塩基の小分子RNAをコードする遺伝子であり,カイコはこれまで発見された性決定機構とは一線を画すユニークな機構を採用していることがわかった.
はじめに
生物の性決定機構は多様である.ヒトを含む哺乳類やショウジョウバエにおける性決定機構についてはそれに関与する因子やカスケードの多くが明らかにされてきたが,カイコを含むチョウ目の昆虫においては転写因子をコードするdoublesex(dsx)遺伝子が性決定カスケードの最下流で機能することがわかっているだけで,その上流についてはほとんど明らかにされていなかった.カイコの性染色体はWZ型であり,1933年,W染色体の存在が雌性を決定することが報告されている1).このことから,W染色体には仮想の雌性決定遺伝子であるFeminizer(Fem)遺伝子が存在すると考えられ,長年にわたりその実体が探し求められてきた.しかし,カイコのW染色体はトランスポゾンが入れ子状に配置された構造をしており2),全領域にわたる塩基配列の決定がむずかしいこと,また,雌では組換えが起こらないため順遺伝学的なアプローチが行えないことから,その同定は困難をきわめていた.2011年,筆者らは,W染色体から産生される転写産物としてはじめて,小分子RNAのひとつであるpiRNA(PIWI-interacting RNA)を同定した3).そして,当時から,この雌に多く存在するpiRNAが性の決定に関与するという仮説をたてていた.
1.W染色体から産生される雌に特異的な転写産物はpiRNAの前駆体である
カイコの性は発生の初期に決定されると考えられている.雌雄が分化する発生ステージを明瞭にするため,産卵ののちの卵を経時的にサンプリングし,昆虫の性決定において主要な役割を担うdsx遺伝子のカイコにおけるオルソログのスプライシングパターンの変化を追跡した.カイコのdsx遺伝子(Bmdsx遺伝子)もほかの昆虫と同様に,雌雄で異なる選択的スプライシングをうけることにより個体の雌雄を分化させる4-6).初期胚の雌雄を外見から区別することは不可能であるため,W染色体に特異的な3つのRAPDマーカー7) を用いることにより卵ひとつひとつの雌雄を鑑別する方法を開発し,雌雄別にRNAを調製したうえでRT-PCR法により解析した.Bmdsx mRNAは産卵ののち15時間では雌雄ともに雌に特異的なスプライシングを示したが,産卵ののち21時間から24時間にかけて雄では雄に特異的なスプライシングを示しはじめた.すなわち,W染色体からの雌化シグナルは産卵ののち21時間よりまえにすでに伝達されていると推測された.
そこで,産卵ののち15時間から24時間の雌雄別のRNAをRNA-Seq法により解析し,雌雄の胚におけるトランスクリプトームを詳細に比較した.その結果,調べたどの時間帯においてもつねに雌において発現量の高い転写産物を1つ見い出した.この転写産物の配列は全ゲノムが解読されている雄のカイコのゲノムには存在しないこと,雌のゲノムDNAあるいはcDNAを鋳型に用いた場合のみPCR法により増幅産物が確認されたことから,この転写産物はW染色体から産生されると考えられた.しかし,この雌に特異的な転写産物は既知の配列との相同性がまったくなく,さらに,機能をもつタンパク質をコードしているとも考えられなかった.そこで,この転写産物に対し,以前に構築した初期胚や卵巣のpiRNAライブラリー3,8,9) をマッピングしたところ,この転写産物から29塩基の単一のpiRNAが産生されていることがわかった.興味深いことに,この雌に特異的なpiRNAは産卵ののち15時間から蓄積しはじめ18時間から21時間にかけて急激に増加しており,性分化の生じる時期との強い相関がみられた.
2.雌に特異的なpiRNAは雌化に必要である
雌に特異的な転写産物から産生されるpiRNAの性決定への関与について調べるため,このpiRNAに相補な配列をもつインヒビターRNAを合成し初期胚に注入した.その結果,インヒビターRNAを注入した雌の胚においてBmdsx mRNAの雄に特異的なスプライシングが確認され,雌に特異的なpiRNAの性決定への関与が裏づけられた.piRNAはその名のとおりPIWIタンパク質と複合体を形成することにより機能する.カイコのPIWIをコードするSiwi遺伝子10,11) をRNAi法によりノックダウンしたところ,雌の胚においてBmdsx mRNAの雄に特異的なスプライシングが確認された.さらに,この雌に特異的なpiRNAはSiwiと複合体を形成したことから,この複合体が雌化に寄与していると考えられた.以上の結果から,このW染色体から産生される転写産物をコードする遺伝子こそが,仮想の雌性決定遺伝子であるFem遺伝子であると結論し,その転写産物から産生される29塩基の雌に特異的なpiRNAをFem piRNAと命名した.
3.Fem piRNAはZ染色体のタンパク質をコードする遺伝子から転写されるmRNAの切断に関与する
Fem piRNAは雌化をどのように誘導しているのだろうか.piRNAはその配列を利用して1本鎖のRNAを切断する活性をもつPIWIを標的となるRNAへと誘導し,標的であるトランスポゾンなどを切断する8,12,13).Fem piRNAの相補配列をカイコのゲノムにおいて探索したところ,唯一,Z染色体に存在するタンパク質をコードする遺伝子のエキソン部分との相同性が確認された.この遺伝子はCCCHドメインをタンデムに2つもつジンクフィンガータンパク質をコードしていた.系統解析の結果,このタンパク質はチョウ目の昆虫でのみオルソログの存在が認められた.piRNAとPIWIとの複合体は標的となる配列をpiRNAの5’末端から10番目と11番目の塩基のあいだで切断することが知られている12,13).改良RACE法14) を用いた切断部位の同定により,Fem piRNAがこのZ染色体に存在する遺伝子から転写されるmRNAを予想された部位で切断していることが明らかになった.このことから,このZ染色体の遺伝子は雄化遺伝子であり,雌においてはW染色体から産生されるFem piRNAがSiwiと複合体を形成し,この雄化遺伝子の転写産物を切断することにより雌化を誘導していることが示唆された(図1).この遺伝子はMasculinizer(Masc)遺伝子と命名された.
カイコではSiwiとBmAgo3という2つの異なるPIWIを介したピンポンサイクルをへてpiRNAが生合成されるため10-13),Fem piRNAの産生にはFem piRNAと10塩基のオーバーラップをもつpiRNAの存在が必要である.以前に構築したpiRNAライブラリー3,8,9) のなかから,Fem piRNAと10塩基のオーバーラップをもつpiRNAが発見され,それがMasc遺伝子から産生されることがわかった.このMasc遺伝子から産生されるFem piRNAのパートナーをMasc piRNAと命名した.Fem piRNAはSiwiに多く結合していたが,Masc piRNAはBmAgo3に多く結合していたことから,これらpiRNAの生合成のモデルが想定された(図2).これは,非コードRNAとタンパク質をコードする遺伝子とのあいだにピンポンサイクルが存在することを実験的に示したはじめての例であった.
4.Mascは性決定および遺伝子量の補正の両方を制御する
Mascの雄化能を証明するため,初期胚においてRNAi法によるノックダウンを行った.Masc遺伝子に対するsiRNAを注入した雄の胚では雌の胚と同じ程度までMasc遺伝子の発現が抑制されており,Bmdsx遺伝子の雌に特異的なスプライシングが確認された.さらに,Masc遺伝子のsiRNAの注入は雄の胚でのみ孵化を阻害した.Masc遺伝子のノックダウンによる影響を調べるため,Masc遺伝子のsiRNAを注入した胚をRNA-Seq法により解析した結果,雄の胚においてMasc遺伝子のノックダウンにより発現の変化する転写産物の多くはZ染色体にコードされており,発現が亢進するものがほとんどであった.すなわち,Mascは雄が2本もつZ染色体における遺伝子の発現をグローバルに負に制御しており,遺伝子量の補正を担っていると考えられた(図1).チョウ目の昆虫における遺伝子量の補正については長期にわたりその存否が議論されてきたが,少なくとも,初期胚においてはじめてその存在が証明された.
おわりに
今回,Fem piRNAの発見を契機に,その標的として雄化遺伝子であるMasc遺伝子が同定され,さらに,結論があいまいであったチョウ目の昆虫における遺伝子量の補正の存在も示された.チョウ目昆虫のモデルであるカイコの性決定カスケードの最上流が明らかにされたことにより,その下流の因子やほかのチョウ目昆虫における性決定カスケードもそう遠くない将来に明らかにされるだろう.とくに,W染色体を欠くZ0/ZZ型のチョウ目昆虫の性決定様式の解明は,W染色体の進化を語るうえでも興味深いテーマであると考えている.また,Mascからの性決定カスケードの遮断がひき起こす雄に特異的な胚の致死は,チョウ目昆虫において観察される共生細菌Wolbachiaによる“雄殺し”と類似した現象であり15),共生細菌による性操作の標的がMasc遺伝子やその下流の因子であることを示唆している.今後,これらの因子を標的としたアプローチにより“雄殺し”の分子機構が解明されるかもしれない.
チョウ目の昆虫は主要な農業害虫を多く含んでいる.また,カイコは遺伝子組換え技術を利用した有用物質の生産が期待される昆虫である.この研究の成果が,性操作を利用したチョウ目害虫の防除法の開発や,カイコは雄の生産性のほうが高いことから限性系統の作出につながることを期待している.
80年以上もその実体がつかめなかったW染色体の雌性決定遺伝子は,たった29塩基の小さなRNAをコードするものであった.長年にわたり雌性決定遺伝子が発見できなかった理由も合点がいく.このトランスポゾンに埋もれた“宝石”と,生物らしい“美しい”性決定機構を発見できたのは,筆者らの研究チームにさまざまな人材がバランスよくそろっていたからだと感じている.それにくわえ,カイコの性決定機構の解明に挑んできた多くの研究者たちの知見の蓄積が必要不可欠であったことも,まちがいない事実であろう.
文 献
- 橋本春雄.: 蚕に於けるW染色体の性決定に対する役割. 遺伝学雑誌, 8, 245-247 (1933)
- Abe, H., Mita, K., Yasukochi, Y. et al.: Retrotransposable elements on the W chromosome of the silkworm, Bombyx mori. Cytogenet. Genome Res., 110, 144-151 (2005)[PubMed]
- Kawaoka, S., Kadota, K., Arai, Y. et al.: The silkworm W chromosome is a source of female-enriched piRNAs. RNA, 17, 2144-2151 (2011)[PubMed]
- Ohbayashi, F., Suzuki, M. G., Mita, K. et al.: A homologue of the Drosophila doublesex gene is transcribed into sex-specific mRNA isoforms in the silkworm, Bombyx mori. Comp. Biochem. Physiol. B Biochem. Mol. Biol., 128, 145-158 (2001)[PubMed]
- Suzuki, M. G., Funaguma, S., Kanda, T. et al.: Analysis of the biological functions of a doublesex homologue in Bombyx mori. Dev. Genes Evol., 213, 345-354 (2003)[PubMed]
- Suzuki, M. G., Funaguma, S., Kanda, T. et al.: Role of the male BmDSX protein in the sexual differentiation of Bombyx mori. Evol. Dev., 7, 58-68 (2005)[PubMed]
- Abe, H., Seki, M., Ohbayashi, F. et al.: Partial deletions of the W chromosome due to reciprocal translocation in the silkworm Bombyx mori. Insect Mol. Biol., 14, 339-352 (2005)[PubMed]
- Kawaoka, S., Hayashi, N., Katsuma, S. et al.: Bombyx small RNAs: genomic defense system against transposons in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol., 38, 1058-1065 (2008)[PubMed]
- Kawaoka, S., Arai, Y., Kadota, K. et al.: Zygotic amplification of secondary piRNAs during silkworm embryogenesis. RNA, 17, 1401-1407 (2011)[PubMed]
- Kawaoka, S. Hayashi, N., Suzuki, Y. et al.: The Bombyx ovary-derived cell line endogenously expresses PIWI/PIWI-interacting RNA complexes. RNA, 15, 1258-1264 (2009)[PubMed]
- Kawaoka, S., Minami, K., Katsuma, S. et al.: Developmentally synchronized expression of two Bombyx mori Piwi subfamily genes, SIWIand BmAGO3 in germ-line cells. Biochem. Biophys. Res. Commun., 367, 755-760 (2008)[PubMed]
- Brennecke, J., Aravin, A. A., Stark, A. et al.: Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell, 128, 1089-1103 (2007)[PubMed]
- Gunawardane, L. S., Saito, K., Nishida, K. M. et al.: A slicer-mediated mechanism for repeat-associated siRNA 5’ end formation in Drosophila. Science, 315, 1587-1590 (2007)[PubMed]
- Watanabe, T., Tomizawa, S., Mitsuya, K. et al.: Role of piRNAs and noncoding RNA in de novo DNA methylation of the imprinted mouse Rasgrf1 locus. Science, 332, 848-852 (2011)[PubMed] [新着論文レビュー]
- Sugimoto, T. N. & Ishikawa, Y.: A male-killing Wolbachia carries a feminizing factor and is associated with degradation of the sex-determining system of its host. Biol. Lett., 8, 412-415 (2012)[PubMed]
著者プロフィール
略歴:2008年 東京大学大学院新領域創成科学研究科博士課程 修了,同年 同 博士研究員,2009年 東京大学大学院農学生命科学研究科 博士研究員を経て,2011年より同 特任助教(現 助教).
抱負:昆虫の生命現象を分子レベルで理解し,応用研究に生かしたい.
勝間 進(Susumu Katsuma)
東京大学大学院農学生命科学研究科 准教授.
© 2014 木内隆史・勝間 進 Licensed under CC 表示 2.1 日本
