やさしく押された圧感覚を担うMerkel細胞による機械刺激の受容および神経への伝達
仲谷正史・馬場欣哉・Srdjan Maksimovic・Ellen A. Lumpkin
(米国Columbia大学Department of Dermatology)
email:仲谷正史,馬場欣哉
DOI: 10.7875/first.author.2014.062
Epidermal Merkel cells are mechanosensory cells that tune mammalian touch receptors.
Srdjan Maksimovic, Masashi Nakatani, Yoshichika Baba, Aislyn M. Nelson, Kara L. Marshall, Scott A. Wellnitz, Pervez Firozi, Seung-Hyun Woo, Sanjeev Ranade, Ardem Patapoutian, Ellen A. Lumpkin
Nature, 509, 617-621 (2014)
近年,触覚の細胞生理学的あるいは分子生物学的な研究がさかんに行われているが,その一方で,いまだに触覚における基本的な問題は十分に明らかになってはいない.そのひとつとして,長いあいだ機械受容器と考えられてきているMerkel細胞が,本当に機械受容器として外界からの刺激を受容しこれを末梢神経に伝達しているのかという問題については,これを解決する直接的な証拠は得られていなかった.今回は,遺伝子組換え技術と生理学的な手法を用いたMerkel細胞の研究をとおして,この問題を解決した.また,Merkel細胞-神経複合体がそれぞれ機械受容器としての役割をはたし,それらをあわせたシステムが生体触覚センサーとして機能しているとのモデルを提案した.
触覚は五感のなかでもっとも基礎的な感覚であるにもかかわらず,もっとも解明が遅れている.歴史的にみると,機械受容の研究は微生物のもつ機械受容チャネルの解明から進展してきたが1),近年になり,無脊椎動物の体表の機械感覚や哺乳類の皮膚感覚の分子生物学的な研究が急速に進展している2,3).皮膚に入り込む末梢神経それ自体が触覚受容器として機能していることが明らかになり4),また,発現する神経伝達物質受容体の種類により末梢神経がいくつかの種類に分けられることがわかってきている5).
一方,皮膚には神経だけでなく,機械受容器とよばれる,神経の終末において特異な形態をもちほかの表皮細胞や真皮細胞と区別される構造体が報告されている.そのひとつであるMerkel細胞は,1875年に発見され,触覚受容器としての機能が予測されていた.Merkel細胞は神経の終末と隣接していることが知られており,Merkel細胞-神経複合体とよばれている.触盤とよばれるMerkel細胞-神経複合体の密集する有毛部の皮膚を機械刺激すると,接続する求心性神経から遅順応の応答が得られる.Merkel細胞は神経と隣接する部位に神経伝達物質を含む小胞を保持していることが電子顕微鏡による観察で明らかにされたことより,神経の終末とのあいだでなんらかのシグナル伝達が行われていると予測されている6).
これまで,Merkel細胞それ自体が機械受容の特性をもつことが間接的に示されてきた.つまり,Merkel細胞は,浸透圧の変化7),陰圧8),せん断力9) により応答することが,Ca2+イメージング法により示されてきた.しかしながら,Ca2+イメージング法は定性的に生理応答を調べられる一方で,保持電位などを操作できないことから,機械受容チャネルの生物物理的な特性を詳細に調べることはむずかしい.また,本当の意味でMerkel細胞が機械受容器であることを示すためには,Merkel細胞が機械刺激に対し応答するだけでなく,それを神経に伝達できることを示す必要があった.
有毛部の皮膚においてMerkel細胞の量は少なく,表皮を構成する細胞の0.5%しかない.Merkel細胞において特異的に発現する転写因子Atoh1の遺伝子のエンハンサー領域にGFPの遺伝子を導入したトランスジェニックマウス10) を用いて,生体においてMerkel細胞を標識し,この標識を目印としてフローサイトメトリーによりMerkel細胞のみを単離し,数日間にわたり培養した.そして,先行研究と同様の方法11) により,ホールセルパッチクランプ法を用いて機械刺激に対する応答を調べた.その結果,培養したMerkel細胞は単体で機械刺激に対し応答し,保持電位が-70 mVのとき,数μmの機械刺激により内向き電流応答は飽和することが明らかになった.Merkel細胞を機械刺激することにより膜電位は脱分極するという知見12) から,皮膚の大多数をしめる表皮細胞がなくとも,Merkel細胞は単体で機械刺激に対し興奮性の応答を示すと考えられた.ラットの洞毛のin situ標本を用いた実験においても,同様の結果が報告されている13).
Merkel細胞において機械受容を担うチャネルは何であろうか.このチャネルの生物物理学的な特性を知るため,機械刺激にともなう応答をさまざまな固定電位において測定し平衡電位を求めた.その結果,8 mV前後で機械刺激による電流の向きは逆転したことから,非選択性の陽イオンチャネルであることが示唆された.また,細胞の外液にルテニウムレッドやGd3+ 13) が存在すると機械刺激の受容による電流は抑制されることが観察された.以上の結果は,機械刺激受容チャネルのひとつであるPiezoチャネルの性質と類似していた11).実際に,Merkel細胞を定量的PCR法により解析した結果,確かにPiezo1およびPiezo2が発現していること,皮膚の全体と比較してPiezo2がより多く発現していることがわかった.さらに,皮膚のみでPiezo2チャネルをノックアウトしたマウスのMerkel細胞では機械刺激の受容による電流が測定されないことから12),Merkel細胞にはPiezo2チャネルが発現し,その機械受容の特性を担っていることが明らかになった.
Merkel細胞が機械刺激に対し応答することが明らかになったが,機械受容器として機能していることを証明するためには,Merkel細胞が刺激されたのち,接続する遅順応の求心性神経にその情報が伝達されていることを示す必要がある.しかし,この確認実験は容易ではなかった.なぜなら,末梢神経それ自体もまた,機械受容器として機能しているからである.たとえば,後根神経節に由来する神経細胞を機械刺激すると脱分極性の反応を起こすことが報告されている4).そのため,Merkel細胞のみを興奮させ,それが神経細胞に興奮を伝達することを示す必要があった.
この目標を達成するため,光遺伝学の手法を採用した.具体的には,Merkel細胞にチャネルロドプシン2を発現させたトランスジェニックマウスを作製し,刺激光によりMerkel細胞を興奮させることにした.このトランスジェニックマウスの表皮からMerkel細胞を単離して培養し,ホールセルパッチクランプを施したうえで光刺激すると,光刺激に応答した内向き電流が観測された.これは,機械刺激をしたときの応答と類似しており,光刺激により確かにチャネルロドプシン2を発現するMerkel細胞が興奮したことが示された.そこで,このトランスジェニックマウスから皮膚-神経標本を作製した.皮膚-神経標本は生体のマウスから下肢の皮膚と伏在神経との接続を断つことなく取り出すことにより,ex vivoにおいて皮膚を刺激した際の求心性の神経応答の測定が可能となる実験系である.この系を用いてMerkel細胞に対し光刺激したところ,遅順応の求心性神経から活動電位が観測された.この結果から,Merkel細胞は機械受容の特性をもつだけでなく,神経の終末にシグナルを伝達しうることが示された.
以上の知見は,Merkel細胞の存在することが遅順応の神経応答をひき起こすための十分条件であることを示した.一方で,この実験ではMerkel細胞が標準的な遅順応の求心性の神経応答に寄与しているのかどうかについては確認できない.そこで,光刺激によりプロトンポンプとして機能するアーキロドプシンをMerkel細胞に発現させたトランスジェニックマウスを作製した.皮膚-神経標本において機械刺激に対し応答を示しているときにMerkel細胞を光刺激したところ,遅順応の神経応答は抑制されることがわかった.このことは,Merkel細胞が遅順応の神経応答を能動的に変調していることを示した.すなわち,Merkel細胞の存在は神経応答における遅順応性を担保するための必要条件であった.
Merkel細胞は求心性神経へ触覚の情報を伝達する際に,具体的にはどのような役割を担っているのであろうか.このことを詳細に知るため,Merkel細胞において特異的に発現する転写因子Atoh1を表皮のみで欠損させたトランスジェニックマウスを作製した.触覚刺激に対する応答の違いを皮膚-神経標本を用いてAβ感覚神経から細胞外記録して比較したところ,対照となるマウスでは,動的な機械刺激により神経発火が高い頻度でみられた一方,静的な圧刺激に対しては順応の遅い(刺激をあたえているあいだ反応しつづける)神経発火が得られた.一方,表皮に特異的なAtoh1ノックアウトマウスでは,動的な機械刺激に対する神経発火の頻度は減少し,静的な機械刺激の開始から4秒以内に順応する中順応性の反応が観察された.閾値より十分に大きな強度の機械刺激をあたえた場合,表皮に特異的なAtoh1ノックアウトマウスでは81%に中順応性の神経応答が観察されたのに対し,対照となるマウスではわずか3%しかみられなかった.また,Merkel細胞における主要な機械受容チャネルであるPiezo2が欠損したマウスにおいても,同様な中順応性の神経応答が観察されている12).
以上のことから,Merkel細胞は遅順応の神経応答に対し2つの方法により貢献していることが示唆された(図1).ひとつは,Merkel細胞はPiezo2チャネルに依存した機構により,静的な機械刺激に対する応答に寄与する.これはすなわち,Merkel細胞は“やさしく押された”圧感覚をつかさどっていると考えられる.実際に,触覚に対する応答の閾値を求めるvon Frey試験において,強い刺激に対する閾値については野生型マウスとPiezo2ノックアウトマウスとのあいだで有意な差はない一方,弱い刺激に対する閾値についてはPiezo2ノックアウトマウスにおいて有意に上昇していることが確認されている12).もうひとつは,Merkel細胞は皮膚の動的な変形をみのがすことなく頑健に感じとるために必要である.この特徴は動的な刺激に対し高い神経発火の頻度により応答することから導かれる.Merkel細胞が動的な刺激に対しどのように応答しているのか,そのしくみについては今後の検討の余地がある.
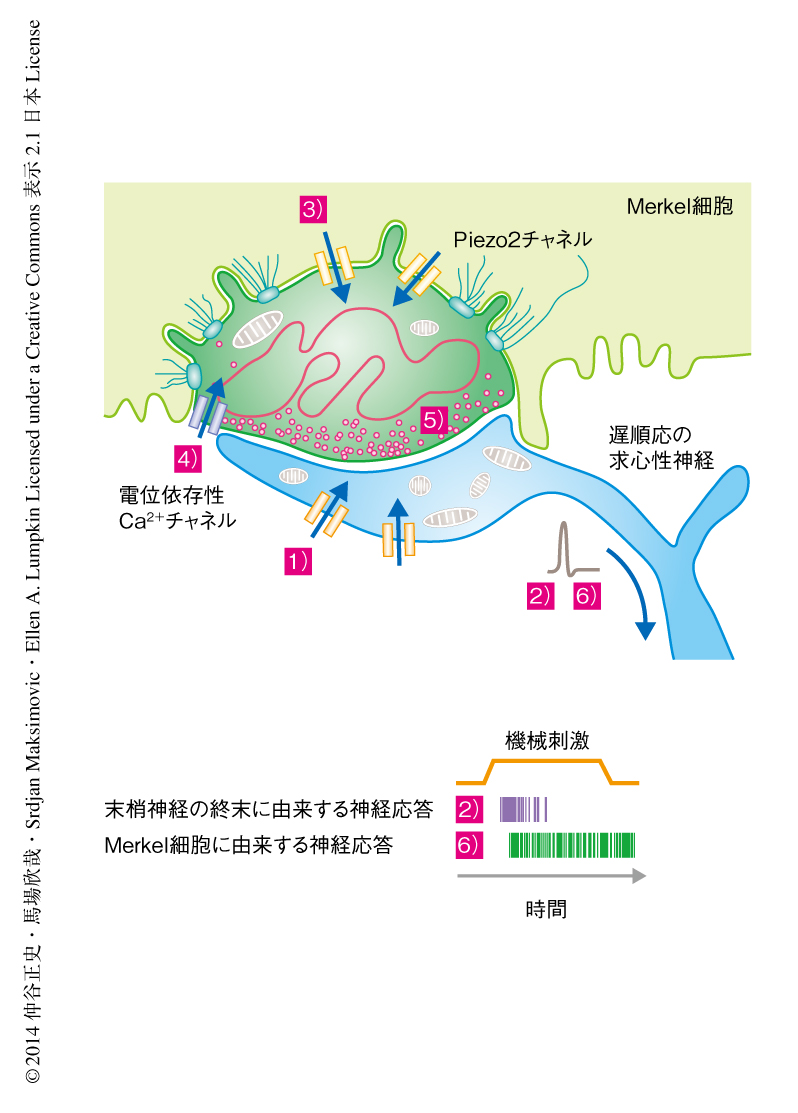
以上の研究により,Merkel細胞が機械刺激に対し興奮性の応答をすること,その興奮を神経に伝達すること,神経応答を積極的に変調する役割のあること,がわかった.また,Merkel細胞における機械受容チャネルとしてPiezo2チャネルが機能しており,これを欠損すると機械受容の特性がみられなくなることもわかった.これらの知見は,Merkel細胞および神経の終末はともに遅順応の神経応答に寄与しているという2受容部位仮説6,14) を支持したうえで,かつ,Merkel細胞はシグナルの変調にかかわることを示唆した(図1).そして,Merkel細胞-神経複合体が機械受容器として相互に協力しながら,これまで多くの研究により報告されてきた,典型的な遅順応の求心性の神経応答をかたちづくることが示唆された.神経の終末が動的な刺激に対し活発に応答するのに対し,Merkel細胞は持続的に押されているときの応答に寄与していることが想定されたことから,Merkel細胞の機械受容器としての役割は“やさしく押された圧感覚”(light touch)にあると考えられた.
まだ2つの興味深い疑問が残されている.ひとつ,Merkel細胞のもつ機械受容チャネルは機械刺激を敏感に検知するため細胞内に何か特別な機構をもっているのだろうか.機械受容チャネルは細胞膜に一様に分布しているのかもしれないし,有毛細胞のステレオシリアのように特定の場所に局在しているのかもしれない15).指先のような無毛部では遅順応の神経応答は粗いテクスチャやエッジに対し活発に応答することが知られているが,Merkel細胞がこのような高い空間解像度を実現するのに寄与していることが予想される.もうひとつ,Merkel細胞から神経の終末へどのようにしてシグナルが伝達されるのかについても明らかではない.先行研究により,神経伝達物質であるATP,グルタミン酸,セロトニンなどがMerkel細胞からのシグナル伝達に寄与すると指摘される一方,その具体的な様式について直接的な証拠は得られていない.これらの問題をひとつひとつ明らかにし,Merkel細胞のつかさどる触知覚の研究に緒を得ることにより,五感の最後のフロンティアである触覚を明らかにする手法の開発とさらなる知見の獲得をめざす考えである.
略歴:2008年 東京大学大学院情報理工学系研究科 修了,同年 資生堂リサーチセンター 研究員を経て,2012年より慶應義塾大学大学院システムデザイン・マネジメント研究科 訪問研究員.
研究テーマ:Merkel細胞のメカノバイオロジー,触覚工学(触覚ディスプレイセンサーの開発),触覚心理学.
関心事:生体が機械刺激を受容して神経に伝達し中枢において知覚される過程の包括的な理解と,その計算論および工学的な応用.
馬場 欣哉(Yoshichika Baba)
略歴:1991年 岡山大学大学院自然科学研究科 修了,同年 北海道大学電子科学研究所 助手,2000年 富山大学工学部 講師,2002年 米国Illinois大学Chicago校Research Assistant Professor,2010年 米国Montana大学Missoula校Research Assistant Professorを経て,2011年より米国Columbia大学Associate Researcher.
研究テーマ:逃避行動の神経行動学.皮膚の接触感覚の信号処理.
関心事:外界を生体内の信号(スパイク列)に変換するときの符号化の方法.また,その信号から状況に適した情報を取り出してくる神経機構のアルゴリズム.
Srdjan Maksimovic
米国Columbia大学Postdoctoral Research Fellow.
Ellen A. Lumpkin
米国Columbia大学Associate Professor.
© 2014 仲谷正史・馬場欣哉・Srdjan Maksimovic・Ellen A. Lumpkin Licensed under CC 表示 2.1 日本
(米国Columbia大学Department of Dermatology)
email:仲谷正史,馬場欣哉
DOI: 10.7875/first.author.2014.062
Epidermal Merkel cells are mechanosensory cells that tune mammalian touch receptors.
Srdjan Maksimovic, Masashi Nakatani, Yoshichika Baba, Aislyn M. Nelson, Kara L. Marshall, Scott A. Wellnitz, Pervez Firozi, Seung-Hyun Woo, Sanjeev Ranade, Ardem Patapoutian, Ellen A. Lumpkin
Nature, 509, 617-621 (2014)
この論文に出現する遺伝子・タンパク質のUniprot ID
要 約
近年,触覚の細胞生理学的あるいは分子生物学的な研究がさかんに行われているが,その一方で,いまだに触覚における基本的な問題は十分に明らかになってはいない.そのひとつとして,長いあいだ機械受容器と考えられてきているMerkel細胞が,本当に機械受容器として外界からの刺激を受容しこれを末梢神経に伝達しているのかという問題については,これを解決する直接的な証拠は得られていなかった.今回は,遺伝子組換え技術と生理学的な手法を用いたMerkel細胞の研究をとおして,この問題を解決した.また,Merkel細胞-神経複合体がそれぞれ機械受容器としての役割をはたし,それらをあわせたシステムが生体触覚センサーとして機能しているとのモデルを提案した.
はじめに
触覚は五感のなかでもっとも基礎的な感覚であるにもかかわらず,もっとも解明が遅れている.歴史的にみると,機械受容の研究は微生物のもつ機械受容チャネルの解明から進展してきたが1),近年になり,無脊椎動物の体表の機械感覚や哺乳類の皮膚感覚の分子生物学的な研究が急速に進展している2,3).皮膚に入り込む末梢神経それ自体が触覚受容器として機能していることが明らかになり4),また,発現する神経伝達物質受容体の種類により末梢神経がいくつかの種類に分けられることがわかってきている5).
一方,皮膚には神経だけでなく,機械受容器とよばれる,神経の終末において特異な形態をもちほかの表皮細胞や真皮細胞と区別される構造体が報告されている.そのひとつであるMerkel細胞は,1875年に発見され,触覚受容器としての機能が予測されていた.Merkel細胞は神経の終末と隣接していることが知られており,Merkel細胞-神経複合体とよばれている.触盤とよばれるMerkel細胞-神経複合体の密集する有毛部の皮膚を機械刺激すると,接続する求心性神経から遅順応の応答が得られる.Merkel細胞は神経と隣接する部位に神経伝達物質を含む小胞を保持していることが電子顕微鏡による観察で明らかにされたことより,神経の終末とのあいだでなんらかのシグナル伝達が行われていると予測されている6).
これまで,Merkel細胞それ自体が機械受容の特性をもつことが間接的に示されてきた.つまり,Merkel細胞は,浸透圧の変化7),陰圧8),せん断力9) により応答することが,Ca2+イメージング法により示されてきた.しかしながら,Ca2+イメージング法は定性的に生理応答を調べられる一方で,保持電位などを操作できないことから,機械受容チャネルの生物物理的な特性を詳細に調べることはむずかしい.また,本当の意味でMerkel細胞が機械受容器であることを示すためには,Merkel細胞が機械刺激に対し応答するだけでなく,それを神経に伝達できることを示す必要があった.
1.Merkel細胞は機械刺激に対し応答する
有毛部の皮膚においてMerkel細胞の量は少なく,表皮を構成する細胞の0.5%しかない.Merkel細胞において特異的に発現する転写因子Atoh1の遺伝子のエンハンサー領域にGFPの遺伝子を導入したトランスジェニックマウス10) を用いて,生体においてMerkel細胞を標識し,この標識を目印としてフローサイトメトリーによりMerkel細胞のみを単離し,数日間にわたり培養した.そして,先行研究と同様の方法11) により,ホールセルパッチクランプ法を用いて機械刺激に対する応答を調べた.その結果,培養したMerkel細胞は単体で機械刺激に対し応答し,保持電位が-70 mVのとき,数μmの機械刺激により内向き電流応答は飽和することが明らかになった.Merkel細胞を機械刺激することにより膜電位は脱分極するという知見12) から,皮膚の大多数をしめる表皮細胞がなくとも,Merkel細胞は単体で機械刺激に対し興奮性の応答を示すと考えられた.ラットの洞毛のin situ標本を用いた実験においても,同様の結果が報告されている13).
Merkel細胞において機械受容を担うチャネルは何であろうか.このチャネルの生物物理学的な特性を知るため,機械刺激にともなう応答をさまざまな固定電位において測定し平衡電位を求めた.その結果,8 mV前後で機械刺激による電流の向きは逆転したことから,非選択性の陽イオンチャネルであることが示唆された.また,細胞の外液にルテニウムレッドやGd3+ 13) が存在すると機械刺激の受容による電流は抑制されることが観察された.以上の結果は,機械刺激受容チャネルのひとつであるPiezoチャネルの性質と類似していた11).実際に,Merkel細胞を定量的PCR法により解析した結果,確かにPiezo1およびPiezo2が発現していること,皮膚の全体と比較してPiezo2がより多く発現していることがわかった.さらに,皮膚のみでPiezo2チャネルをノックアウトしたマウスのMerkel細胞では機械刺激の受容による電流が測定されないことから12),Merkel細胞にはPiezo2チャネルが発現し,その機械受容の特性を担っていることが明らかになった.
2.Merkel細胞は遅順応の求心性神経に触覚の情報を伝達する
Merkel細胞が機械刺激に対し応答することが明らかになったが,機械受容器として機能していることを証明するためには,Merkel細胞が刺激されたのち,接続する遅順応の求心性神経にその情報が伝達されていることを示す必要がある.しかし,この確認実験は容易ではなかった.なぜなら,末梢神経それ自体もまた,機械受容器として機能しているからである.たとえば,後根神経節に由来する神経細胞を機械刺激すると脱分極性の反応を起こすことが報告されている4).そのため,Merkel細胞のみを興奮させ,それが神経細胞に興奮を伝達することを示す必要があった.
この目標を達成するため,光遺伝学の手法を採用した.具体的には,Merkel細胞にチャネルロドプシン2を発現させたトランスジェニックマウスを作製し,刺激光によりMerkel細胞を興奮させることにした.このトランスジェニックマウスの表皮からMerkel細胞を単離して培養し,ホールセルパッチクランプを施したうえで光刺激すると,光刺激に応答した内向き電流が観測された.これは,機械刺激をしたときの応答と類似しており,光刺激により確かにチャネルロドプシン2を発現するMerkel細胞が興奮したことが示された.そこで,このトランスジェニックマウスから皮膚-神経標本を作製した.皮膚-神経標本は生体のマウスから下肢の皮膚と伏在神経との接続を断つことなく取り出すことにより,ex vivoにおいて皮膚を刺激した際の求心性の神経応答の測定が可能となる実験系である.この系を用いてMerkel細胞に対し光刺激したところ,遅順応の求心性神経から活動電位が観測された.この結果から,Merkel細胞は機械受容の特性をもつだけでなく,神経の終末にシグナルを伝達しうることが示された.
以上の知見は,Merkel細胞の存在することが遅順応の神経応答をひき起こすための十分条件であることを示した.一方で,この実験ではMerkel細胞が標準的な遅順応の求心性の神経応答に寄与しているのかどうかについては確認できない.そこで,光刺激によりプロトンポンプとして機能するアーキロドプシンをMerkel細胞に発現させたトランスジェニックマウスを作製した.皮膚-神経標本において機械刺激に対し応答を示しているときにMerkel細胞を光刺激したところ,遅順応の神経応答は抑制されることがわかった.このことは,Merkel細胞が遅順応の神経応答を能動的に変調していることを示した.すなわち,Merkel細胞の存在は神経応答における遅順応性を担保するための必要条件であった.
3.Merkel細胞の有無による遅順応の求心性の神経応答の違い
Merkel細胞は求心性神経へ触覚の情報を伝達する際に,具体的にはどのような役割を担っているのであろうか.このことを詳細に知るため,Merkel細胞において特異的に発現する転写因子Atoh1を表皮のみで欠損させたトランスジェニックマウスを作製した.触覚刺激に対する応答の違いを皮膚-神経標本を用いてAβ感覚神経から細胞外記録して比較したところ,対照となるマウスでは,動的な機械刺激により神経発火が高い頻度でみられた一方,静的な圧刺激に対しては順応の遅い(刺激をあたえているあいだ反応しつづける)神経発火が得られた.一方,表皮に特異的なAtoh1ノックアウトマウスでは,動的な機械刺激に対する神経発火の頻度は減少し,静的な機械刺激の開始から4秒以内に順応する中順応性の反応が観察された.閾値より十分に大きな強度の機械刺激をあたえた場合,表皮に特異的なAtoh1ノックアウトマウスでは81%に中順応性の神経応答が観察されたのに対し,対照となるマウスではわずか3%しかみられなかった.また,Merkel細胞における主要な機械受容チャネルであるPiezo2が欠損したマウスにおいても,同様な中順応性の神経応答が観察されている12).
以上のことから,Merkel細胞は遅順応の神経応答に対し2つの方法により貢献していることが示唆された(図1).ひとつは,Merkel細胞はPiezo2チャネルに依存した機構により,静的な機械刺激に対する応答に寄与する.これはすなわち,Merkel細胞は“やさしく押された”圧感覚をつかさどっていると考えられる.実際に,触覚に対する応答の閾値を求めるvon Frey試験において,強い刺激に対する閾値については野生型マウスとPiezo2ノックアウトマウスとのあいだで有意な差はない一方,弱い刺激に対する閾値についてはPiezo2ノックアウトマウスにおいて有意に上昇していることが確認されている12).もうひとつは,Merkel細胞は皮膚の動的な変形をみのがすことなく頑健に感じとるために必要である.この特徴は動的な刺激に対し高い神経発火の頻度により応答することから導かれる.Merkel細胞が動的な刺激に対しどのように応答しているのか,そのしくみについては今後の検討の余地がある.
おわりに
以上の研究により,Merkel細胞が機械刺激に対し興奮性の応答をすること,その興奮を神経に伝達すること,神経応答を積極的に変調する役割のあること,がわかった.また,Merkel細胞における機械受容チャネルとしてPiezo2チャネルが機能しており,これを欠損すると機械受容の特性がみられなくなることもわかった.これらの知見は,Merkel細胞および神経の終末はともに遅順応の神経応答に寄与しているという2受容部位仮説6,14) を支持したうえで,かつ,Merkel細胞はシグナルの変調にかかわることを示唆した(図1).そして,Merkel細胞-神経複合体が機械受容器として相互に協力しながら,これまで多くの研究により報告されてきた,典型的な遅順応の求心性の神経応答をかたちづくることが示唆された.神経の終末が動的な刺激に対し活発に応答するのに対し,Merkel細胞は持続的に押されているときの応答に寄与していることが想定されたことから,Merkel細胞の機械受容器としての役割は“やさしく押された圧感覚”(light touch)にあると考えられた.
まだ2つの興味深い疑問が残されている.ひとつ,Merkel細胞のもつ機械受容チャネルは機械刺激を敏感に検知するため細胞内に何か特別な機構をもっているのだろうか.機械受容チャネルは細胞膜に一様に分布しているのかもしれないし,有毛細胞のステレオシリアのように特定の場所に局在しているのかもしれない15).指先のような無毛部では遅順応の神経応答は粗いテクスチャやエッジに対し活発に応答することが知られているが,Merkel細胞がこのような高い空間解像度を実現するのに寄与していることが予想される.もうひとつ,Merkel細胞から神経の終末へどのようにしてシグナルが伝達されるのかについても明らかではない.先行研究により,神経伝達物質であるATP,グルタミン酸,セロトニンなどがMerkel細胞からのシグナル伝達に寄与すると指摘される一方,その具体的な様式について直接的な証拠は得られていない.これらの問題をひとつひとつ明らかにし,Merkel細胞のつかさどる触知覚の研究に緒を得ることにより,五感の最後のフロンティアである触覚を明らかにする手法の開発とさらなる知見の獲得をめざす考えである.
文 献
- Kung, C., Martinac, B. & Sukharev, S.: Mechanosensitive channels in microbes. Annu. Rev. Microbiol., 64, 313-329 (2010)[PubMed]
- Lumpkin, E. A. & Caterina, M. J.: Mechanisms of sensory transduction in the skin. Nature, 445, 858-865 (2007)[PubMed]
- Chalfie, M.: Neurosensory mechanotransduction. Nat. Rev. Mol. Cell Biol., 10, 44-52 (2009)[PubMed]
- Hu, J. & Lewin, G. R.: Mechanosensitive currents in the neurites of cultured mouse sensory neurones. J. Physiol., 577, 815-828 (2006)[PubMed]
- Abraira, V. E. & Ginty, D. D.: The sensory neurons of touch. Neuron, 79, 618-639 (2013)[PubMed]
- Iggo, A. & Muir, A. R.: The structure and function of a slowly adapting touch corpuscle in hairy skin. J. Physiol., 200, 763-796 (1969)[PubMed]
- Haeberle, H., Bryan, L. A., Vadakkan, T. J. et al.: Swelling-activated Ca2+ channels trigger Ca2+ signals in Merkel cells. PLoS One, 3, e1750 (2008)[PubMed]
- Boulais, N., Pennec, J. P., Lebonvallet, N. et al.: Rat Merkel cells are mechanoreceptors and osmoreceptors. PLoS One, 4, e7759 (2009)[PubMed]
- Cha, M., Ling, J., Xu, G. Y., et al.: Shear mechanical force induces an increase of intracellular Ca2+ in cultured Merkel cells prepared from rat vibrissal hair follicles. J. Neurophysiol., 106, 460-469 (2011)[PubMed]
- Lumpkin, E. A., Collisson, T., Parab, P. et al.: Math1-driven GFP expression in the developing nervous system of transgenic mice. Gene Expr. Patterns, 3, 389-395 (2003)[PubMed]
- Coste, B., Mathur, J., Schmidt, M. et al.: Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science, 330, 55-60 (2010)[PubMed]
- Woo, S. H., Ranade, S., Weyer, A. D. et al.: Piezo2 is required for Merkel-cell mechanotransduction. Nature, 509, 622-626 (2014)[PubMed]
- Ikeda, R., Cha, M., Ling, J. et al.: Merkel cells transduce and encode tactile stimuli to drive abeta-afferent impulses. Cell, 157, 664-675 (2014)[PubMed] [新着論文レビュー]
- Ogawa, H.: The Merkel cell as a possible mechanoreceptor cell. Prog. Neurobiol., 49, 317-334 (1996)[PubMed]
- Lumpkin, E. A. & Hudspeth, A. J.: Detection of Ca2+ entry through mechanosensitive channels localizes the site of mechanoelectrical transduction in hair cells. Proc. Natl. Acad. Sci. USA, 92, 10297-10301 (1995)[PubMed]
著者プロフィール
略歴:2008年 東京大学大学院情報理工学系研究科 修了,同年 資生堂リサーチセンター 研究員を経て,2012年より慶應義塾大学大学院システムデザイン・マネジメント研究科 訪問研究員.
研究テーマ:Merkel細胞のメカノバイオロジー,触覚工学(触覚ディスプレイセンサーの開発),触覚心理学.
関心事:生体が機械刺激を受容して神経に伝達し中枢において知覚される過程の包括的な理解と,その計算論および工学的な応用.
馬場 欣哉(Yoshichika Baba)
略歴:1991年 岡山大学大学院自然科学研究科 修了,同年 北海道大学電子科学研究所 助手,2000年 富山大学工学部 講師,2002年 米国Illinois大学Chicago校Research Assistant Professor,2010年 米国Montana大学Missoula校Research Assistant Professorを経て,2011年より米国Columbia大学Associate Researcher.
研究テーマ:逃避行動の神経行動学.皮膚の接触感覚の信号処理.
関心事:外界を生体内の信号(スパイク列)に変換するときの符号化の方法.また,その信号から状況に適した情報を取り出してくる神経機構のアルゴリズム.
Srdjan Maksimovic
米国Columbia大学Postdoctoral Research Fellow.
Ellen A. Lumpkin
米国Columbia大学Associate Professor.
© 2014 仲谷正史・馬場欣哉・Srdjan Maksimovic・Ellen A. Lumpkin Licensed under CC 表示 2.1 日本
