僧帽細胞の誕生の時期は嗅覚における神経回路の構築に関与している
今村 文昭
(米国Yale大学School of Medicine,Department of Neurosurgery)
email:今村文昭
DOI: 10.7875/first.author.2011.044
Timing of neurogenesis is a determinant of olfactory circuitry.
Fumiaki Imamura, Albert E. Ayoub, Pasko Rakic, Charles A. Greer
Nature Neuroscience, 14, 331-337 (2011)
匂いの情報はまず嗅細胞を介して嗅上皮から嗅球へ伝わり,さらに僧帽細胞や房飾細胞により嗅皮質へと送られる.それぞれの匂い分子は嗅球において決められた領域の僧帽細胞および房飾細胞を活性化するが,嗅球におけるそれぞれの領域と嗅皮質とのつながりについてはわかっていないことが多い.この研究では,まず,マウスの僧帽細胞を誕生の時期の違いにより区別し,初期に誕生した僧帽細胞は主として嗅球の背内側領域に,逆に,後期に誕生したものは主として腹外側領域に分布していることを明らかにした.また,嗅皮質のひとつである嗅結節は後期に誕生した僧帽細胞から,より密な軸索投射をうけていることを見い出した.これらの結果は,嗅球の特定の領域と嗅皮質の特定の領域とがおもに同じ時期に誕生した僧帽細胞により結ばれていることを示唆しており,嗅球から嗅皮質への匂い情報の伝達様式についてその全体像の解明につながるものである.
鼻腔に取り込まれた匂い分子は嗅細胞の表面に発現する匂い分子受容体を活性化させ,その情報は嗅細胞の軸索を介してまず嗅球へと送られる.これまでの研究から,ヒトでは約350種類,マウスでは約1000種類の匂い分子受容体があって,個々の嗅細胞はそのうち1種類のみを発現していることがわかっている.嗅球の表面には多数の“糸球”とよばれる細胞体に囲まれた球状構造が存在し,個々の糸球には同種の匂い分子受容体を発現する嗅細胞の軸索が収束する(図1).その結果,嗅球の表面には糸球により匂い分子受容体に対応した“匂い地図”が形成されている1).糸球にはまた嗅球の投射ニューロンである僧帽細胞および房飾細胞も樹状突起を伸ばしてきており,糸球において嗅細胞の軸索とシナプスを形成している.したがって,個々の匂い分子は匂い地図における特定の糸球およびそれらにつながる僧帽細胞や房飾細胞を活性化する2).嗅球へ送られた匂い情報は僧帽細胞あるいは房飾細胞の軸索を介してさらに梨状皮質や嗅結節などからなる嗅皮質へと伝達される(図1).しかし,梨状皮質では個々の匂い分子に応答する細胞は分散して存在することが示されるなど3),嗅球の“匂い地図”に対応する地図は嗅皮質ではみつかっていない.そのため,嗅球における領域情報を僧帽細胞や房飾細胞がどのように嗅皮質へと伝達しているかを解明することは脳の匂い情報の処理機構を理解するうえで非常に重要である.
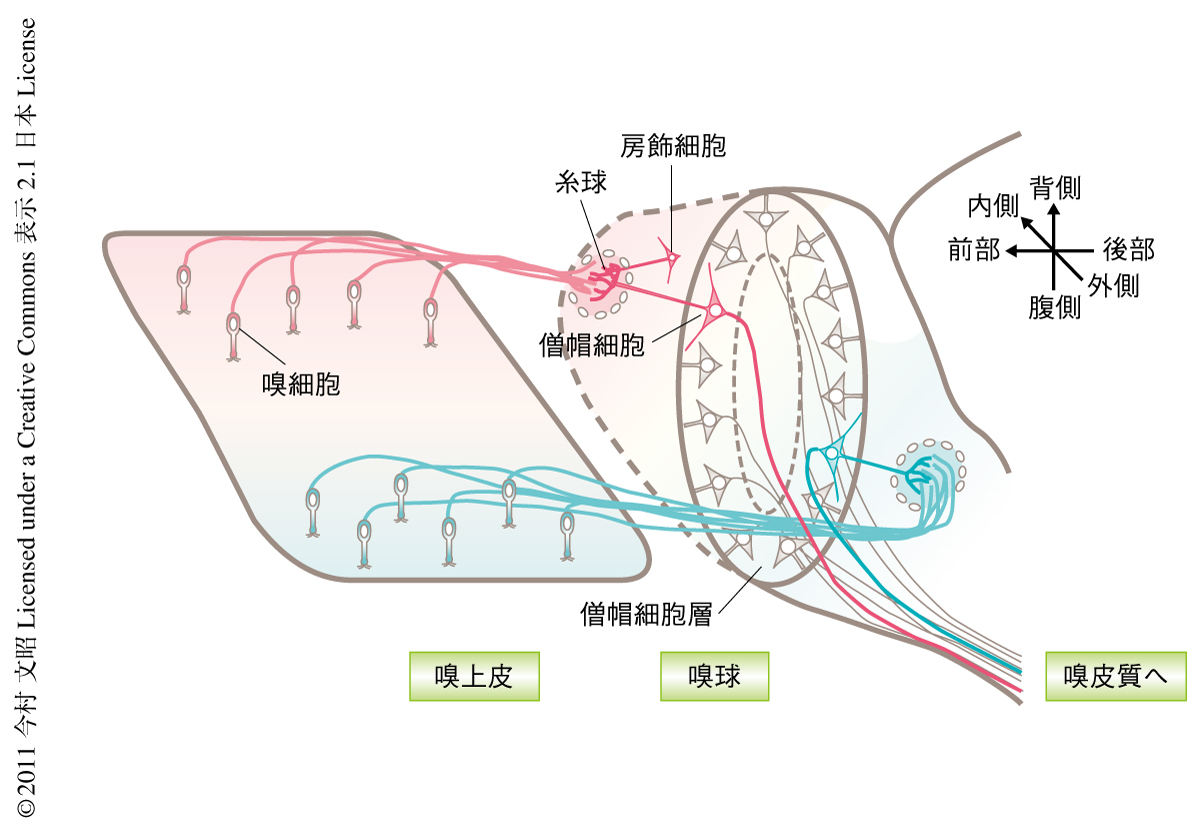
異なる匂い分子受容体を発現する嗅細胞が異なる糸球に軸索を収束させる分子機構は徐々に明らかになってきており,糸球は嗅細胞の軸索が発現する分子パターンの違いによりいくつかの領域に分けることができる4).しかし,僧帽細胞や房飾細胞ではこれらの領域に対応した分子の発現の違いはみつかっていない.6つの層からなる大脳皮質では,同じ時期に誕生した錐体細胞は同じ層に移動し層ごとに異なる領域へと軸索を投射することがよく知られている5).そこでこの研究では,僧帽細胞の誕生の時期に着目し,異なる時期に誕生した僧帽細胞は嗅球における細胞体の分布および嗅結節への軸索投射のパターンの異なることを示した.これは,嗅球の僧帽細胞でも誕生の時期が細胞体の場所や軸索の投射先を制御する一因であることを示唆した結果である.
まず,僧帽細胞をその誕生の時期により区別し嗅球での分布を比較した.実験は,BrdU,IdU,CldUなどのチミジン類似体を妊娠マウスの腹腔に投与し,生まれてきた仔(生後20日)の嗅球においてこのチミジン類似体を取り込んだ僧帽細胞を調べることで行った.チミジン類似体はチミジンと同様にDNA合成期に染色体に取り込まれるため,チミジン類似体に陽性を示す僧帽細胞はその投与時期に誕生したものと考えられる.実験の結果,ほとんどの僧帽細胞は胎生9日期から胎生13日期のあいだに誕生することがわかった.興味深いことに,初期(胎生9日期および胎生10日期)に誕生した僧帽細胞は嗅球においても背内側領域に多くみられ,逆に,後期(胎生12日期および胎生13日期)に誕生したものの多くは腹外側領域にみつかった.胎生11日期に誕生した僧帽細胞では嗅球における分布に差はみられなかった.この結果より,誕生の時期の異なる僧帽細胞は嗅球において背内側領域と腹外側領域とに異なって分布していることが示された.さらに,生後0日の嗅球においても同様に,初期に誕生した僧帽細胞は背内側領域に多く,後期に誕生したものは腹外側領域に多いことが確認され,僧帽細胞の基本的な分布は胚期に確立されることが示された.
誕生の時期の異なる僧帽細胞が嗅球において異なった分布をする機構を知るため,つぎに,胎生10日期,胎生11日期,胎生12日期に誕生した僧帽細胞の分布を胎生15日期の嗅球で調べた.嗅球の発生において,僧帽細胞は脳室帯で誕生し嗅球の表面へと放射状に移動して最終的に僧帽細胞層へと到達することが知られている.実験の結果,胎生15日期の嗅球では,胎生10日期に誕生した僧帽細胞はすでに僧帽細胞層へ到達していたが,胎生12日期に誕生したものの大半は脳室帯もしくは脳室帯と僧帽細胞層とのあいだに位置する中間層に存在していることがわかった.さらに,それらの嗅球における分布を調べた結果,胎生12日期に誕生した僧帽細胞は腹外側領域にかたよって分布していたのに対して,胎生10日期に誕生したものは一様に分布していた.また,胎生11日期に誕生した僧帽細胞の分布は腹外側領域にややかたよっていた.これらの結果は,後期に誕生する僧帽細胞は選択的に腹外側領域に組み込まれることを示していた.これによって,結果的に胎生10日期に誕生した僧帽細胞においても背内側領域と腹外側領域とのあいだで密度差の生じることとなり,誕生の時期に依存的に僧帽細胞の嗅球における分布の異なる理由が説明された.
それでは,後期に誕生した僧帽細胞はどのようにして選択的に嗅球の腹外側領域に組み込まれるのだろうか? 胎生12日期に誕生した僧帽細胞の分布を胎生13日期および胎生14日期の嗅球で調べた.胎生13日期では胎生12日期に誕生した細胞の大半は脳室帯にみられた.その細胞核は嗅球の内部を放射状に伸びる放射状グリア細胞にそって長く伸展しており,脳室帯を表面にむかって移動していることが示唆された.しかし,嗅球における分布を調べると腹外側領域にかたよった分布はみられず,分布のピークは背外側領域であった.胎生14日期になると胎生12日期に誕生した細胞の多くは中間層に移動しており,これらの細胞核は水平方向に長く伸展していた.嗅球における分布のピークは背外側領域から外側領域へと変化していたものの,胎生15日期でみられたような腹外側領域へのかたよった分布は依然としてみられなかった.しかし,嗅球における分布をその前部と後部とで比較したところ,後部の嗅球においてより多くの細胞が腹外側領域にあることがわかった.これらの結果から筆者らは,“後期に誕生した僧帽細胞は脳室帯を放射状に中間層まで移動したのちに,その向きを変え,中間層をさらに嗅球の後部の腹外側領域へと水平に移動する”という仮説をたてた.
この仮説を検証するため,つぎに嗅球切片を用いたin vitroタイムラプスイメージング実験を行い,発生の段階で嗅球の内部を水平に移動する細胞が存在するかどうかを調べた.実験にはDoublecortinの遺伝子プロモーターのもとGFPを発現する遺伝子改変マウスを用い,GFPを発現する細胞の挙動を顕微鏡下で経時的に観察した.微小管結合タンパク質のひとつDoublecortinは移動中のニューロンに発現することが知られているため,この遺伝子改変マウスはニューロンの移動のようすを調べるのによく用いられている.実験の結果,胎生14日期および胎生15日期の嗅球の内部を後部外側にむかって水平方向に移動するニューロンが実際に存在することが確認された.さらに,タイムラプスイメージングの終了ののち用いた切片を免疫染色することで,水平に移動した細胞が僧帽細胞のマーカーであるTbr1を発現していることを確認した.また,脳室帯を放射方向に移動する細胞,移動の方向を放射方向から水平方向へと変える細胞の存在も確認できた.この結果は,筆者らの仮説を支持していた.
大脳皮質の錐体細胞と同様に,僧帽細胞が脳室帯を放射状に移動する際には嗅球の放射状グリア細胞が足場を提供しているものと考えられる.それでは,後期に誕生した僧帽細胞が中間層を水平移動する際の足場は何であろうか? この候補のひとつが初期に誕生した僧帽細胞の軸索である.まず,胎生14日期の嗅球を用いて,すでに形成されている僧帽細胞の軸索が嗅球では中間層を後部腹外側領域へと走行していることをみつけた.また,さきに述べたように胎生12日期に誕生した僧帽細胞はこの時期には中間層で水平方向に伸びた形態を示しているが,これらの細胞が中間層で僧帽細胞の軸索と近接していることも発見した.さらに,電子顕微鏡による解析を行い,中間層で僧帽細胞と軸索とがじかに接していることを明らかにした.これらの観察の結果は,後期に誕生した僧帽細胞が中間層を水平に移動する際に,すでに形成されている僧帽細胞の軸索を足場としている可能性を示していた.これは,初期に誕生した僧帽細胞が最初は嗅球において一様に分布することの理由(水平移動のための足場が形成されていないため)を説明することもできる有力な説と考えており,今後,さらなる検証を行う.
過去の研究により,嗅皮質のひとつである嗅結節は腹外側領域に存在する僧帽細胞からより密な軸索投射をうけていることが示されている6).後期に誕生する僧帽細胞もまた腹外側領域に多くみられることから,僧帽細胞の誕生の時期と嗅結節への軸索投射との関連性を調べた.実験では,胎生10日期,胎生11日期,または,胎生12日期のマウスの腹腔内にチミジン類似体EdUを投与した.そののち,生まれてきた仔(生後5日)の嗅結節に蛍光トレーサーDiIを注入し,嗅結節に軸索投射している僧帽細胞を逆行性に標識した.これにより,胎生10日期,胎生11日期,胎生12日期のそれぞれの時期に誕生しかつ嗅結節へと軸索を投射している僧帽細胞がEdUとDiIの両方で標識される.DiI陽性の僧帽細胞のうちEdUでも標識されているものの割合を解析したところ,EdUを胎生12日期で投与したときがもっとも高く,胎生10日期で投与したときがもっとも低かった.この結果は,嗅結節は後期に誕生した僧帽細胞からより多くの軸索投射をうけていることを示していた.
これまでの研究から,嗅細胞の軸索の投射先である嗅球の糸球が分子発現パターンの違いにより背内側領域と腹外側領域とに区別できることはよく知られている7,8).また,背内側領域の糸球へ軸索投射する嗅細胞を選択的に欠失させた遺伝子改変マウスは,嗅覚は保持しているもののネコやキツネの匂いに対し忌避行動をとらないことが示され,それぞれの領域は特定の生理学的な機能を担っていることが示唆されていた9).この研究では,初期と後期に誕生した僧帽細胞がそれぞれ嗅球において背内側領域と腹外側領域に異なって分布していることを明らかにした(図2).これは,嗅球の背内側領域と腹外側領域での匂い情報の処理にはそれぞれ,おもに初期と後期に誕生した僧帽細胞が関与していることを示しており,それぞれの領域の機能差を生み出す機構の一端を明らかにした研究結果である.
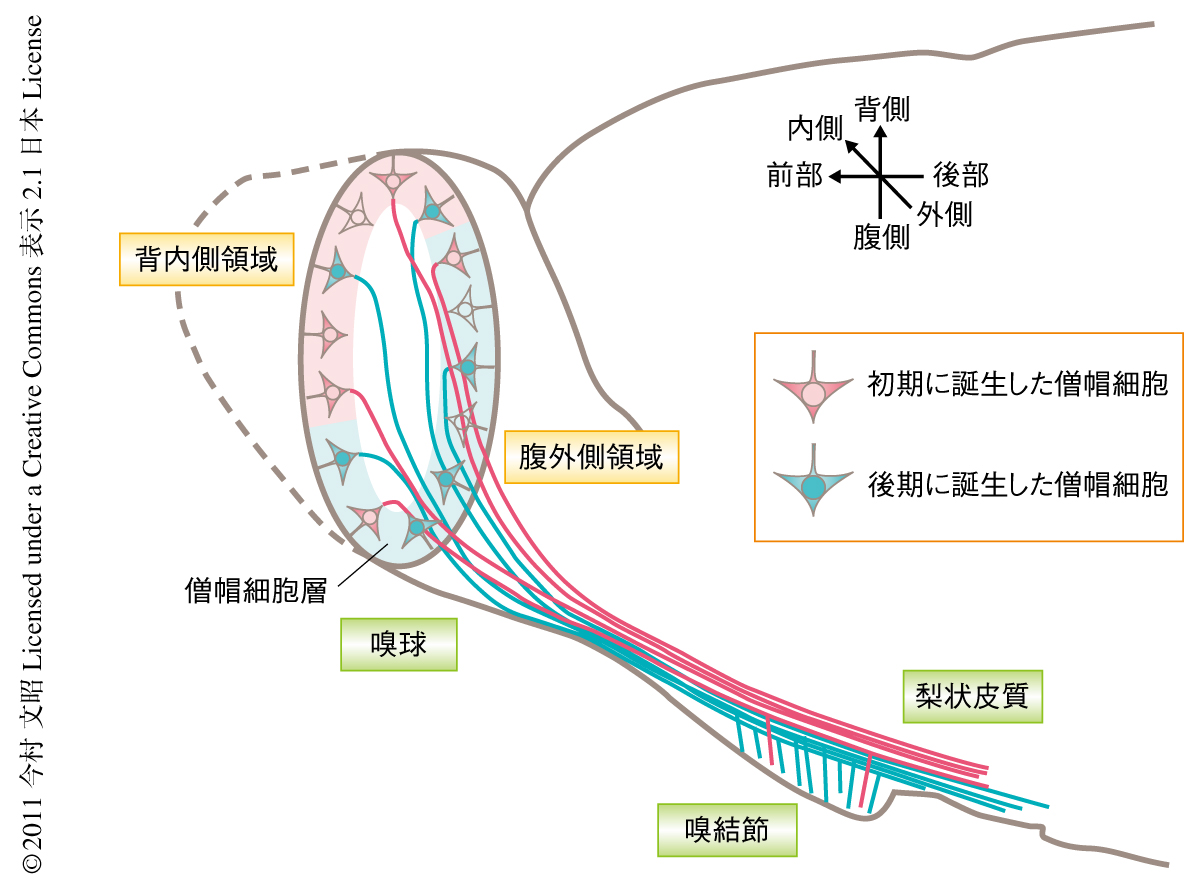
この研究ではさらに,嗅皮質のひとつである嗅結節は後期に誕生した僧帽細胞からより多くの軸索投射をうけていることを示した(図2).さらに最近の研究では,同じく嗅球からの入力を受け取る扁桃体には背内側領域に存在する僧帽細胞が密に軸索を投射していることが明らかにされている10)(新着論文レビュー でも掲載).筆者らが得た結果とあわせて,扁桃体には主として初期に誕生した僧帽細胞が軸索投射しているものと考えている.さらに,そのほかの嗅皮質への僧帽細胞の軸索投射のパターンもある程度まで誕生の時期により制御されているものと考えており,この研究を足がかりに,嗅球から嗅皮質への匂い情報の伝達様式および嗅皮質のそれぞれの領域が担う機能の解明がいっそう進むことを期待している.
略歴:2003年 京都大学大学院理学研究科 修了,同年 東京大学大学院医学系研究科 博士研究員を経て,2005年 米国Yale大学 博士研究員,2010年より同Associate Research Scientist.
研究テーマ:嗅覚の神経回路の形成機構.
抱負:われわれの脳で匂い情報がどのように処理されているのかを明らかにしたい.
© 2011 今村 文昭 Licensed under CC 表示 2.1 日本
(米国Yale大学School of Medicine,Department of Neurosurgery)
email:今村文昭
DOI: 10.7875/first.author.2011.044
Timing of neurogenesis is a determinant of olfactory circuitry.
Fumiaki Imamura, Albert E. Ayoub, Pasko Rakic, Charles A. Greer
Nature Neuroscience, 14, 331-337 (2011)
要 約
匂いの情報はまず嗅細胞を介して嗅上皮から嗅球へ伝わり,さらに僧帽細胞や房飾細胞により嗅皮質へと送られる.それぞれの匂い分子は嗅球において決められた領域の僧帽細胞および房飾細胞を活性化するが,嗅球におけるそれぞれの領域と嗅皮質とのつながりについてはわかっていないことが多い.この研究では,まず,マウスの僧帽細胞を誕生の時期の違いにより区別し,初期に誕生した僧帽細胞は主として嗅球の背内側領域に,逆に,後期に誕生したものは主として腹外側領域に分布していることを明らかにした.また,嗅皮質のひとつである嗅結節は後期に誕生した僧帽細胞から,より密な軸索投射をうけていることを見い出した.これらの結果は,嗅球の特定の領域と嗅皮質の特定の領域とがおもに同じ時期に誕生した僧帽細胞により結ばれていることを示唆しており,嗅球から嗅皮質への匂い情報の伝達様式についてその全体像の解明につながるものである.
はじめに
鼻腔に取り込まれた匂い分子は嗅細胞の表面に発現する匂い分子受容体を活性化させ,その情報は嗅細胞の軸索を介してまず嗅球へと送られる.これまでの研究から,ヒトでは約350種類,マウスでは約1000種類の匂い分子受容体があって,個々の嗅細胞はそのうち1種類のみを発現していることがわかっている.嗅球の表面には多数の“糸球”とよばれる細胞体に囲まれた球状構造が存在し,個々の糸球には同種の匂い分子受容体を発現する嗅細胞の軸索が収束する(図1).その結果,嗅球の表面には糸球により匂い分子受容体に対応した“匂い地図”が形成されている1).糸球にはまた嗅球の投射ニューロンである僧帽細胞および房飾細胞も樹状突起を伸ばしてきており,糸球において嗅細胞の軸索とシナプスを形成している.したがって,個々の匂い分子は匂い地図における特定の糸球およびそれらにつながる僧帽細胞や房飾細胞を活性化する2).嗅球へ送られた匂い情報は僧帽細胞あるいは房飾細胞の軸索を介してさらに梨状皮質や嗅結節などからなる嗅皮質へと伝達される(図1).しかし,梨状皮質では個々の匂い分子に応答する細胞は分散して存在することが示されるなど3),嗅球の“匂い地図”に対応する地図は嗅皮質ではみつかっていない.そのため,嗅球における領域情報を僧帽細胞や房飾細胞がどのように嗅皮質へと伝達しているかを解明することは脳の匂い情報の処理機構を理解するうえで非常に重要である.
異なる匂い分子受容体を発現する嗅細胞が異なる糸球に軸索を収束させる分子機構は徐々に明らかになってきており,糸球は嗅細胞の軸索が発現する分子パターンの違いによりいくつかの領域に分けることができる4).しかし,僧帽細胞や房飾細胞ではこれらの領域に対応した分子の発現の違いはみつかっていない.6つの層からなる大脳皮質では,同じ時期に誕生した錐体細胞は同じ層に移動し層ごとに異なる領域へと軸索を投射することがよく知られている5).そこでこの研究では,僧帽細胞の誕生の時期に着目し,異なる時期に誕生した僧帽細胞は嗅球における細胞体の分布および嗅結節への軸索投射のパターンの異なることを示した.これは,嗅球の僧帽細胞でも誕生の時期が細胞体の場所や軸索の投射先を制御する一因であることを示唆した結果である.
1.誕生の時期の異なる僧帽細胞は嗅球において異なる分布を示す
まず,僧帽細胞をその誕生の時期により区別し嗅球での分布を比較した.実験は,BrdU,IdU,CldUなどのチミジン類似体を妊娠マウスの腹腔に投与し,生まれてきた仔(生後20日)の嗅球においてこのチミジン類似体を取り込んだ僧帽細胞を調べることで行った.チミジン類似体はチミジンと同様にDNA合成期に染色体に取り込まれるため,チミジン類似体に陽性を示す僧帽細胞はその投与時期に誕生したものと考えられる.実験の結果,ほとんどの僧帽細胞は胎生9日期から胎生13日期のあいだに誕生することがわかった.興味深いことに,初期(胎生9日期および胎生10日期)に誕生した僧帽細胞は嗅球においても背内側領域に多くみられ,逆に,後期(胎生12日期および胎生13日期)に誕生したものの多くは腹外側領域にみつかった.胎生11日期に誕生した僧帽細胞では嗅球における分布に差はみられなかった.この結果より,誕生の時期の異なる僧帽細胞は嗅球において背内側領域と腹外側領域とに異なって分布していることが示された.さらに,生後0日の嗅球においても同様に,初期に誕生した僧帽細胞は背内側領域に多く,後期に誕生したものは腹外側領域に多いことが確認され,僧帽細胞の基本的な分布は胚期に確立されることが示された.
2.後期に誕生した僧帽細胞は選択的に嗅球の腹外側領域に組み込まれる
誕生の時期の異なる僧帽細胞が嗅球において異なった分布をする機構を知るため,つぎに,胎生10日期,胎生11日期,胎生12日期に誕生した僧帽細胞の分布を胎生15日期の嗅球で調べた.嗅球の発生において,僧帽細胞は脳室帯で誕生し嗅球の表面へと放射状に移動して最終的に僧帽細胞層へと到達することが知られている.実験の結果,胎生15日期の嗅球では,胎生10日期に誕生した僧帽細胞はすでに僧帽細胞層へ到達していたが,胎生12日期に誕生したものの大半は脳室帯もしくは脳室帯と僧帽細胞層とのあいだに位置する中間層に存在していることがわかった.さらに,それらの嗅球における分布を調べた結果,胎生12日期に誕生した僧帽細胞は腹外側領域にかたよって分布していたのに対して,胎生10日期に誕生したものは一様に分布していた.また,胎生11日期に誕生した僧帽細胞の分布は腹外側領域にややかたよっていた.これらの結果は,後期に誕生する僧帽細胞は選択的に腹外側領域に組み込まれることを示していた.これによって,結果的に胎生10日期に誕生した僧帽細胞においても背内側領域と腹外側領域とのあいだで密度差の生じることとなり,誕生の時期に依存的に僧帽細胞の嗅球における分布の異なる理由が説明された.
3.後期に誕生した僧帽細胞は嗅球の内部を水平方向に移動する
それでは,後期に誕生した僧帽細胞はどのようにして選択的に嗅球の腹外側領域に組み込まれるのだろうか? 胎生12日期に誕生した僧帽細胞の分布を胎生13日期および胎生14日期の嗅球で調べた.胎生13日期では胎生12日期に誕生した細胞の大半は脳室帯にみられた.その細胞核は嗅球の内部を放射状に伸びる放射状グリア細胞にそって長く伸展しており,脳室帯を表面にむかって移動していることが示唆された.しかし,嗅球における分布を調べると腹外側領域にかたよった分布はみられず,分布のピークは背外側領域であった.胎生14日期になると胎生12日期に誕生した細胞の多くは中間層に移動しており,これらの細胞核は水平方向に長く伸展していた.嗅球における分布のピークは背外側領域から外側領域へと変化していたものの,胎生15日期でみられたような腹外側領域へのかたよった分布は依然としてみられなかった.しかし,嗅球における分布をその前部と後部とで比較したところ,後部の嗅球においてより多くの細胞が腹外側領域にあることがわかった.これらの結果から筆者らは,“後期に誕生した僧帽細胞は脳室帯を放射状に中間層まで移動したのちに,その向きを変え,中間層をさらに嗅球の後部の腹外側領域へと水平に移動する”という仮説をたてた.
この仮説を検証するため,つぎに嗅球切片を用いたin vitroタイムラプスイメージング実験を行い,発生の段階で嗅球の内部を水平に移動する細胞が存在するかどうかを調べた.実験にはDoublecortinの遺伝子プロモーターのもとGFPを発現する遺伝子改変マウスを用い,GFPを発現する細胞の挙動を顕微鏡下で経時的に観察した.微小管結合タンパク質のひとつDoublecortinは移動中のニューロンに発現することが知られているため,この遺伝子改変マウスはニューロンの移動のようすを調べるのによく用いられている.実験の結果,胎生14日期および胎生15日期の嗅球の内部を後部外側にむかって水平方向に移動するニューロンが実際に存在することが確認された.さらに,タイムラプスイメージングの終了ののち用いた切片を免疫染色することで,水平に移動した細胞が僧帽細胞のマーカーであるTbr1を発現していることを確認した.また,脳室帯を放射方向に移動する細胞,移動の方向を放射方向から水平方向へと変える細胞の存在も確認できた.この結果は,筆者らの仮説を支持していた.
4.僧帽細胞の水平移動において僧帽細胞の軸索が足場を提供する
大脳皮質の錐体細胞と同様に,僧帽細胞が脳室帯を放射状に移動する際には嗅球の放射状グリア細胞が足場を提供しているものと考えられる.それでは,後期に誕生した僧帽細胞が中間層を水平移動する際の足場は何であろうか? この候補のひとつが初期に誕生した僧帽細胞の軸索である.まず,胎生14日期の嗅球を用いて,すでに形成されている僧帽細胞の軸索が嗅球では中間層を後部腹外側領域へと走行していることをみつけた.また,さきに述べたように胎生12日期に誕生した僧帽細胞はこの時期には中間層で水平方向に伸びた形態を示しているが,これらの細胞が中間層で僧帽細胞の軸索と近接していることも発見した.さらに,電子顕微鏡による解析を行い,中間層で僧帽細胞と軸索とがじかに接していることを明らかにした.これらの観察の結果は,後期に誕生した僧帽細胞が中間層を水平に移動する際に,すでに形成されている僧帽細胞の軸索を足場としている可能性を示していた.これは,初期に誕生した僧帽細胞が最初は嗅球において一様に分布することの理由(水平移動のための足場が形成されていないため)を説明することもできる有力な説と考えており,今後,さらなる検証を行う.
5.嗅結節は後期に誕生した僧帽細胞からより多くの軸索投射をうける
過去の研究により,嗅皮質のひとつである嗅結節は腹外側領域に存在する僧帽細胞からより密な軸索投射をうけていることが示されている6).後期に誕生する僧帽細胞もまた腹外側領域に多くみられることから,僧帽細胞の誕生の時期と嗅結節への軸索投射との関連性を調べた.実験では,胎生10日期,胎生11日期,または,胎生12日期のマウスの腹腔内にチミジン類似体EdUを投与した.そののち,生まれてきた仔(生後5日)の嗅結節に蛍光トレーサーDiIを注入し,嗅結節に軸索投射している僧帽細胞を逆行性に標識した.これにより,胎生10日期,胎生11日期,胎生12日期のそれぞれの時期に誕生しかつ嗅結節へと軸索を投射している僧帽細胞がEdUとDiIの両方で標識される.DiI陽性の僧帽細胞のうちEdUでも標識されているものの割合を解析したところ,EdUを胎生12日期で投与したときがもっとも高く,胎生10日期で投与したときがもっとも低かった.この結果は,嗅結節は後期に誕生した僧帽細胞からより多くの軸索投射をうけていることを示していた.
おわりに
これまでの研究から,嗅細胞の軸索の投射先である嗅球の糸球が分子発現パターンの違いにより背内側領域と腹外側領域とに区別できることはよく知られている7,8).また,背内側領域の糸球へ軸索投射する嗅細胞を選択的に欠失させた遺伝子改変マウスは,嗅覚は保持しているもののネコやキツネの匂いに対し忌避行動をとらないことが示され,それぞれの領域は特定の生理学的な機能を担っていることが示唆されていた9).この研究では,初期と後期に誕生した僧帽細胞がそれぞれ嗅球において背内側領域と腹外側領域に異なって分布していることを明らかにした(図2).これは,嗅球の背内側領域と腹外側領域での匂い情報の処理にはそれぞれ,おもに初期と後期に誕生した僧帽細胞が関与していることを示しており,それぞれの領域の機能差を生み出す機構の一端を明らかにした研究結果である.
この研究ではさらに,嗅皮質のひとつである嗅結節は後期に誕生した僧帽細胞からより多くの軸索投射をうけていることを示した(図2).さらに最近の研究では,同じく嗅球からの入力を受け取る扁桃体には背内側領域に存在する僧帽細胞が密に軸索を投射していることが明らかにされている10)(新着論文レビュー でも掲載).筆者らが得た結果とあわせて,扁桃体には主として初期に誕生した僧帽細胞が軸索投射しているものと考えている.さらに,そのほかの嗅皮質への僧帽細胞の軸索投射のパターンもある程度まで誕生の時期により制御されているものと考えており,この研究を足がかりに,嗅球から嗅皮質への匂い情報の伝達様式および嗅皮質のそれぞれの領域が担う機能の解明がいっそう進むことを期待している.
文 献
- Mori, K., Nagao, H. & Yoshihara, Y.: The olfactory bulb: Coding and processing of odor molecule information. Science, 286, 711-715 (1999)[PubMed]
- Mori, K., Takahashi, Y. K., Igarashi, K. M. et al.: Maps of odorant molecular features in the mammalian olfactory bulb. Physiol. Rev., 86, 409-433 (2006)[PubMed]
- Stettler, D. D. & Axel, R.: Representations of odor in the piriform cortex. Neuron, 63, 854-864 (2009)[PubMed]
- Sakano, H.: Neural map formation in the mouse olfactory system. Neuron, 67, 530-542 (2010)[PubMed]
- Molyneaux, B. J., Arlotta, P., Menezes, J. R. et al.: Neuronal subtype specification in the cerebral cortex. Nat. Rev. Neurosci., 8, 427-437 (2007)[PubMed]
- Scott, J. W., McBride, R. L. & Schneider, S. P.: The organization of projections from the olfactory bulb to the piriform cortex and olfactory tubercle in the rat. J. Comp. Neurol., 194, 519-534 (1980)[PubMed]
- Yoshihara, Y., Kawasaki, M., Tamada, A. et al.: OCAM: A new member of the neural cell adhesion molecule family related to zone-to-zone projection of olfactory and vomeronasal axons. J. Neurosci., 17, 5830-5842 (1997)[PubMed]
- Oka, Y., Kobayakawa, K., Nishizumi, H. et al.: O-MACS, a novel member of the medium-chain acyl-coa synthetase family, specifically expressed in the olfactory epithelium in a zone-specific manner. Eur. J. Biochem., 270, 1995-2004 (2003)[PubMed]
- Kobayakawa, K., Kobayakawa, R., Matsumoto, H. et al.: Innate versus learned odour processing in the mouse olfactory bulb. Nature, 450, 503-508 (2007)[PubMed]
- Miyamichi, K., Amat, F., Moussavi, F. et al.: Cortical representations of olfactory input by trans-synaptic tracing. Nature, 472, 191-196 (2011)[PubMed] [新着論文レビュー]
著者プロフィール
略歴:2003年 京都大学大学院理学研究科 修了,同年 東京大学大学院医学系研究科 博士研究員を経て,2005年 米国Yale大学 博士研究員,2010年より同Associate Research Scientist.
研究テーマ:嗅覚の神経回路の形成機構.
抱負:われわれの脳で匂い情報がどのように処理されているのかを明らかにしたい.
© 2011 今村 文昭 Licensed under CC 表示 2.1 日本
