模倣学習を発動させる中脳から感覚運動野へのドーパミンの出力
田中雅史・Richard Mooney
(米国Duke大学Department of Neurobiology)
email:田中雅史
DOI: 10.7875/first.author.2018.106
A mesocortical dopamine circuit enables the cultural transmission of vocal behaviour.
Masashi Tanaka, Fangmiao Sun, Yulong Li, Richard Mooney
Nature, 563, 117-120 (2018)
模倣は,規範的なモデルとなる他者の行動を記憶し自らの行動により正確に再現することを可能にする,きわめて効率的な学習能力である.しかし,脳がどのように適切な対象を選び模倣学習を発動させるのか,その機構については不明であった.この研究において,筆者らは,歌の正確な伝承能力をもつキンカチョウという鳥に着目し,成鳥の歌を聞いて模倣を開始するときの幼いキンカチョウの脳について調べた.その結果,中脳の水道周囲灰白質に存在するニューロンが,成鳥から歌いかけられたときにのみ強い神経活動を示し,歌のさえずりを制御する皮質の感覚運動野HVCへとドーパミンを出力することが明らかにされた.幼鳥が歌を聞いているときにHVCにおけるドーパミンのはたらきを阻害するとその歌の模倣は発動されず,逆に,HVCへのドーパミンの出力をひき起こすと通常は模倣されないはずのスピーカーから流れた歌によっても模倣学習が発動された.ドーパミンは幼鳥のHVCの神経活動に可塑的な変化をひき起こすとともに,幼鳥の歌を成熟化させる作用をもつことも明らかにされ,歌の模倣学習の発動にはこの運動をつかさどる皮質へと出力されるドーパミンが重要な役割をはたすことが示された.
模倣は,ヒトなど霊長目や一部の鳥のほかにはごくかぎられた動物のみがもつ,きわめてめずらしい学習能力である1).模倣の能力をもつ動物は世代をこえてすぐれた行動を伝承することにより,それぞれに固有の文化や技術を発展させることができる.しかし,外界のあらゆる対象を模倣していては安定した技術や文化の継承はできないであろう.ヒトは適切な対象を選択しその行動のみを模倣することができるが,脳がどのようにして模倣を発動させるのか,その詳細な機構は謎につつまれている.
歌をさえずる鳥であるスズメ亜目の一種のキンカチョウは,親の歌を正確に模倣し,その歌を子孫へ伝え世代をこえた文化として歌を伝承する能力をもつ2).キンカチョウは幼少期に出会った成熟した鳥の歌を記憶し,数十日かけてくり返し練習することにより模倣を完了する.歌の模倣においては社会的な相互作用が重要であることが知られ,通常,成鳥の歌をスピーカーから流しただけでは模倣は発動されない3).この研究において,筆者らは,キンカチョウの歌の模倣を制御する脳の機能について調べるため,成熟した歌のさえずりに必要不可欠であることが知られる皮質の感覚運動野HVCに着目した4,5).HVCは下流の運動野,大脳基底核,聴覚野へと運動のシグナルを送ることにより歌のさえずりを制御しているが,近年の研究により,歌の学習にも重要な役割をはたすことが明らかになりはじめている6,7).そこで,模倣学習を開始するころの幼いキンカチョウのHVCにトレーサーを注入したところ,HVCは中脳の水道周囲灰白質という領域から密なドーパミンの入力をうけることが明らかにされた.
水道周囲灰白質の機能について知るため,自由行動下の幼いキンカチョウの水道周囲灰白質にテトロード電極を埋め込み神経活動を記録した.その結果,水道周囲灰白質に存在するニューロンの多くは適切な模倣の対象であるオスの成鳥から歌いかけられたときにのみ強い神経活動を起こすことが明らかにされた.同じ歌をスピーカーから流しただけでは顕著な応答はみられず,また,歌わないオスやメスの成鳥に出会ったとき,あるいは,メスの成鳥がいる条件にてオスの歌をスピーカーから流したときには,強い応答は起こらなかった.
水道周囲灰白質に存在するニューロンの一部はドーパミン作動性ニューロンであり,神経活動によりHVCへとドーパミンを出力する.HVCへのドーパミンの出力を検出するため,ドーパミンのセンサーであるGRABDA 8) をHVCのニューロンに発現させ,2光子顕微鏡により観察した.すると,オスの成鳥から歌いかけられたときにのみ,HVCへと多量のドーパミンが出力されることがわかった.このドーパミンの出力は,水道周囲灰白質のドーパミン作動性ニューロンを薬剤を用いて破壊することにより消失したため,HVCへのドーパミンの出力の大部分は水道周囲灰白質に由来すると考えられた.これらの結果から,水道周囲灰白質は適切な模倣の対象であるオスの成鳥の歌にのみ選択的に応答し,HVCへとドーパミンを出力することが明らかにされた.
HVCへと出力されるドーパミンが模倣にはたす機能について調べるため,幼いキンカチョウが親の歌を聞いて記憶しはじめる時期に,薬剤を用いてHVCのドーパミン作動性ニューロンを破壊したところ,その歌を模倣できなくなることがわかった.一方,幼鳥が親の歌を十分に記憶したであろうころに薬剤を用いてHVCのドーパミン作動性ニューロンを破壊したところ,幼鳥は問題なく親の歌を模倣した.したがって,HVCへと出力されるドーパミンは,模倣学習においても最初期の,歌を記憶する時期に必要であることが示唆された.
幼いころに親から隔離された幼鳥が,はじめて出会った成鳥から歌いかけられたときに,ドーパミン受容体の阻害剤を用いてHVCにおけるドーパミンのシグナル伝達を遮断したところ,幼鳥は歌を模倣できなくなることがわかった.一方,成鳥と出会って歌を聞きおえたのちにHVCにおけるドーパミンのシグナル伝達を遮断しても,幼鳥は問題なく歌を模倣した.したがって,HVCへと出力されるドーパミンは,成鳥がさえずる歌を模倣のため記憶するときに重要な役割をはたすことが示された.
もし,ドーパミンが適切な模倣の対象である成鳥が歌っているという情報をHVCに伝達することによりその歌の記憶を助けているのであれば,人工的にHVCにドーパミンを出力させることにより,通常は模倣しないスピーカーから流した歌に対しても模倣学習を発動させることができるかもしれない.そこで,水道周囲灰白質のニューロンに光により神経活動を駆動できるチャネルロドプシン2を発現させ,HVCに光を照射することにより水道周囲灰白質からHVCへのドーパミンの出力を駆動したところ,スピーカーから流した歌に対しても模倣が発動された.これらの結果から,HVCへと出力されたドーパミンはそのとき聞こえた歌に対する模倣学習を発動させることが明らかにされた.
ドーパミンはHVCに出力されることによりどのように模倣学習を発動させるのだろうか? 幼鳥のHVCにおいて神経活動を記録したところ,成鳥から歌いかけられたあとには,HVCにおいてときおり自発的に生じるバースト発火がより頻繁に観察されるようになった.HVCにおけるバースト発火は,おもにnucleus interfacialis(NIf)とよばれる聴覚野6) からの入力により駆動されることが知られているため,バースト発火の増大からNIfからHVCへと伝達される聴覚のシグナルが増強されたことが示唆された.そこで,スピーカーから歌を流して幼鳥のHVCへと伝達される聴覚のシグナルについて調べたところ,その歌を成鳥から歌いかけられるまでは明瞭な応答がみられなかったのに対し,成鳥から歌いかけられたあとには,あたかもその歌を記憶したかのように,歌の特定の要素に対する応答が出現した.したがって,ドーパミンはHVCの神経活動に可塑的な変化をひき起こし,成鳥の歌の表象を記録することにより模倣を発動させると考えられた.
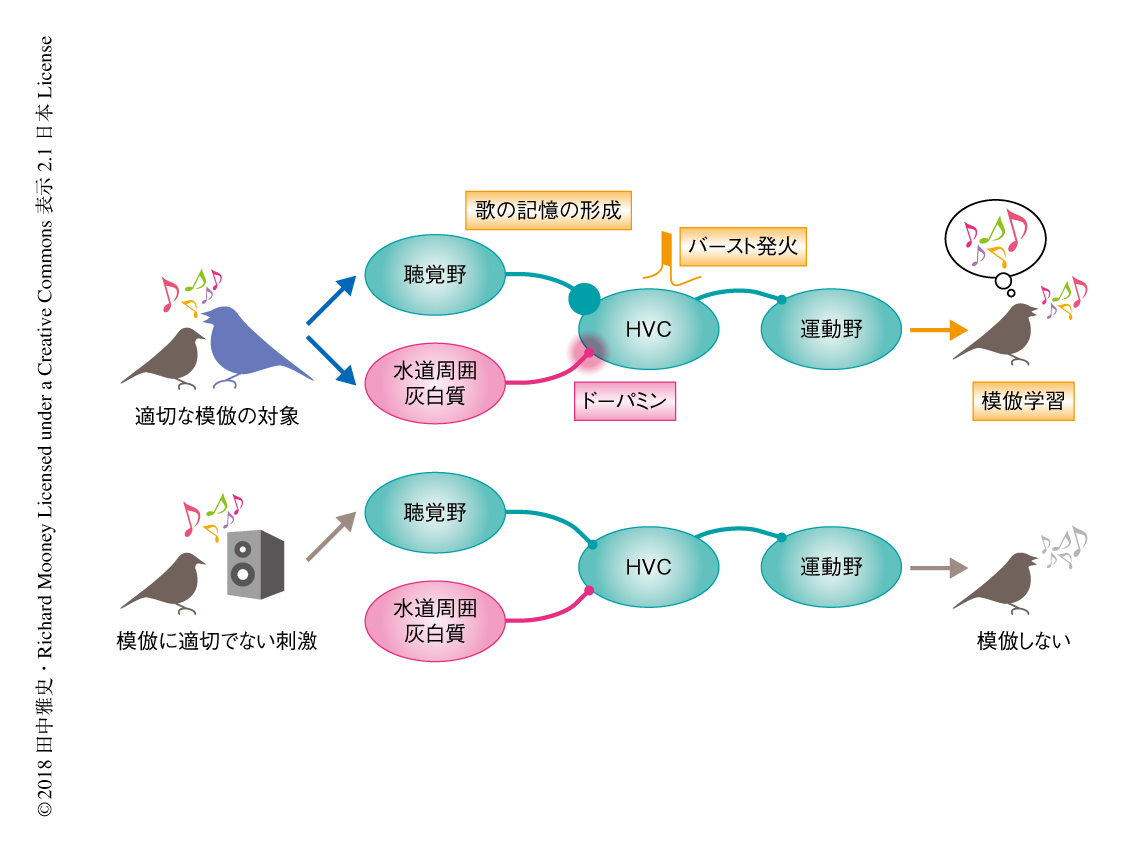
HVCにおける神経活動は運動野や大脳基底核に伝達されることにより歌のさえずりを制御する4).したがって,ドーパミンの出力によりひき起こされたHVCにおける神経活動の可塑的な変化は,ただちに幼鳥の歌を変化させ模倣を発動させる可能性がある.事実,成鳥から歌いかけられたあとの幼鳥の歌を解析したところ,その歌には時間的および周波数的に成鳥の歌に近い複雑な構造がみられた.HVCにおいてドーパミンのシグナル伝達を遮断した状態においては,成鳥から歌いかけられたあとの幼鳥の歌の変化もHVCにおける神経活動の可塑的な変化も起こらなかった.したがって,成鳥から歌いかけられると,幼鳥のHVCへと出力されたドーパミンの作用によりHVCの神経活動に可塑的な変化がひき起こされると同時に,早くも幼鳥のさえずりの制御がはじまり模倣が発動されることが明らかにされた(図1).
この研究により,中脳の水道周囲灰白質から皮質の感覚運動野HVCへのドーパミンの出力が歌の模倣学習の発動において重要な役割をはたすことが明らかにされた.なぜ,水道周囲灰白質が適切な模倣の対象であるオスの成鳥の歌に対してのみ強い神経活動を起こすのかについてはいまだ不明であるが,水道周囲灰白質からHVCへとドーパミンが出力され,聴覚野からHVCへの情報の伝達に可塑的な変化がひき起こされることにより,模倣学習が発動されると考えられた.
複雑な運動の模倣学習は,模倣の対象となる行動の記憶のみで完了するわけではなく,そののち,長い時間をかけて練習されなければ達成されない9).この試行錯誤をとおした模倣の完了には,HVCの下流の大脳基底核におけるドーパミンのシグナル伝達が重要な役割をはたすことが知られ10),そのあいだ,HVCにおいて伝達されるシグナルにもさらなる変化が生じ11)(新着論文レビュー でも掲載),最終的に歌の運動のタイミングの情報を符号化するようになる12,13)(文献12) は 新着論文レビュー でも掲載,文献13) は 新着論文レビュー でも掲載).また,模倣学習の完了までには,聴覚野も模倣の対象となる記憶を保存するなど重要な役割をはたすことが知られており7,14),模倣学習をささえる神経回路の全貌はいまだ明らかにされていない.
スズメ亜目の鳥は模倣能力のほかにもさまざまな能力をヒトと共有している.たとえば,スズメ亜目の鳥は求愛の際に,歌,ダンス,巣の装飾など,ヒトの芸術に似た美しい行動をとる.また,スズメ亜目の鳥は模倣をとおして獲得した文法的な発声パターンを用いてコミュニケーションをとるめずらしい動物であり,HVCなど歌をつかさどる神経回路には,ヒトの言語をつかさどる神経回路との遺伝的および機能的な類似点がみつかりつつある15,16).さらに,スズメ亜目の鳥はきわめて社会的な動物で,特定の他者とのあいだにきわめて強固な社会的な結合を形成する.なぜ,ヒトとスズメ亜目の鳥というはなれた種がこれらのめずらしい能力を共有するのかは不明であるが,この研究において明らかにされた中脳から運動をつかさどる皮質へのドーパミンの出力は,ヒトなどの霊長目においても特別に発達しているという報告もあるため17),ヒトとスズメ亜目の鳥はそれぞれ独自にこの神経回路を進化させ,模倣能力を発達させることにより特殊な能力を獲得した可能性がある.今後,模倣能力をささえる神経回路を解明することにより,スズメ亜目の鳥とヒトの芸術,言語,社会的な行動の起源が明らかにされるかもしれない.
略歴:2013年 東京大学大学院人文社会系研究科 修了,同年 同 特任研究員,同年 米国Duke大学Postdoctoral Associateを経て,2018年より東北大学大学院生命科学研究科 助教.
研究テーマ:スズメ亜目の鳥が示す模倣能力や芸術,言語,社会的な行動をささえる神経機構.
関心事:なぜ,スズメ亜目の鳥はヒトとさまざまな能力を共有するのか.
Richard Mooney
米国Duke大学 教授.
研究室URL:https://www.neuro.duke.edu/mooney-lab
© 2018 田中雅史・Richard Mooney Licensed under CC 表示 2.1 日本
(米国Duke大学Department of Neurobiology)
email:田中雅史
DOI: 10.7875/first.author.2018.106
A mesocortical dopamine circuit enables the cultural transmission of vocal behaviour.
Masashi Tanaka, Fangmiao Sun, Yulong Li, Richard Mooney
Nature, 563, 117-120 (2018)
要 約
模倣は,規範的なモデルとなる他者の行動を記憶し自らの行動により正確に再現することを可能にする,きわめて効率的な学習能力である.しかし,脳がどのように適切な対象を選び模倣学習を発動させるのか,その機構については不明であった.この研究において,筆者らは,歌の正確な伝承能力をもつキンカチョウという鳥に着目し,成鳥の歌を聞いて模倣を開始するときの幼いキンカチョウの脳について調べた.その結果,中脳の水道周囲灰白質に存在するニューロンが,成鳥から歌いかけられたときにのみ強い神経活動を示し,歌のさえずりを制御する皮質の感覚運動野HVCへとドーパミンを出力することが明らかにされた.幼鳥が歌を聞いているときにHVCにおけるドーパミンのはたらきを阻害するとその歌の模倣は発動されず,逆に,HVCへのドーパミンの出力をひき起こすと通常は模倣されないはずのスピーカーから流れた歌によっても模倣学習が発動された.ドーパミンは幼鳥のHVCの神経活動に可塑的な変化をひき起こすとともに,幼鳥の歌を成熟化させる作用をもつことも明らかにされ,歌の模倣学習の発動にはこの運動をつかさどる皮質へと出力されるドーパミンが重要な役割をはたすことが示された.
はじめに
模倣は,ヒトなど霊長目や一部の鳥のほかにはごくかぎられた動物のみがもつ,きわめてめずらしい学習能力である1).模倣の能力をもつ動物は世代をこえてすぐれた行動を伝承することにより,それぞれに固有の文化や技術を発展させることができる.しかし,外界のあらゆる対象を模倣していては安定した技術や文化の継承はできないであろう.ヒトは適切な対象を選択しその行動のみを模倣することができるが,脳がどのようにして模倣を発動させるのか,その詳細な機構は謎につつまれている.
歌をさえずる鳥であるスズメ亜目の一種のキンカチョウは,親の歌を正確に模倣し,その歌を子孫へ伝え世代をこえた文化として歌を伝承する能力をもつ2).キンカチョウは幼少期に出会った成熟した鳥の歌を記憶し,数十日かけてくり返し練習することにより模倣を完了する.歌の模倣においては社会的な相互作用が重要であることが知られ,通常,成鳥の歌をスピーカーから流しただけでは模倣は発動されない3).この研究において,筆者らは,キンカチョウの歌の模倣を制御する脳の機能について調べるため,成熟した歌のさえずりに必要不可欠であることが知られる皮質の感覚運動野HVCに着目した4,5).HVCは下流の運動野,大脳基底核,聴覚野へと運動のシグナルを送ることにより歌のさえずりを制御しているが,近年の研究により,歌の学習にも重要な役割をはたすことが明らかになりはじめている6,7).そこで,模倣学習を開始するころの幼いキンカチョウのHVCにトレーサーを注入したところ,HVCは中脳の水道周囲灰白質という領域から密なドーパミンの入力をうけることが明らかにされた.
1.水道周囲灰白質は適切な模倣の対象に応答してHVCへとドーパミンを出力する
水道周囲灰白質の機能について知るため,自由行動下の幼いキンカチョウの水道周囲灰白質にテトロード電極を埋め込み神経活動を記録した.その結果,水道周囲灰白質に存在するニューロンの多くは適切な模倣の対象であるオスの成鳥から歌いかけられたときにのみ強い神経活動を起こすことが明らかにされた.同じ歌をスピーカーから流しただけでは顕著な応答はみられず,また,歌わないオスやメスの成鳥に出会ったとき,あるいは,メスの成鳥がいる条件にてオスの歌をスピーカーから流したときには,強い応答は起こらなかった.
水道周囲灰白質に存在するニューロンの一部はドーパミン作動性ニューロンであり,神経活動によりHVCへとドーパミンを出力する.HVCへのドーパミンの出力を検出するため,ドーパミンのセンサーであるGRABDA 8) をHVCのニューロンに発現させ,2光子顕微鏡により観察した.すると,オスの成鳥から歌いかけられたときにのみ,HVCへと多量のドーパミンが出力されることがわかった.このドーパミンの出力は,水道周囲灰白質のドーパミン作動性ニューロンを薬剤を用いて破壊することにより消失したため,HVCへのドーパミンの出力の大部分は水道周囲灰白質に由来すると考えられた.これらの結果から,水道周囲灰白質は適切な模倣の対象であるオスの成鳥の歌にのみ選択的に応答し,HVCへとドーパミンを出力することが明らかにされた.
2.HVCへのドーパミンの出力を操作することにより模倣学習を制御できる
HVCへと出力されるドーパミンが模倣にはたす機能について調べるため,幼いキンカチョウが親の歌を聞いて記憶しはじめる時期に,薬剤を用いてHVCのドーパミン作動性ニューロンを破壊したところ,その歌を模倣できなくなることがわかった.一方,幼鳥が親の歌を十分に記憶したであろうころに薬剤を用いてHVCのドーパミン作動性ニューロンを破壊したところ,幼鳥は問題なく親の歌を模倣した.したがって,HVCへと出力されるドーパミンは,模倣学習においても最初期の,歌を記憶する時期に必要であることが示唆された.
幼いころに親から隔離された幼鳥が,はじめて出会った成鳥から歌いかけられたときに,ドーパミン受容体の阻害剤を用いてHVCにおけるドーパミンのシグナル伝達を遮断したところ,幼鳥は歌を模倣できなくなることがわかった.一方,成鳥と出会って歌を聞きおえたのちにHVCにおけるドーパミンのシグナル伝達を遮断しても,幼鳥は問題なく歌を模倣した.したがって,HVCへと出力されるドーパミンは,成鳥がさえずる歌を模倣のため記憶するときに重要な役割をはたすことが示された.
もし,ドーパミンが適切な模倣の対象である成鳥が歌っているという情報をHVCに伝達することによりその歌の記憶を助けているのであれば,人工的にHVCにドーパミンを出力させることにより,通常は模倣しないスピーカーから流した歌に対しても模倣学習を発動させることができるかもしれない.そこで,水道周囲灰白質のニューロンに光により神経活動を駆動できるチャネルロドプシン2を発現させ,HVCに光を照射することにより水道周囲灰白質からHVCへのドーパミンの出力を駆動したところ,スピーカーから流した歌に対しても模倣が発動された.これらの結果から,HVCへと出力されたドーパミンはそのとき聞こえた歌に対する模倣学習を発動させることが明らかにされた.
3.ドーパミンはHVCに可塑的な変化をひき起こす
ドーパミンはHVCに出力されることによりどのように模倣学習を発動させるのだろうか? 幼鳥のHVCにおいて神経活動を記録したところ,成鳥から歌いかけられたあとには,HVCにおいてときおり自発的に生じるバースト発火がより頻繁に観察されるようになった.HVCにおけるバースト発火は,おもにnucleus interfacialis(NIf)とよばれる聴覚野6) からの入力により駆動されることが知られているため,バースト発火の増大からNIfからHVCへと伝達される聴覚のシグナルが増強されたことが示唆された.そこで,スピーカーから歌を流して幼鳥のHVCへと伝達される聴覚のシグナルについて調べたところ,その歌を成鳥から歌いかけられるまでは明瞭な応答がみられなかったのに対し,成鳥から歌いかけられたあとには,あたかもその歌を記憶したかのように,歌の特定の要素に対する応答が出現した.したがって,ドーパミンはHVCの神経活動に可塑的な変化をひき起こし,成鳥の歌の表象を記録することにより模倣を発動させると考えられた.
4.HVCの可塑的な変化とともに幼鳥の模倣学習がはじまる
HVCにおける神経活動は運動野や大脳基底核に伝達されることにより歌のさえずりを制御する4).したがって,ドーパミンの出力によりひき起こされたHVCにおける神経活動の可塑的な変化は,ただちに幼鳥の歌を変化させ模倣を発動させる可能性がある.事実,成鳥から歌いかけられたあとの幼鳥の歌を解析したところ,その歌には時間的および周波数的に成鳥の歌に近い複雑な構造がみられた.HVCにおいてドーパミンのシグナル伝達を遮断した状態においては,成鳥から歌いかけられたあとの幼鳥の歌の変化もHVCにおける神経活動の可塑的な変化も起こらなかった.したがって,成鳥から歌いかけられると,幼鳥のHVCへと出力されたドーパミンの作用によりHVCの神経活動に可塑的な変化がひき起こされると同時に,早くも幼鳥のさえずりの制御がはじまり模倣が発動されることが明らかにされた(図1).
おわりに
この研究により,中脳の水道周囲灰白質から皮質の感覚運動野HVCへのドーパミンの出力が歌の模倣学習の発動において重要な役割をはたすことが明らかにされた.なぜ,水道周囲灰白質が適切な模倣の対象であるオスの成鳥の歌に対してのみ強い神経活動を起こすのかについてはいまだ不明であるが,水道周囲灰白質からHVCへとドーパミンが出力され,聴覚野からHVCへの情報の伝達に可塑的な変化がひき起こされることにより,模倣学習が発動されると考えられた.
複雑な運動の模倣学習は,模倣の対象となる行動の記憶のみで完了するわけではなく,そののち,長い時間をかけて練習されなければ達成されない9).この試行錯誤をとおした模倣の完了には,HVCの下流の大脳基底核におけるドーパミンのシグナル伝達が重要な役割をはたすことが知られ10),そのあいだ,HVCにおいて伝達されるシグナルにもさらなる変化が生じ11)(新着論文レビュー でも掲載),最終的に歌の運動のタイミングの情報を符号化するようになる12,13)(文献12) は 新着論文レビュー でも掲載,文献13) は 新着論文レビュー でも掲載).また,模倣学習の完了までには,聴覚野も模倣の対象となる記憶を保存するなど重要な役割をはたすことが知られており7,14),模倣学習をささえる神経回路の全貌はいまだ明らかにされていない.
スズメ亜目の鳥は模倣能力のほかにもさまざまな能力をヒトと共有している.たとえば,スズメ亜目の鳥は求愛の際に,歌,ダンス,巣の装飾など,ヒトの芸術に似た美しい行動をとる.また,スズメ亜目の鳥は模倣をとおして獲得した文法的な発声パターンを用いてコミュニケーションをとるめずらしい動物であり,HVCなど歌をつかさどる神経回路には,ヒトの言語をつかさどる神経回路との遺伝的および機能的な類似点がみつかりつつある15,16).さらに,スズメ亜目の鳥はきわめて社会的な動物で,特定の他者とのあいだにきわめて強固な社会的な結合を形成する.なぜ,ヒトとスズメ亜目の鳥というはなれた種がこれらのめずらしい能力を共有するのかは不明であるが,この研究において明らかにされた中脳から運動をつかさどる皮質へのドーパミンの出力は,ヒトなどの霊長目においても特別に発達しているという報告もあるため17),ヒトとスズメ亜目の鳥はそれぞれ独自にこの神経回路を進化させ,模倣能力を発達させることにより特殊な能力を獲得した可能性がある.今後,模倣能力をささえる神経回路を解明することにより,スズメ亜目の鳥とヒトの芸術,言語,社会的な行動の起源が明らかにされるかもしれない.
文 献
- Whiten, A.: Social learning and culture in child and chimpanzee. Annu. Rev. Psychol., 68, 129-154 (2017)[PubMed]
- Marler, P. & Tamura, M.: Culturally transmitted patterns of vocal behavior in sparrows. Science, 146, 1483-1486 (1964)[PubMed]
- Chen, Y., Matheson, L. E. & Sakata, J. T.: Mechanisms underlying the social enhancement of vocal learning in songbirds. Proc. Natl Acad. Sci. USA, 113, 6641-6646 (2016)[PubMed]
- Aronov, D., Andalman, A. S. & Fee, M. S.: A specialized forebrain circuit for vocal babbling in the juvenile songbird. Science, 320, 630-634 (2008)[PubMed]
- Long, M. S. & Fee, M. S.: Using temperature to analyse temporal dynamics in the songbird motor pathway. Nature, 456, 189-194 (2008)[PubMed]
- Roberts, T. F., Gobes, S. M., Murugan, M. et al.: Motor circuits are required to encode a sensory model for imitative learning. Nat. Neurosci., 15, 1454-1459 (2012)[PubMed]
- Roberts, T. F., Hisey, E., Tanaka, M. et al.: Identification of a motor-to-auditory pathway important for vocal learning. Nat. Neurosci., 20, 978-986 (2017)[PubMed]
- Sun, F., Zeng, J., Jing, M. et al.: A genetically encoded fluorescent sensor enables rapid and specific detection of dopamine in flies, fish, and mice. Cell, 174, 481-496 (2018)[PubMed]
- Hayase, S., Wang, H., Ohgushi, E. et al.: Vocal practice regulates singing activity-dependent genes underlying age-independent vocal learning in songbirds. PLoS Biol., 16, e2006537 (2018)[PubMed]
- Hisey, E., Kearney, M. G. & Mooney, R.: A common neural circuit mechanism for internally guided and externally reinforced forms of motor learning. Nat. Neurosci., 21, 589-597 (2018)[PubMed]
- Okubo, T. S., Mackevicius, E. L., Payne, H. L. et al.: Growth and splitting of neural sequences in songbird vocal development. Nature, 528, 352-357 (2015)[PubMed] [新着論文レビュー]
- Hamaguchi, K., Tanaka, M. & Mooney, R.: A distributed recurrent network contributes to temporally precise vocalizations. Neuron, 91, 680-693 (2016)[PubMed] [新着論文レビュー]
- Lynch, G. F., Okubo, T. S., Hanuschkin, A. et al.: Rhythmic continuous-time coding in the songbird analog of vocal motor cortex. Neuron, 90, 877-892 (2016)[PubMed] [新着論文レビュー]
- Yanagihara, S. & Yazaki-Sugiyama, Y.: Auditory experience-dependent cortical circuit shaping for memory formation in bird song learning. Nat. Commun., 7, 11946 (2016)[PubMed]
- Pfenning, A. R., Hara, E., Whitney, O. et al.: Convergent transcriptional specializations in the brains of humans and song-learning birds. Science, 346, 1256846 (2014)[PubMed]
- Long, M. A., Katlowitz, K. A., Svirsky, M. A. et al.: Functional segregation of cortical regions underlying speech timing and articulation. Neuron, 89, 1187-1193 (2016)[PubMed]
- Berger, B., Gaspar, P. & Verney, C.: Dopaminergic innervation of the cerebral cortex: unexpected differences between rodents and primates. Trends Neurosci., 14, 21-27 (1991)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2013年 東京大学大学院人文社会系研究科 修了,同年 同 特任研究員,同年 米国Duke大学Postdoctoral Associateを経て,2018年より東北大学大学院生命科学研究科 助教.
研究テーマ:スズメ亜目の鳥が示す模倣能力や芸術,言語,社会的な行動をささえる神経機構.
関心事:なぜ,スズメ亜目の鳥はヒトとさまざまな能力を共有するのか.
Richard Mooney
米国Duke大学 教授.
研究室URL:https://www.neuro.duke.edu/mooney-lab
© 2018 田中雅史・Richard Mooney Licensed under CC 表示 2.1 日本
