小鳥の歌の学習における神経シークエンスの成長および分裂
大久保達夫・Michale S. Fee
(米国Massachusetts Institute of Technology,Department of Brain and Cognitive Sciences,McGovern Institute for Brain Research)
email:大久保達夫
DOI: 10.7875/first.author.2015.142
Growth and splitting of neural sequences in songbird vocal development.
Tatsuo S. Okubo, Emily L. Mackevicius, Hannah L. Payne, Galen F. Lynch, Michale S. Fee
Nature, 528, 352-357 (2015)
近年,複雑な運動を生み出す脳の機構として神経シークエンスが注目されている.しかし,学習中の動物の脳においてどのように神経シークエンスが形成されているのかについては解明されていない部分が多い.この研究においては,複雑な運動の一例である小鳥の歌の学習をモデルとしてこの問題に取り組んだ.先行研究により,成熟した小鳥が歌っている際には,運動前野において神経シークエンスが観察されることが示されていた.そこで,歌を学習している幼い小鳥において神経シークエンスがどのように形成されるかについて調べるため,さまざまな発達の段階において運動前野における神経活動を記録した.その結果,シラブルの原型が形成される際には,まず,神経シークエンスが短いものから長いものへと成長し,つづいて,神経シークエンスが2つに分裂することにより2種類のシラブルが形成されることがわかった.また,このような神経シークエンスの分裂はさまざまなシラブルの形成においても観察されたことから,複雑な歌を学習するための普遍的な機構であると考えられた.
楽器の演奏やスポーツといったわれわれの複雑な運動の多くは細かい動作を正確なタイミングでつぎつぎに行うことにより成り立っている.脳はいったいどのようにしてこのような複雑な運動を生み出すのだろうか? そして,これらの運動を学習する際に,脳においてはどのような変化が起こっているのだろうか? 複雑な行動の際に,個々のニューロンは特定の動作のときのみ,しかも,その動作のなかでもさらに特定のタイミングのときのみ,活動していることが多い.そして,異なるニューロンはそれぞれ異なるタイミングで活動するため,ニューロンの集団としては早いタイミングで活動するニューロンから遅いタイミングで活動するニューロンにいたるまで順番に活動する1,2).ここでは,そのような活動を総称して“神経シークエンス”とよぶ.神経シークエンスは多くの動物,しかも,脳のさまざまな部位においてみられるが3),先行研究の多くは学習をすませた動物から神経活動を記録しており,学習の過程においてどのように神経シークエンスが形成されるかという問題の解明ははじまったばかりであった2).
小鳥の歌の学習はこの問題を明らかにするのに適したモデルである.第1に,小鳥の歌は複数のシラブルがつぎからつぎへとタイミングよくくり出される複雑な運動である.第2に,小鳥において親の歌をまねるという行動は自然にみられ特別なトレーニングの必要はない.なおかつ,幼い小鳥が2カ月にわたり練習した歌をすべて録音することにより学習の過程を詳細に解析することが可能である4).第3に,成熟した小鳥が歌っているときには運動前野であるHVCとよばれる部位には神経シークエンスがみられる.すなわち,運動前野における個々のニューロンのほとんどは特定のシラブルの特定のタイミングでのみ活動しそれ以外はまったく活動しない.そして,異なるニューロンは異なるタイミングで活動するため,ニューロンの集団としては神経シークエンスを形成している5,6)(図1a).したがって,歌を学習している幼い小鳥の運動前野において神経活動を記録すれば,学習をしている動物の脳においてどのように神経シークエンスが形成されるかという問題に答えることができると期待された.
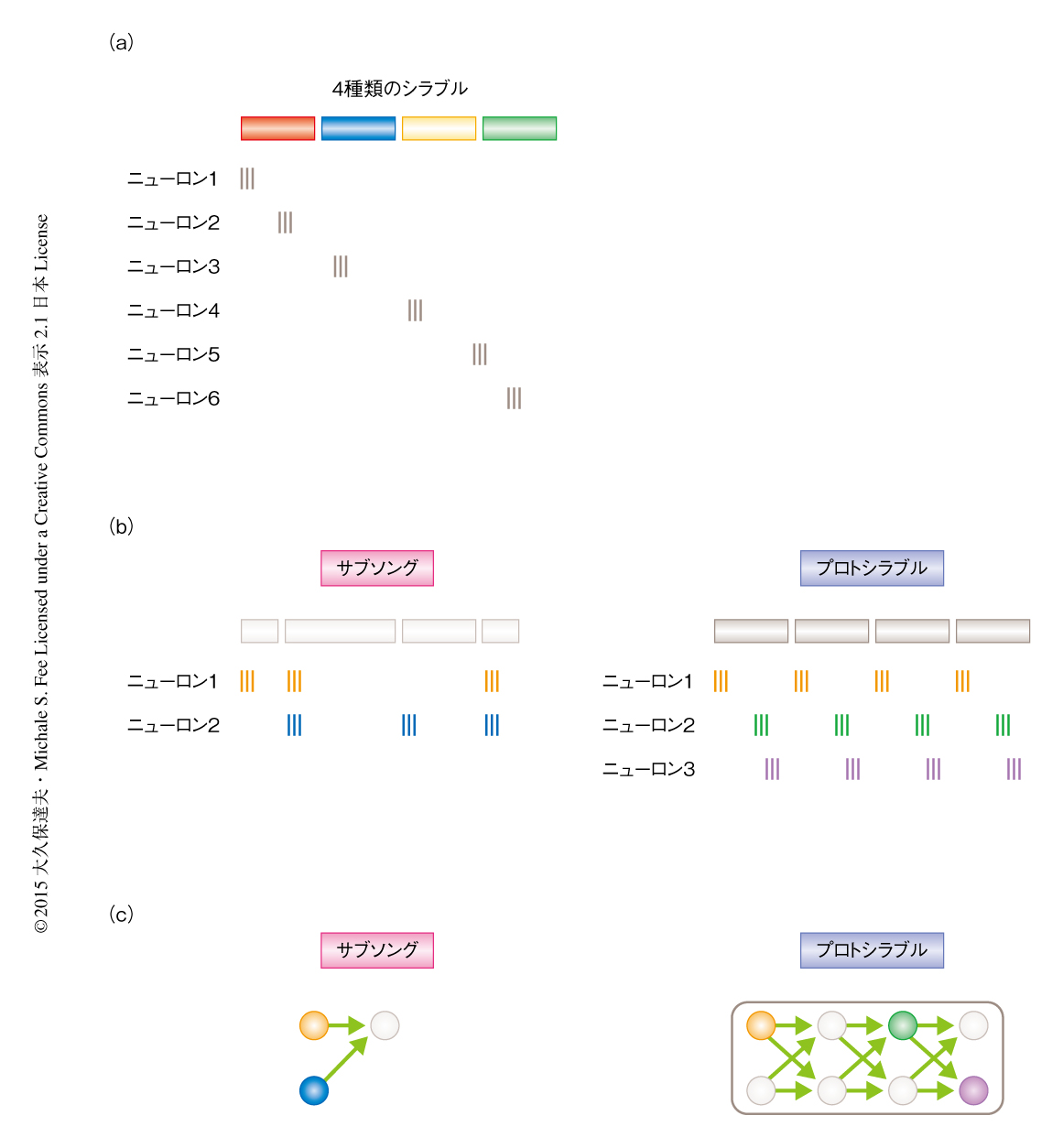
はじめに,歌の学習におけるはじめの段階であるサブソングに着目した.サブソングはヒトの赤ん坊における喃語に相当し,シラブルの長さはランダムで,音もはっきりとした構造をもたない.サブソングの段階の幼い小鳥から神経活動を記録するため,小型モーターを搭載したマイクロドライブを用い,運動前野の投射ニューロンから単一のニューロンにおいて細胞外記録を行った.その結果,先行研究において運動前野はサブソングに必要ないとされていたにもかかわらず7),サブソングを歌っている際の運動前野において歌に関連する神経活動が記録された.そのうち約半数のニューロンはシラブルのタイミングと時間的に同期していなかったが,残りの約半数のニューロンはシラブルの特定のタイミングにおいて活動し,かつ,シラブルの開始のときに活動するものがほとんどであった(図1b).この結果から,サブソングの段階では神経シークエンスが存在したとしてもそれはごく短いものであり,シラブルの開始からすぐに活動は途絶えてしまうと解釈された.
歌の学習が進行するにつれ,シラブルの長さがランダムなサブソングから,しだいに約100ミリ秒の決まった長さをもつシラブルの原型(プロトシラブル)が現われるようになる4).歌に構造が現われるのと並行して,運動前野における神経活動にも時間的な構造が現われはじめた.すなわち,多くのニューロンは約100ミリ秒ごとにリズミカルな活動をくり返し,かつ,異なるニューロンはシラブルに対し異なるタイミングで活動した(図1b).これはまさに神経シークエンスであり,この結果から,サブソングからプロトシラブルに移行する際に,短い神経シークエンスがより長いものへと成長することが示された(図1c).
ここまで,サブソングからひとつのプロトシラブルが形成されるまでの過程をみてきたが,成熟した小鳥の歌には複数の種類のシラブルが含まれている.どのようにひとつのプロトシラブルから複数のシラブルが形成され,その際,運動前野における神経活動はどのように変化するのだろうか? 新たなシラブルを学習するごとにそのつど神経シークエンスは成長するのだろうか,それとも,プロトシラブルの形成に使用された神経シークエンスが分裂することにより新たなシラブルが形成されるのであろうか? もし後者であるなら,神経シークエンスが分裂している途中においては1つのニューロンが2つのシラブルに共有され,両方のシラブルにおいて活動すると予想された.この可能性を探るため,幼い小鳥において1つのプロトシラブルから2種類のシラブルが形成される数週間の過程において運動前野における神経活動を記録した.
形成された2つのシラブルは学習の初期においては音響的に似かよっていたが,しだいにその差異は拡大した.そして,2つのシラブルが形成された直後には,多くのニューロンが2つのシラブルにおいて活動していた(図2a).しかも,これら共有されたニューロンは単に2つのシラブルにおいて活動していただけでなく,2つのシラブルにおいて似たタイミングで活動していた.たとえば,一方のシラブルのおわりのほうで活動していたニューロンは,他方のシラブルにおいてもおわりのほうで活動していた.この結果から,学習の初期においては2つのシラブルをコードするニューロンの多くは重複しており,これらの共有されたニューロンは2つのシラブルのあいだで共有された神経シークエンスを形成することが示唆された.
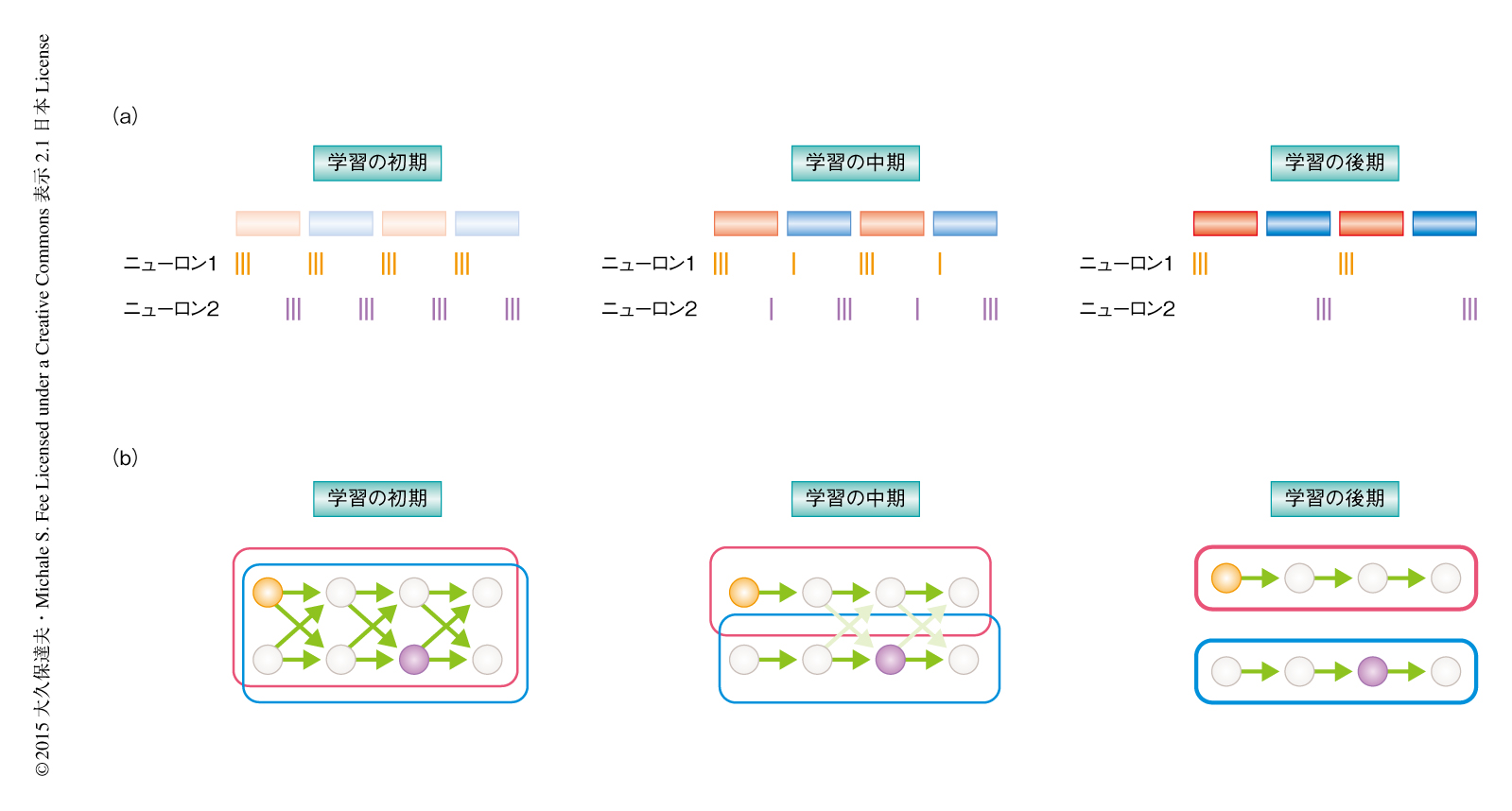
シラブルの形成が進行するにつれ2つのシラブルの音響的な違いは大きくなったが,それと並行して,共有されるニューロンの割合は減少し,1つのシラブルでのみ活動するニューロンの割合が増加した(図2a).以上の結果は,プロトシラブルをコードする神経シークエンスが学習の進行とともに分裂して2つに分かれることにより2種類の新しいシラブルが形成されるという仮説と合致した(図2b).
ここまでの例においては,1つのプロトシラブルから2つのシラブルが形成され,それらが交互にくり返されるというかたちでシラブルの数が増加していた.しかし,これまでの行動学的な研究より,これ以外の方法を用いても複数の種類のシラブルが形成されることがわかっている8,9).たとえば,歌の開始時や終了時に新たなシラブルが現われることもあれば8),モチーフを構成する複数のシラブルがほぼ同時に形成されることもある9).神経シークエンスの分裂はこれらの方法にもかかわっているのだろうか? それについて調べるため,これらの方法によりシラブルを学習した個体において長期間にわたり運動前野における神経活動を記録した.その結果,これらの個体においても学習の初期においては複数のシラブルにて活動するニューロンが多くみられ,シラブルの形成につれこれら共有されるニューロンの割合は減少した.これは,さきに述べた,神経シークエンスの分裂という解釈と合致するものであった.また,ほかにも,いったん分裂した神経シークエンスがふたたび分裂してより多くのシラブルを形成することも観察された.この方法を使用すれば,1つのプロトシラブルから2種類のシラブルを形成するだけでなく,それらをさらに分裂させることにより4種類のシラブルを形成することができる.以上の結果から,神経シークエンスの分裂は小鳥がシラブルを学習するのに使用するさまざまな方法にかかわっており,小鳥がシラブルを学習するうえで普遍的な機構であると考えらえた.
小鳥の歌の学習は古代より多くの人々の興味をひいてきた.わが国においても,ウグイスやメジロを用いて歌の優劣を競う“鳴き合わせ”といった伝統がある.一方で,小鳥の歌の学習の科学的な研究は脳科学の分野においてもすぐれた成果をもたらしたが,学習の過程において脳の活動がどのように変化するかという問題に直接的に挑んだ研究は少なかった.この研究では,歌を学習している幼い小鳥のさまざまな発達の段階において,運動前野における神経活動を個々のニューロンのレベルにて調べることによりシラブルが形成される機構を探った.その結果,神経シークエンスの成長および分裂が重要な機構であることが示唆された.
この機構はニューロンどうしが連結した神経回路において実際に起こりうるのか,その妥当性を探るため,先行研究において提案されたモデルに10,11),いくつかの要素をくわえて神経回路ネットワークの数理的なシミュレーションを行った.その結果,実験の結果と似た挙動がみられた.このシミュレーションにおいて仮定した要素が正しいかどうかは,今後の実験によるさらなる検証が必要である.
また,残された課題もある.たとえば,今回の実験においては個々のニューロンから記録できるのは5~15分間であるため,モデルを厳密に証明するためには脳の活動のイメージングなどにより同じニューロンを数週間にわたり追跡する手法2) を開発する必要がある.また,今回の結果は,必ずしもすべてのシラブルが神経シークエンスの分裂により形成されていることを示すものではなく,これとは別の機構がはたらいている可能性もある.
最後に,なぜ多くの種類のシラブルを学習するのに神経シークエンスの分裂が用いられているのか考察したい.ミクロなスケールでは,遺伝子の多様性を生み出す機構として,1つの遺伝子が重複により2つになり,それぞれに別の変異が積み重なることにより2つの異なる遺伝子が生じることがあると考えられている.同様に,脳においても,プロトシラブルを形成する神経シークエンスが,まず,シラブルに共通する部分を学習し,そののち,神経シークエンスを分裂させそれぞれを独立に変化させることによりシラブルの多様性を生み出している可能性が考えられる.ヒトを含めた動物の行動は多様な運動パターンにより成り立っており,小鳥の歌の学習において示された神経シークエンスの成長および分裂と似たような機構が,広く運動の学習の全般において使用されていることが期待される.
略歴:2015年 米国Massachusetts Institute of Technology修了,同年より米国Harvard Medical School博士研究員.
抱負:脳がどのようにして動物の自然な行動を生み出すか,電気生理学をはじめとしたさまざまな方法を駆使することにより総合的に理解していきたい.
Michale S. Fee
米国Massachusetts Institute of Technology教授.
研究室URL:http://web.mit.edu/feelab/
© 2015 大久保達夫・Michale S. Fee Licensed under CC 表示 2.1 日本
(米国Massachusetts Institute of Technology,Department of Brain and Cognitive Sciences,McGovern Institute for Brain Research)
email:大久保達夫
DOI: 10.7875/first.author.2015.142
Growth and splitting of neural sequences in songbird vocal development.
Tatsuo S. Okubo, Emily L. Mackevicius, Hannah L. Payne, Galen F. Lynch, Michale S. Fee
Nature, 528, 352-357 (2015)
要 約
近年,複雑な運動を生み出す脳の機構として神経シークエンスが注目されている.しかし,学習中の動物の脳においてどのように神経シークエンスが形成されているのかについては解明されていない部分が多い.この研究においては,複雑な運動の一例である小鳥の歌の学習をモデルとしてこの問題に取り組んだ.先行研究により,成熟した小鳥が歌っている際には,運動前野において神経シークエンスが観察されることが示されていた.そこで,歌を学習している幼い小鳥において神経シークエンスがどのように形成されるかについて調べるため,さまざまな発達の段階において運動前野における神経活動を記録した.その結果,シラブルの原型が形成される際には,まず,神経シークエンスが短いものから長いものへと成長し,つづいて,神経シークエンスが2つに分裂することにより2種類のシラブルが形成されることがわかった.また,このような神経シークエンスの分裂はさまざまなシラブルの形成においても観察されたことから,複雑な歌を学習するための普遍的な機構であると考えられた.
はじめに
楽器の演奏やスポーツといったわれわれの複雑な運動の多くは細かい動作を正確なタイミングでつぎつぎに行うことにより成り立っている.脳はいったいどのようにしてこのような複雑な運動を生み出すのだろうか? そして,これらの運動を学習する際に,脳においてはどのような変化が起こっているのだろうか? 複雑な行動の際に,個々のニューロンは特定の動作のときのみ,しかも,その動作のなかでもさらに特定のタイミングのときのみ,活動していることが多い.そして,異なるニューロンはそれぞれ異なるタイミングで活動するため,ニューロンの集団としては早いタイミングで活動するニューロンから遅いタイミングで活動するニューロンにいたるまで順番に活動する1,2).ここでは,そのような活動を総称して“神経シークエンス”とよぶ.神経シークエンスは多くの動物,しかも,脳のさまざまな部位においてみられるが3),先行研究の多くは学習をすませた動物から神経活動を記録しており,学習の過程においてどのように神経シークエンスが形成されるかという問題の解明ははじまったばかりであった2).
小鳥の歌の学習はこの問題を明らかにするのに適したモデルである.第1に,小鳥の歌は複数のシラブルがつぎからつぎへとタイミングよくくり出される複雑な運動である.第2に,小鳥において親の歌をまねるという行動は自然にみられ特別なトレーニングの必要はない.なおかつ,幼い小鳥が2カ月にわたり練習した歌をすべて録音することにより学習の過程を詳細に解析することが可能である4).第3に,成熟した小鳥が歌っているときには運動前野であるHVCとよばれる部位には神経シークエンスがみられる.すなわち,運動前野における個々のニューロンのほとんどは特定のシラブルの特定のタイミングでのみ活動しそれ以外はまったく活動しない.そして,異なるニューロンは異なるタイミングで活動するため,ニューロンの集団としては神経シークエンスを形成している5,6)(図1a).したがって,歌を学習している幼い小鳥の運動前野において神経活動を記録すれば,学習をしている動物の脳においてどのように神経シークエンスが形成されるかという問題に答えることができると期待された.
1.神経シークエンスが成長することによりシラブルの原型が形成される
はじめに,歌の学習におけるはじめの段階であるサブソングに着目した.サブソングはヒトの赤ん坊における喃語に相当し,シラブルの長さはランダムで,音もはっきりとした構造をもたない.サブソングの段階の幼い小鳥から神経活動を記録するため,小型モーターを搭載したマイクロドライブを用い,運動前野の投射ニューロンから単一のニューロンにおいて細胞外記録を行った.その結果,先行研究において運動前野はサブソングに必要ないとされていたにもかかわらず7),サブソングを歌っている際の運動前野において歌に関連する神経活動が記録された.そのうち約半数のニューロンはシラブルのタイミングと時間的に同期していなかったが,残りの約半数のニューロンはシラブルの特定のタイミングにおいて活動し,かつ,シラブルの開始のときに活動するものがほとんどであった(図1b).この結果から,サブソングの段階では神経シークエンスが存在したとしてもそれはごく短いものであり,シラブルの開始からすぐに活動は途絶えてしまうと解釈された.
歌の学習が進行するにつれ,シラブルの長さがランダムなサブソングから,しだいに約100ミリ秒の決まった長さをもつシラブルの原型(プロトシラブル)が現われるようになる4).歌に構造が現われるのと並行して,運動前野における神経活動にも時間的な構造が現われはじめた.すなわち,多くのニューロンは約100ミリ秒ごとにリズミカルな活動をくり返し,かつ,異なるニューロンはシラブルに対し異なるタイミングで活動した(図1b).これはまさに神経シークエンスであり,この結果から,サブソングからプロトシラブルに移行する際に,短い神経シークエンスがより長いものへと成長することが示された(図1c).
2.神経シークエンスが分裂することによりシラブルの種類が増加する
ここまで,サブソングからひとつのプロトシラブルが形成されるまでの過程をみてきたが,成熟した小鳥の歌には複数の種類のシラブルが含まれている.どのようにひとつのプロトシラブルから複数のシラブルが形成され,その際,運動前野における神経活動はどのように変化するのだろうか? 新たなシラブルを学習するごとにそのつど神経シークエンスは成長するのだろうか,それとも,プロトシラブルの形成に使用された神経シークエンスが分裂することにより新たなシラブルが形成されるのであろうか? もし後者であるなら,神経シークエンスが分裂している途中においては1つのニューロンが2つのシラブルに共有され,両方のシラブルにおいて活動すると予想された.この可能性を探るため,幼い小鳥において1つのプロトシラブルから2種類のシラブルが形成される数週間の過程において運動前野における神経活動を記録した.
形成された2つのシラブルは学習の初期においては音響的に似かよっていたが,しだいにその差異は拡大した.そして,2つのシラブルが形成された直後には,多くのニューロンが2つのシラブルにおいて活動していた(図2a).しかも,これら共有されたニューロンは単に2つのシラブルにおいて活動していただけでなく,2つのシラブルにおいて似たタイミングで活動していた.たとえば,一方のシラブルのおわりのほうで活動していたニューロンは,他方のシラブルにおいてもおわりのほうで活動していた.この結果から,学習の初期においては2つのシラブルをコードするニューロンの多くは重複しており,これらの共有されたニューロンは2つのシラブルのあいだで共有された神経シークエンスを形成することが示唆された.
シラブルの形成が進行するにつれ2つのシラブルの音響的な違いは大きくなったが,それと並行して,共有されるニューロンの割合は減少し,1つのシラブルでのみ活動するニューロンの割合が増加した(図2a).以上の結果は,プロトシラブルをコードする神経シークエンスが学習の進行とともに分裂して2つに分かれることにより2種類の新しいシラブルが形成されるという仮説と合致した(図2b).
3.神経シークエンスの分裂はさまざまなシラブルの形成において観察される
ここまでの例においては,1つのプロトシラブルから2つのシラブルが形成され,それらが交互にくり返されるというかたちでシラブルの数が増加していた.しかし,これまでの行動学的な研究より,これ以外の方法を用いても複数の種類のシラブルが形成されることがわかっている8,9).たとえば,歌の開始時や終了時に新たなシラブルが現われることもあれば8),モチーフを構成する複数のシラブルがほぼ同時に形成されることもある9).神経シークエンスの分裂はこれらの方法にもかかわっているのだろうか? それについて調べるため,これらの方法によりシラブルを学習した個体において長期間にわたり運動前野における神経活動を記録した.その結果,これらの個体においても学習の初期においては複数のシラブルにて活動するニューロンが多くみられ,シラブルの形成につれこれら共有されるニューロンの割合は減少した.これは,さきに述べた,神経シークエンスの分裂という解釈と合致するものであった.また,ほかにも,いったん分裂した神経シークエンスがふたたび分裂してより多くのシラブルを形成することも観察された.この方法を使用すれば,1つのプロトシラブルから2種類のシラブルを形成するだけでなく,それらをさらに分裂させることにより4種類のシラブルを形成することができる.以上の結果から,神経シークエンスの分裂は小鳥がシラブルを学習するのに使用するさまざまな方法にかかわっており,小鳥がシラブルを学習するうえで普遍的な機構であると考えらえた.
おわりに
小鳥の歌の学習は古代より多くの人々の興味をひいてきた.わが国においても,ウグイスやメジロを用いて歌の優劣を競う“鳴き合わせ”といった伝統がある.一方で,小鳥の歌の学習の科学的な研究は脳科学の分野においてもすぐれた成果をもたらしたが,学習の過程において脳の活動がどのように変化するかという問題に直接的に挑んだ研究は少なかった.この研究では,歌を学習している幼い小鳥のさまざまな発達の段階において,運動前野における神経活動を個々のニューロンのレベルにて調べることによりシラブルが形成される機構を探った.その結果,神経シークエンスの成長および分裂が重要な機構であることが示唆された.
この機構はニューロンどうしが連結した神経回路において実際に起こりうるのか,その妥当性を探るため,先行研究において提案されたモデルに10,11),いくつかの要素をくわえて神経回路ネットワークの数理的なシミュレーションを行った.その結果,実験の結果と似た挙動がみられた.このシミュレーションにおいて仮定した要素が正しいかどうかは,今後の実験によるさらなる検証が必要である.
また,残された課題もある.たとえば,今回の実験においては個々のニューロンから記録できるのは5~15分間であるため,モデルを厳密に証明するためには脳の活動のイメージングなどにより同じニューロンを数週間にわたり追跡する手法2) を開発する必要がある.また,今回の結果は,必ずしもすべてのシラブルが神経シークエンスの分裂により形成されていることを示すものではなく,これとは別の機構がはたらいている可能性もある.
最後に,なぜ多くの種類のシラブルを学習するのに神経シークエンスの分裂が用いられているのか考察したい.ミクロなスケールでは,遺伝子の多様性を生み出す機構として,1つの遺伝子が重複により2つになり,それぞれに別の変異が積み重なることにより2つの異なる遺伝子が生じることがあると考えられている.同様に,脳においても,プロトシラブルを形成する神経シークエンスが,まず,シラブルに共通する部分を学習し,そののち,神経シークエンスを分裂させそれぞれを独立に変化させることによりシラブルの多様性を生み出している可能性が考えられる.ヒトを含めた動物の行動は多様な運動パターンにより成り立っており,小鳥の歌の学習において示された神経シークエンスの成長および分裂と似たような機構が,広く運動の学習の全般において使用されていることが期待される.
文 献
- Tanji, J.: Sequential organization of multiple movements: involvement of cortical motor areas. Annu. Rev. Neurosci., 24, 631-651 (2001)[PubMed]
- Peters, A. J., Chen, S. X. & Komiyama, T.: Emergence of reproducible spatiotemporal activity during motor learning. Nature, 510, 263-267 (2014)[PubMed]
- Buzsaki, G.: Neural syntax: cell assemblies, synapsembles, and readers. Neuron, 68, 362-385 (2010)[PubMed]
- Tchernichovski, O.: Dynamics of the vocal imitation process: How a zebra finch learns its song. Science, 291, 2564-2569 (2001)[PubMed]
- Hahnloser, R. H. R., Kozhevnikov, A. A. & Fee, M. S.: An ultra-sparse code underlies the generation of neural sequences in a songbird. Nature, 419, 65-70 (2002)[PubMed]
- Long, M. A., Jin, D. Z. & Fee, M. S.: Support for a synaptic chain model of neuronal sequence generation. Nature, 468, 394-399 (2010)[PubMed]
- Aronov, D., Andalman, A. S. & Fee, M. S.: A specialized forebrain circuit for vocal babbling in the juvenile songbird. Science, 320, 630-634 (2008)[PubMed]
- Lipkind, D., Marcus, G. F., Bemis, D. K. et al.: Stepwise acquisition of vocal combinatorial capacity in songbirds and human infants. Nature, 498, 104-108 (2013)[PubMed]
- Liu, W., Gardner, T. J. & Nottebohm, F.: Juvenile zebra finches can use multiple strategies to learn the same song. Proc. Natl. Acad. Sci. USA, 101, 18177-18182 (2004)[PubMed]
- Jun, J. K. & Jin, D. Z.: Development of neural circuitry for precise temporal sequences through spontaneous activity, axon remodeling, and synaptic plasticity. PLoS One, 2, e723 (2007)[PubMed]
- Fiete, I. R., Senn, W., Wang, C. Z. H. et al.: Spike-time-dependent plasticity and heterosynaptic competition organize networks to produce long scale-free sequences of neural activity. Neuron, 65, 563-576 (2010)[PubMed]
著者プロフィール
略歴:2015年 米国Massachusetts Institute of Technology修了,同年より米国Harvard Medical School博士研究員.
抱負:脳がどのようにして動物の自然な行動を生み出すか,電気生理学をはじめとしたさまざまな方法を駆使することにより総合的に理解していきたい.
Michale S. Fee
米国Massachusetts Institute of Technology教授.
研究室URL:http://web.mit.edu/feelab/
© 2015 大久保達夫・Michale S. Fee Licensed under CC 表示 2.1 日本
