マウスの性フェロモンESP1がメスの性行動を促進するための専用の神経回路
石井健太郎・宮道和成・東原和成
(東京大学大学院農学生命科学研究科 応用生命化学専攻生物化学研究室)
email:宮道和成,東原和成
DOI: 10.7875/first.author.2017.057
A labeled-line neural circuit for pheromone-mediated sexual behaviors in mice.
Kentaro K. Ishii, Takuya Osakada, Hiromi Mori, Nobuhiko Miyasaka, Yoshihiro Yoshihara, Kazunari Miyamichi, Kazushige Touhara
Neuron, 95, 123-137 (2017)
生物の行動,とくに本能的な行動は匂いなどの化学物質により制御される.たとえば,マウスにおいては異性,同性,天敵に由来する物質が,それぞれ,性行動,攻撃行動,忌避行動をひき起こすことが知られている.しかし,特定の感覚シグナルが適切な行動をひき起こすための神経基盤については不明であった.今回,筆者らは,単一の受容体を介して生得的にマウスの行動に影響をおよぼす性フェロモンESP1に着目し,ESP1がメスのマウスにおいて性行動の促進効果をひき起こすための神経回路を末梢から行動を制御する脳の中枢までニューロンのレベルにおいて明らかにした.また,ESP1は雌雄で異なる神経回路より処理されることも見い出された.さらに,ESP1のシグナルと天敵のシグナルを伝達する神経回路は同じ領域を介するものの,ニューロンのレベルにおいては異なっていた.これらの一連の結果から,生得的な行動を制御する感覚シグナルそれぞれには専用の神経回路が用意される可能性が示唆された.このようなシンプルな構造は,特定の感覚入力に対し生得的に適切な行動を出力するうえで好適なのかもしれない.
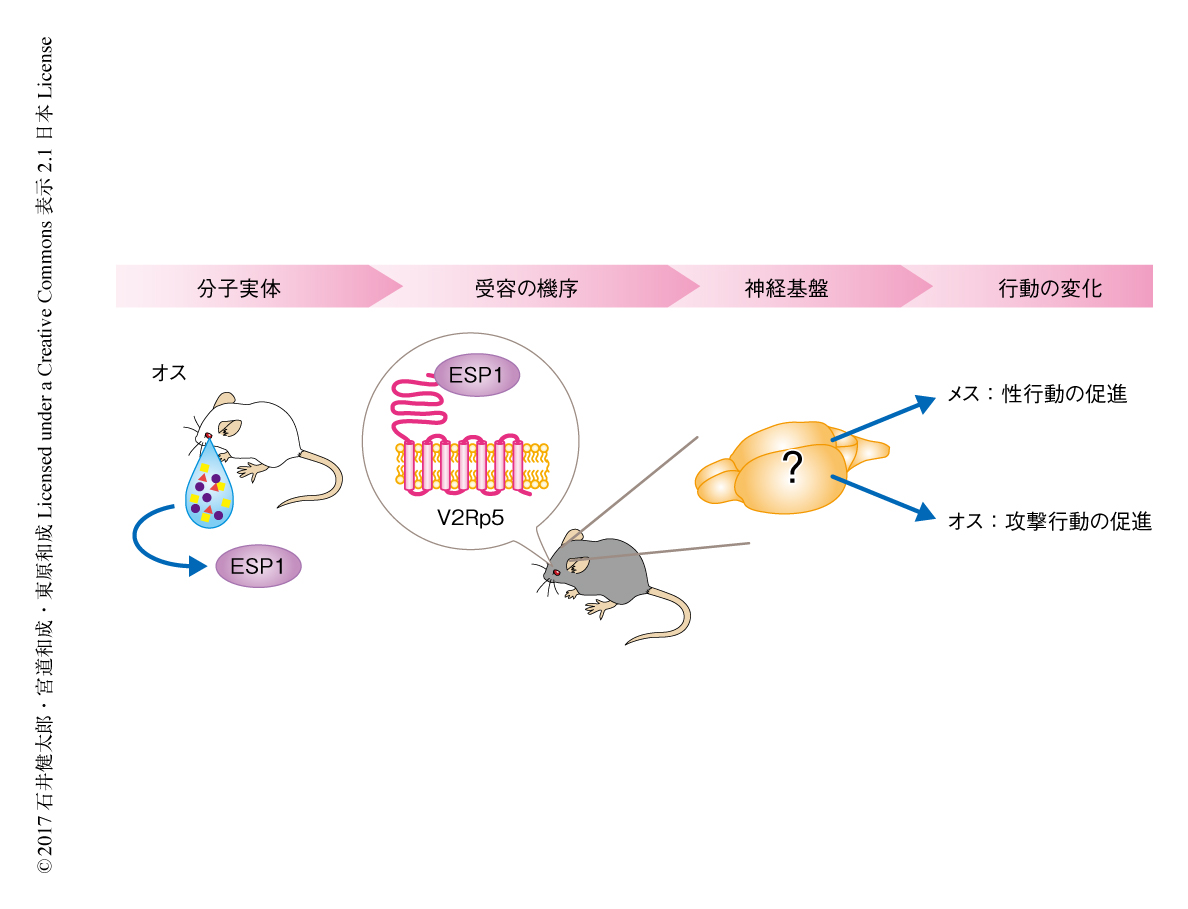
生物は外界からさまざまなシグナルを感知し,それにもとづき適切な行動を選択する.多くの場合,この過程には学習が必要であるが,なかには学習を必要とせず特定の本能的な行動をひき起こすシグナルもある.多くの生物において,匂いなどの化学物質のシグナルは本能的な行動の制御に重要な役割をはたすことが知られている.なかでも,同じ種のほかの個体に対し行動や情動の変化をひき起こす物質はフェロモンとよばれ,さまざまな生物において確認されている.たとえば,はじめて化学構造が明らかにされたフェロモンであるボンビコールは,メスのカイコガから発せられ,それだけでオスを誘引する効果をもつ1).哺乳類においてもさまざまなフェロモンが同定されており,たとえば,オスのブタの唾液に分泌され,発情状態のメスに交尾姿勢をひき起こさせる物質としてアンドロステノンが同定されている1).このように,フェロモンが“何か?”その分子実態が明らかにされつつある一方,フェロモンが“どうはたらくのか?”つまり,特定の感覚シグナルが適切な生得的な行動をひき起こすための神経基盤は哺乳類においてはほとんど明らかにされていない.この問いを解く鍵は,フェロモンのシグナルが伝達される神経回路を,末梢の器官から行動を制御する脳の中枢の領域までニューロンのレベルにおいて明らかにすることであると考えた.そのためには,シンプルな感覚入力により行動の劇的な変化をひき起こすような物質が最適である.
筆者らの研究室における先行研究により,オスのマウスの涙液に分泌されるESP1(exocrine gland-secreting peptide 1)とよばれる約7 kDaのペプチドがフェロモンとして機能することが明らかにされた2)(新着論文レビュー でも掲載).ESP1を受容したメスのマウスは,オスのマウスに対し性的な受け入れ行動であるロードシス反射を示す割合が増加する.また,ESP1はマウスの末梢の感覚器官である鋤鼻器官に発現する1種類の受容体V2Rp5により受容される2).しかし,ESP1が作用するための神経基盤は不明であった(図1).1種類の受容体という限定された入力により個体の生得的な行動に影響をおよぼすESP1はすぐれたモデルになると考え,ESP1のシグナルが伝達される神経回路の解明をめざした.さらに,近年の研究により,ESP1はオスのマウスに対してもフェロモンとしてはたらき,ほかのオスのマウスに対する攻撃行動を促進することがわかった3).同じフェロモンが雌雄で異なる行動をひき起こす機序を理解するため,その神経回路を雌雄で比較することも試みた.
ESP1が1種類の受容体V2Rp5により受容されることを利用し,その下流の領域を順行性ウイルストレーサー4) により可視化することを試みた.このウイルスは組換え酵素Creに依存的してニューロンにおける増殖能をもち蛍光タンパク質tdTomatoを発現する.さらに,このウイルスは順行性にニューロンからニューロンへと伝達されるため,はじめに導入されたニューロンを起点として上流のニューロンから下流のニューロンへとtdTomatoによる標識が広がる.この順行性ウイルストレーサーをV2Rp5を発現するニューロンに選択的に感染させるため,V2Rp5遺伝子の発現制御領域の下流にCre遺伝子を挿入したトランスジェニックマウスを作製し,順行性ウイルストレーサーを感染させてtdTomatoによる標識を経時的に観察した.その結果,はじめにV2Rp5を発現する鋤鼻器官のニューロンにおいて,つづいて内側扁桃体,扁桃体内側皮質核,分界条床核といった扁桃体において,そののち,視床下部腹内側核や視索前野といったマウスの生得的な行動に強くかかわる視床下部において,tdTomatoによる標識が観察された.このとき,標識のパターンを雌雄で比較したものの明瞭な違いは観察されなかった.
V2Rp5を発現するニューロンと接続することが明らかにされた内側扁桃体,扁桃体内側皮質核,分界条床核がESP1の効果に必要であるかどうか検証した.神経活動を抑制するタンパク質hM4Diを用いてこれらの領域をそれぞれ薬理遺伝学的に抑制し,ESP1を受容したのちのメスのマウスの行動を観察した.その結果,内側扁桃体を抑制したときのみ,ESP1によるメスのマウスにおける性行動の促進効果がみられなくなった.したがって,ESP1の効果には内側扁桃体が必要であることが明らかにされた.内側扁桃体は匂いやフェロモンのシグナルを多様な行動あるいは情動の変化にかかわる視床下部の領域へと伝達するハブのような役割をはたすと考えられてきた.そこで,内側扁桃体においてESP1のシグナルがどのように表現され下流の領域へと伝達されるか検証した.
初期応答遺伝子であるcFos遺伝子を指標としてESP1による刺激により活性化されたニューロンの分布を解析した.その結果,雌雄どちらにおいても内側扁桃体の後腹側部が顕著に活性化された.内側扁桃体の後腹側部の下流の領域である視床下部腹内側核の背側部あるいは視索前野に投射するニューロンを逆行性のトレーサーにより標識した個体に対しても同様に解析した.その結果,メスのマウスにおいてはESP1による刺激により視索前野ではなく視床下部腹内側核の背側部に投射するニューロンが活性化されたのに対し,オスのマウスにおいては視索前野に投射するニューロンのみが活性化された.さらに,軸索の投射の様式を解析することにより,これら2種類のニューロンは視床下部腹内側核の背側部あるいは視索前野それぞれに対し優先的に軸索を投射する特異性をもつニューロンであることがわかった.以上の結果から,ESP1のシグナルは内側扁桃体の後腹側部の投射に特異性のあるニューロンにより雌雄で異なる下流の領域へと分配されることが明らかにされた.
ESP1の効果における視床下部腹内側核の背側部の必要性について解析した.視床下部腹内側核の背側部にはSF1遺伝子が特異的に発現する.そこで,視床下部腹内側核の背側部のSF1遺伝子を発現するニューロンをジフテリア毒素により選択的に破壊,あるいは,hM4Diにより神経活動を抑制したしたところ,これらのマウスにおいてはESP1によるメスの性行動の促進効果はみられなかった.したがって,ESP1の効果には視床下部腹内側核の背側部が必要であることがわかった.
視床下部腹内側核の背側部の下流の領域である視索前野や中脳中心灰白質への経路の必要性について検証した.それぞれの経路を選択的に抑制し,ESP1を受容したメスのマウスの性行動を解析したところ,視索前野への経路ではなく,中脳中心灰白質への経路を抑制したときのみ,ESP1によるメスのマウスにおける性行動の促進効果がみられなくなった.以上の結果から,ESP1のシグナルは内側扁桃体の後腹側部から視床下部腹内側核の背側部,さらに,中脳中心灰白質へと伝達されることによりメスのマウスの性行動を促進することがわかった.さらに,中脳中心灰白質のニューロンのうち,エストロゲン受容体αを発現するニューロンを破壊あるいはhM4Diにより抑制したところ,ESP1によるメスのマウスにおける性行動の促進効果が弱まった.したがって,ESP1の効果には中脳中心灰白質のエストロゲン受容体αを発現するニューロンが関与することが示唆された.
内側扁桃体の後腹側部から視床下部腹内側核の背側部への神経回路は,古典的には天敵からの忌避行動に関与することが示唆されている5).さらに近年,鋤鼻器官は天敵に由来する匂い物質を受容することも明らかにされつつある6).では,同じように鋤鼻器官により受容され,内側扁桃体の後腹側部から視床下部腹内側核の背側部の神経回路を活性化させるESP1のシグナルと天敵のシグナルは,どのように性行動と忌避行動という異なる行動を促進するのだろうか? これを明らかにするため,ESP1のシグナルおよび天敵のシグナルが脳においてどのように表現されるかを検証した.連続する2種類の刺激により活性化されるニューロンを同一の個体においてそれぞれ標識することのできるcat-FISH法を用いた解析の結果,ESP1あるいはマウスの天敵であるヘビの抜け殻により活性化されたニューロンは,副嗅球,内側扁桃体の後腹側部,視床下部腹内側核の背側部のいずれにおいても分かれて分布した.
この2種類のニューロンの神経活動が行動の変化をひき起こすのに十分であるかどうかを検証するため,TRAP法を用いた7).TRAP法においては,特定の刺激により活性化されるニューロンにおいてのみCreと改変型のエストロゲン受容体との融合タンパク質が発現するマウスを用いる.通常,この融合タンパク質は細胞質に局在するが,アゴニストが存在するときだけ核へと移行し組換え活性をもつことから,アゴニストを投与したのち特定の刺激をあたえることにより,その刺激に応答したニューロンだけをCreに依存して標識することができる.その結果,ESP1による刺激により活性化される視床下部腹内側核の背側部のニューロンを薬理遺伝学的あるいは光遺伝学的に再活性化するとロードシス反射が促進されたのに対し,ヘビの抜け殻による刺激により活性化されるニューロンの再活性化によりロードシス反射は促進されなかった.以上の結果と,ESP1とヘビに由来する因子は末梢において異なる受容体を発現するニューロンにより受容されるという知見8) とをあわせると,末梢の器官から行動を制御する脳の中枢の領域まで,ESP1のシグナルとヘビの抜け殻のシグナルはニューロンのレベルにおいて異なる神経回路により伝達されることが明らかにされた.
この研究においては,フェロモンが個体の行動に影響をおよぼすための神経回路を,末梢から脳の中枢へ,上流から下流へと,1段階ずつ明らかにするという戦略をとった.結果として,ESP1が末梢においてV2Rp5を発現するニューロンに受容されたのち,そのシグナルが副嗅球,内側扁桃体の後腹側部,視床下部腹内側核の背側部,中脳へと伝達されることによりメスのマウスの性行動を促進することがわかった(図2).さらに,ESP1のシグナルと天敵のシグナルはそれぞれに専用の神経回路を使って脳の行動中枢へと伝達されていた(図2).このシンプルな神経構造は,生得的に特定の感覚入力が対応する行動出力へと変換されるためには好適なシステムなのであろう.
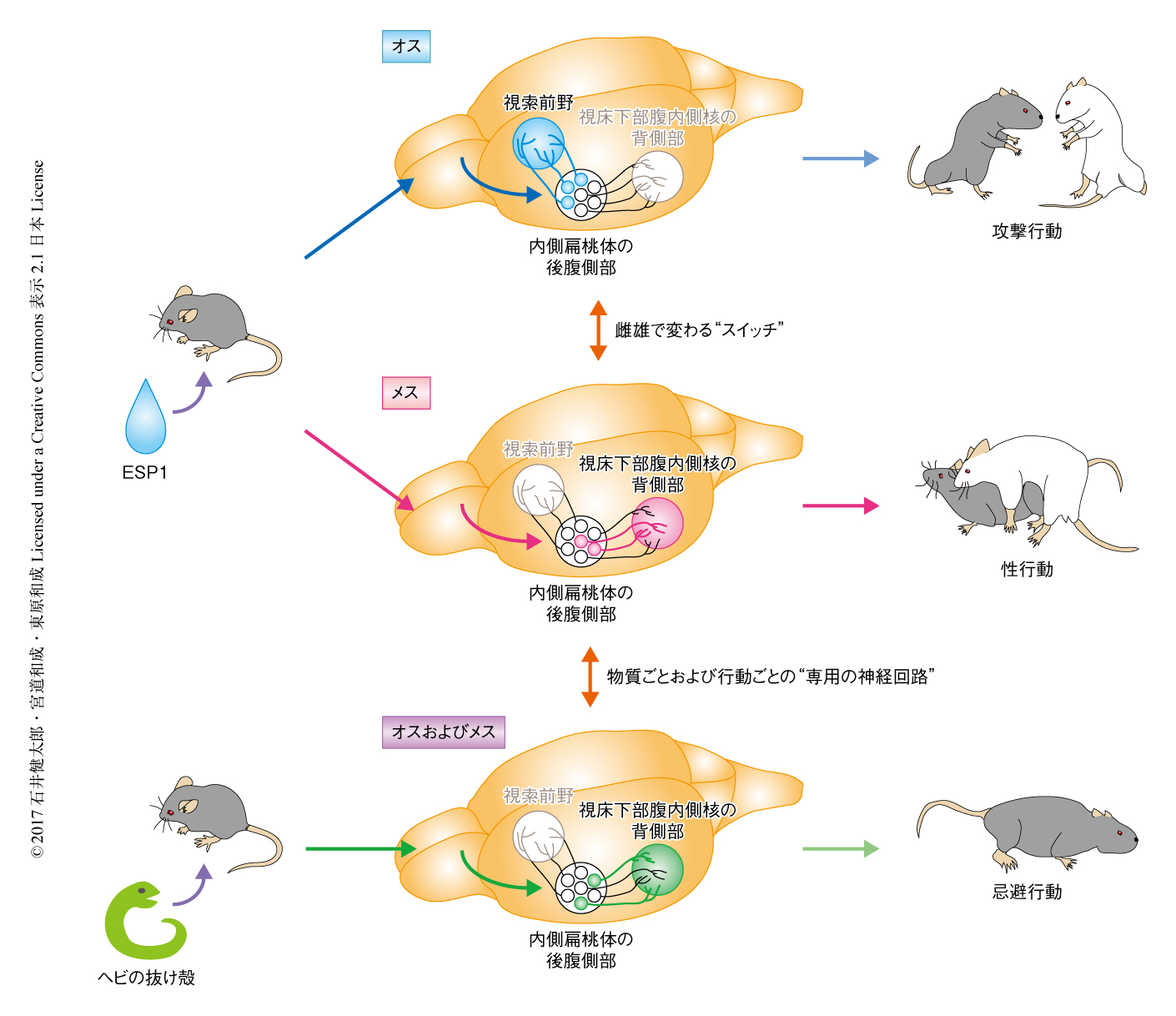
また,同じESP1のシグナルを伝達する神経回路に雌雄の差のあることもわかった(図2).この神経回路の構造の特徴は,ショウジョウバエにおいて性フェロモンcis-バクセン酸アセテートを伝達する神経回路と類似していた.cis-バクセン酸アセテートはオスのショウジョウバエから分泌されるフェロモンで,メスのショウジョウバエに対しては性行動の促進,オスのショウジョウバエに対しては攻撃行動の促進という性的二型な効果をもつ9,10).ショウジョウバエのシグナルは哺乳類の扁桃体にあたる領域と考えられている側角において雌雄で異なるニューロンへと伝達される11).したがって,昆虫から哺乳類まで,扁桃体あるいは側角におけるオスに由来の感覚シグナルの処理には共通する原理があると考察される.ショウジョウバエにおいてはcis-バクセン酸アセテートのシグナル伝達において雌雄の差が生じるための神経基盤の理解が進んでおり,たとえば,fruitless遺伝子はメス型からオス型の回路へのスイッチの役割をもつ11).マウスにおいても,ESP1をモデルとして性的二型な神経回路の構造が生じる機構の解析が進めばおもしろい.
略歴:東京大学大学院農学生命科学研究科博士課程 在学中.
研究テーマ:メスのマウスの性行動を制御する神経基盤.
関心事:ラーメン二郎と糖質制限ダイエット.
宮道 和成(Kazunari Miyamichi)
東京大学大学院農学生命科学研究科 特任准教授.
東原 和成(Kazushige Touhara)
東京大学大学院農学生命科学研究科 教授.
研究室URL:http://park.itc.u-tokyo.ac.jp/biological-chemistry/
© 2017 石井健太郎・宮道和成・東原和成 Licensed under CC 表示 2.1 日本
(東京大学大学院農学生命科学研究科 応用生命化学専攻生物化学研究室)
email:宮道和成,東原和成
DOI: 10.7875/first.author.2017.057
A labeled-line neural circuit for pheromone-mediated sexual behaviors in mice.
Kentaro K. Ishii, Takuya Osakada, Hiromi Mori, Nobuhiko Miyasaka, Yoshihiro Yoshihara, Kazunari Miyamichi, Kazushige Touhara
Neuron, 95, 123-137 (2017)
要 約
生物の行動,とくに本能的な行動は匂いなどの化学物質により制御される.たとえば,マウスにおいては異性,同性,天敵に由来する物質が,それぞれ,性行動,攻撃行動,忌避行動をひき起こすことが知られている.しかし,特定の感覚シグナルが適切な行動をひき起こすための神経基盤については不明であった.今回,筆者らは,単一の受容体を介して生得的にマウスの行動に影響をおよぼす性フェロモンESP1に着目し,ESP1がメスのマウスにおいて性行動の促進効果をひき起こすための神経回路を末梢から行動を制御する脳の中枢までニューロンのレベルにおいて明らかにした.また,ESP1は雌雄で異なる神経回路より処理されることも見い出された.さらに,ESP1のシグナルと天敵のシグナルを伝達する神経回路は同じ領域を介するものの,ニューロンのレベルにおいては異なっていた.これらの一連の結果から,生得的な行動を制御する感覚シグナルそれぞれには専用の神経回路が用意される可能性が示唆された.このようなシンプルな構造は,特定の感覚入力に対し生得的に適切な行動を出力するうえで好適なのかもしれない.
はじめに
生物は外界からさまざまなシグナルを感知し,それにもとづき適切な行動を選択する.多くの場合,この過程には学習が必要であるが,なかには学習を必要とせず特定の本能的な行動をひき起こすシグナルもある.多くの生物において,匂いなどの化学物質のシグナルは本能的な行動の制御に重要な役割をはたすことが知られている.なかでも,同じ種のほかの個体に対し行動や情動の変化をひき起こす物質はフェロモンとよばれ,さまざまな生物において確認されている.たとえば,はじめて化学構造が明らかにされたフェロモンであるボンビコールは,メスのカイコガから発せられ,それだけでオスを誘引する効果をもつ1).哺乳類においてもさまざまなフェロモンが同定されており,たとえば,オスのブタの唾液に分泌され,発情状態のメスに交尾姿勢をひき起こさせる物質としてアンドロステノンが同定されている1).このように,フェロモンが“何か?”その分子実態が明らかにされつつある一方,フェロモンが“どうはたらくのか?”つまり,特定の感覚シグナルが適切な生得的な行動をひき起こすための神経基盤は哺乳類においてはほとんど明らかにされていない.この問いを解く鍵は,フェロモンのシグナルが伝達される神経回路を,末梢の器官から行動を制御する脳の中枢の領域までニューロンのレベルにおいて明らかにすることであると考えた.そのためには,シンプルな感覚入力により行動の劇的な変化をひき起こすような物質が最適である.
筆者らの研究室における先行研究により,オスのマウスの涙液に分泌されるESP1(exocrine gland-secreting peptide 1)とよばれる約7 kDaのペプチドがフェロモンとして機能することが明らかにされた2)(新着論文レビュー でも掲載).ESP1を受容したメスのマウスは,オスのマウスに対し性的な受け入れ行動であるロードシス反射を示す割合が増加する.また,ESP1はマウスの末梢の感覚器官である鋤鼻器官に発現する1種類の受容体V2Rp5により受容される2).しかし,ESP1が作用するための神経基盤は不明であった(図1).1種類の受容体という限定された入力により個体の生得的な行動に影響をおよぼすESP1はすぐれたモデルになると考え,ESP1のシグナルが伝達される神経回路の解明をめざした.さらに,近年の研究により,ESP1はオスのマウスに対してもフェロモンとしてはたらき,ほかのオスのマウスに対する攻撃行動を促進することがわかった3).同じフェロモンが雌雄で異なる行動をひき起こす機序を理解するため,その神経回路を雌雄で比較することも試みた.
1.順行性ウイルストレーサーを用いたESP1のシグナルを伝達する神経回路の可視化
ESP1が1種類の受容体V2Rp5により受容されることを利用し,その下流の領域を順行性ウイルストレーサー4) により可視化することを試みた.このウイルスは組換え酵素Creに依存的してニューロンにおける増殖能をもち蛍光タンパク質tdTomatoを発現する.さらに,このウイルスは順行性にニューロンからニューロンへと伝達されるため,はじめに導入されたニューロンを起点として上流のニューロンから下流のニューロンへとtdTomatoによる標識が広がる.この順行性ウイルストレーサーをV2Rp5を発現するニューロンに選択的に感染させるため,V2Rp5遺伝子の発現制御領域の下流にCre遺伝子を挿入したトランスジェニックマウスを作製し,順行性ウイルストレーサーを感染させてtdTomatoによる標識を経時的に観察した.その結果,はじめにV2Rp5を発現する鋤鼻器官のニューロンにおいて,つづいて内側扁桃体,扁桃体内側皮質核,分界条床核といった扁桃体において,そののち,視床下部腹内側核や視索前野といったマウスの生得的な行動に強くかかわる視床下部において,tdTomatoによる標識が観察された.このとき,標識のパターンを雌雄で比較したものの明瞭な違いは観察されなかった.
2.ESP1のシグナルを雌雄で異なる分配する神経回路の解析
V2Rp5を発現するニューロンと接続することが明らかにされた内側扁桃体,扁桃体内側皮質核,分界条床核がESP1の効果に必要であるかどうか検証した.神経活動を抑制するタンパク質hM4Diを用いてこれらの領域をそれぞれ薬理遺伝学的に抑制し,ESP1を受容したのちのメスのマウスの行動を観察した.その結果,内側扁桃体を抑制したときのみ,ESP1によるメスのマウスにおける性行動の促進効果がみられなくなった.したがって,ESP1の効果には内側扁桃体が必要であることが明らかにされた.内側扁桃体は匂いやフェロモンのシグナルを多様な行動あるいは情動の変化にかかわる視床下部の領域へと伝達するハブのような役割をはたすと考えられてきた.そこで,内側扁桃体においてESP1のシグナルがどのように表現され下流の領域へと伝達されるか検証した.
初期応答遺伝子であるcFos遺伝子を指標としてESP1による刺激により活性化されたニューロンの分布を解析した.その結果,雌雄どちらにおいても内側扁桃体の後腹側部が顕著に活性化された.内側扁桃体の後腹側部の下流の領域である視床下部腹内側核の背側部あるいは視索前野に投射するニューロンを逆行性のトレーサーにより標識した個体に対しても同様に解析した.その結果,メスのマウスにおいてはESP1による刺激により視索前野ではなく視床下部腹内側核の背側部に投射するニューロンが活性化されたのに対し,オスのマウスにおいては視索前野に投射するニューロンのみが活性化された.さらに,軸索の投射の様式を解析することにより,これら2種類のニューロンは視床下部腹内側核の背側部あるいは視索前野それぞれに対し優先的に軸索を投射する特異性をもつニューロンであることがわかった.以上の結果から,ESP1のシグナルは内側扁桃体の後腹側部の投射に特異性のあるニューロンにより雌雄で異なる下流の領域へと分配されることが明らかにされた.
3.ESP1によるメスのマウスにおける性行動の促進効果に必要な神経回路の解析
ESP1の効果における視床下部腹内側核の背側部の必要性について解析した.視床下部腹内側核の背側部にはSF1遺伝子が特異的に発現する.そこで,視床下部腹内側核の背側部のSF1遺伝子を発現するニューロンをジフテリア毒素により選択的に破壊,あるいは,hM4Diにより神経活動を抑制したしたところ,これらのマウスにおいてはESP1によるメスの性行動の促進効果はみられなかった.したがって,ESP1の効果には視床下部腹内側核の背側部が必要であることがわかった.
視床下部腹内側核の背側部の下流の領域である視索前野や中脳中心灰白質への経路の必要性について検証した.それぞれの経路を選択的に抑制し,ESP1を受容したメスのマウスの性行動を解析したところ,視索前野への経路ではなく,中脳中心灰白質への経路を抑制したときのみ,ESP1によるメスのマウスにおける性行動の促進効果がみられなくなった.以上の結果から,ESP1のシグナルは内側扁桃体の後腹側部から視床下部腹内側核の背側部,さらに,中脳中心灰白質へと伝達されることによりメスのマウスの性行動を促進することがわかった.さらに,中脳中心灰白質のニューロンのうち,エストロゲン受容体αを発現するニューロンを破壊あるいはhM4Diにより抑制したところ,ESP1によるメスのマウスにおける性行動の促進効果が弱まった.したがって,ESP1の効果には中脳中心灰白質のエストロゲン受容体αを発現するニューロンが関与することが示唆された.
4.ESP1のシグナルと天敵のシグナルの伝達される神経回路の比較
内側扁桃体の後腹側部から視床下部腹内側核の背側部への神経回路は,古典的には天敵からの忌避行動に関与することが示唆されている5).さらに近年,鋤鼻器官は天敵に由来する匂い物質を受容することも明らかにされつつある6).では,同じように鋤鼻器官により受容され,内側扁桃体の後腹側部から視床下部腹内側核の背側部の神経回路を活性化させるESP1のシグナルと天敵のシグナルは,どのように性行動と忌避行動という異なる行動を促進するのだろうか? これを明らかにするため,ESP1のシグナルおよび天敵のシグナルが脳においてどのように表現されるかを検証した.連続する2種類の刺激により活性化されるニューロンを同一の個体においてそれぞれ標識することのできるcat-FISH法を用いた解析の結果,ESP1あるいはマウスの天敵であるヘビの抜け殻により活性化されたニューロンは,副嗅球,内側扁桃体の後腹側部,視床下部腹内側核の背側部のいずれにおいても分かれて分布した.
この2種類のニューロンの神経活動が行動の変化をひき起こすのに十分であるかどうかを検証するため,TRAP法を用いた7).TRAP法においては,特定の刺激により活性化されるニューロンにおいてのみCreと改変型のエストロゲン受容体との融合タンパク質が発現するマウスを用いる.通常,この融合タンパク質は細胞質に局在するが,アゴニストが存在するときだけ核へと移行し組換え活性をもつことから,アゴニストを投与したのち特定の刺激をあたえることにより,その刺激に応答したニューロンだけをCreに依存して標識することができる.その結果,ESP1による刺激により活性化される視床下部腹内側核の背側部のニューロンを薬理遺伝学的あるいは光遺伝学的に再活性化するとロードシス反射が促進されたのに対し,ヘビの抜け殻による刺激により活性化されるニューロンの再活性化によりロードシス反射は促進されなかった.以上の結果と,ESP1とヘビに由来する因子は末梢において異なる受容体を発現するニューロンにより受容されるという知見8) とをあわせると,末梢の器官から行動を制御する脳の中枢の領域まで,ESP1のシグナルとヘビの抜け殻のシグナルはニューロンのレベルにおいて異なる神経回路により伝達されることが明らかにされた.
おわりに
この研究においては,フェロモンが個体の行動に影響をおよぼすための神経回路を,末梢から脳の中枢へ,上流から下流へと,1段階ずつ明らかにするという戦略をとった.結果として,ESP1が末梢においてV2Rp5を発現するニューロンに受容されたのち,そのシグナルが副嗅球,内側扁桃体の後腹側部,視床下部腹内側核の背側部,中脳へと伝達されることによりメスのマウスの性行動を促進することがわかった(図2).さらに,ESP1のシグナルと天敵のシグナルはそれぞれに専用の神経回路を使って脳の行動中枢へと伝達されていた(図2).このシンプルな神経構造は,生得的に特定の感覚入力が対応する行動出力へと変換されるためには好適なシステムなのであろう.
また,同じESP1のシグナルを伝達する神経回路に雌雄の差のあることもわかった(図2).この神経回路の構造の特徴は,ショウジョウバエにおいて性フェロモンcis-バクセン酸アセテートを伝達する神経回路と類似していた.cis-バクセン酸アセテートはオスのショウジョウバエから分泌されるフェロモンで,メスのショウジョウバエに対しては性行動の促進,オスのショウジョウバエに対しては攻撃行動の促進という性的二型な効果をもつ9,10).ショウジョウバエのシグナルは哺乳類の扁桃体にあたる領域と考えられている側角において雌雄で異なるニューロンへと伝達される11).したがって,昆虫から哺乳類まで,扁桃体あるいは側角におけるオスに由来の感覚シグナルの処理には共通する原理があると考察される.ショウジョウバエにおいてはcis-バクセン酸アセテートのシグナル伝達において雌雄の差が生じるための神経基盤の理解が進んでおり,たとえば,fruitless遺伝子はメス型からオス型の回路へのスイッチの役割をもつ11).マウスにおいても,ESP1をモデルとして性的二型な神経回路の構造が生じる機構の解析が進めばおもしろい.
文 献
- Wyatt, T. D.: Pheromones and Animal Behaviour: Communication by Smell and Taste. Cambridge University Press, Cambridge (2003)
- Haga, S., Hattori, T., Sato, T. et al.: The male mouse pheromone ESP1 enhances female sexual receptive behaviour through a specific vomeronasal receptor. Nature, 466, 118-122 (2010)[PubMed] [新着論文レビュー]
- Hattori, T., Osakada, T., Matsumoto, A. et al.: Self-exposure to the male pheromone ESP1 enhances male aggressiveness in mice. Curr. Biol., 26, 1229-1234 (2016)[PubMed]
- Lo, L. & Anderson, D. J.: A Cre-dependent, anterograde transsynaptic viral tracer for mapping output pathways of genetically marked neurons. Neuron, 72, 938-950 (2011)[PubMed]
- Canteras, N. S.: The medial hypothalamic defensive system: hodological organization and functional implications. Pharmacol. Biochem. Behav., 71, 481-491 (2002)[PubMed]
- Papes, F., Logan, D. W. & Stowers, L.: The vomeronasal organ mediates interspecies defensive behaviors through detection of protein pheromone homologs. Cell, 141, 692-703 (2010)[PubMed]
- Guenthner, C., Miyamichi, K., Yang, H. H. et al.: Permanent genetic access to transiently active neurons via TRAP: targeted recombination in active populations. Neuron, 78, 773-784 (2013)[PubMed]
- Isogai, Y., Si, S., Pont-Lezica, L. et al.: Molecular organization of vomeronasal chemoreception. Nature, 478, 241-245 (2011)[PubMed]
- Kurtovic, A., Widmer, A. & Dickson, B. J.: A single class of olfactory neurons mediates behavioural responses to a Drosophila sex pheromone. Nature, 446, 542-546 (2007)[PubMed]
- Wang, L. & Anderson, D. J.: Identification of an aggression-promoting pheromone and its receptor neurons in Drosophila. Nature, 463, 227-231 (2010)[PubMed]
- Kohl, J., Ostrovsky, A. D., Frechter, S. et al.: A bidirectional circuit switch reroutes pheromone signals in male and female brains. Cell, 155, 1610-1623 (2013)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:東京大学大学院農学生命科学研究科博士課程 在学中.
研究テーマ:メスのマウスの性行動を制御する神経基盤.
関心事:ラーメン二郎と糖質制限ダイエット.
宮道 和成(Kazunari Miyamichi)
東京大学大学院農学生命科学研究科 特任准教授.
東原 和成(Kazushige Touhara)
東京大学大学院農学生命科学研究科 教授.
研究室URL:http://park.itc.u-tokyo.ac.jp/biological-chemistry/
© 2017 石井健太郎・宮道和成・東原和成 Licensed under CC 表示 2.1 日本
