新奇な体験による記憶の保持の強化の機構
竹内 倫徳
(英国Edinburgh大学Centre for Cognitive and Neural Systems)
email:竹内倫徳
DOI: 10.7875/first.author.2016.107
Locus coeruleus and dopaminergic consolidation of everyday memory.
Tomonori Takeuchi, Adrian J. Duszkiewicz, Alex Sonneborn, Patrick A. Spooner, Miwako Yamasaki, Masahiko Watanabe, Caroline C. Smith, Guillén Fernández, Karl Deisseroth, Robert W. Greene, Richard G. M. Morris
Nature, 537, 357-362 (2016)
ヒトや動物において,日常の記憶は記憶の獲得の直前あるいは直後に何か新奇な体験があるとその保持が強化されることが知られている.これまで,この記憶の保持の強化には海馬のドーパミンが重要であり,腹側被蓋野のチロシン水酸化酵素陽性ニューロンがかかわると考えられてきた.筆者らは,マウスにおいて日常の記憶を調べる行動試験を用いて新奇な体験による記憶の保持の強化にかかわるニューロンを探索した.その結果,青斑核のチロシン水酸化酵素陽性ニューロンの発火が新奇な体験に対しとくに感受性の高いこと,青斑核のチロシン水酸化酵素陽性ニューロンは腹側被蓋野のチロシン水酸化酵素陽性ニューロンと比べ海馬により多く投射していること,青斑核のチロシン水酸化酵素陽性ニューロンを光遺伝学的に活性化させると新奇な体験による記憶の保持の強化が模倣されること,新奇な体験による記憶の保持の強化は腹側被蓋野の薬理学的な不活性化に影響されないこと,が明らかにされた.青斑核のチロシン水酸化酵素陽性ニューロンの光遺伝学的な活性化による記憶の保持の強化および海馬CA1領域におけるシナプス伝達の増強は,海馬におけるドーパミンD1受容体の阻害に感受性を示したが,アドレナリン受容体の阻害には影響されなかった.したがって,ノルアドレナリン作動性の青斑核のチロシン水酸化酵素陽性ニューロンは,海馬におけるドーパミンの共放出を介して新奇な体験による記憶の保持の強化を担うことが示唆された.
“晩ごはんにどこで何を食べたか”などのささいな日常の記憶は海馬において無意識に形成され,その多くは1日のあいだに忘れられることが知られている.一方で,“晩ごはんに行く途中に1万円札を拾った”など新奇で思いがけない出来事を直前あるいは直後にともなう場合,ささいな日常の記憶が長期にわたり保持される現象が知られている.その脳における機構を調べるため,近年,新奇の環境の探索をともなうことにより記憶の固定化が促進され,ささいな記憶が長期にわたり保持される動物をモデルとした行動試験が開発された.薬理学的な阻害実験より,この新奇な体験による記憶の保持の強化には海馬におけるドーパミンD1受容体の活性化が必要であることが明らかにされた1).そして,腹側被蓋野のチロシン水酸化酵素陽性ニューロンから海馬への投射がこのドーパミンに依存的な新奇の体験による記憶の保持の強化に関与するとの仮説が提唱された2).一方,海馬の急性切片を用いた電気生理学的な実験により,ノルアドレナリン作動性の青斑核のチロシン水酸化酵素陽性ニューロンが海馬においてドーパミンを放出する可能性が示唆された3).
筆者らは,マウスにおいて日常の記憶を調べる行動試験として日常記憶課題を開発した.そして,腹側被蓋野のチロシン水酸化酵素陽性ニューロンおよび青斑核のチロシン水酸化酵素陽性ニューロンに着目し,日常記憶課題と薬理学的な手法,電気生理学的な手法,解剖学的な手法,光遺伝学的な手法を統合的に適応することにより,脳において新奇性による記憶の保持の強化を担う部位の同定を試みた.
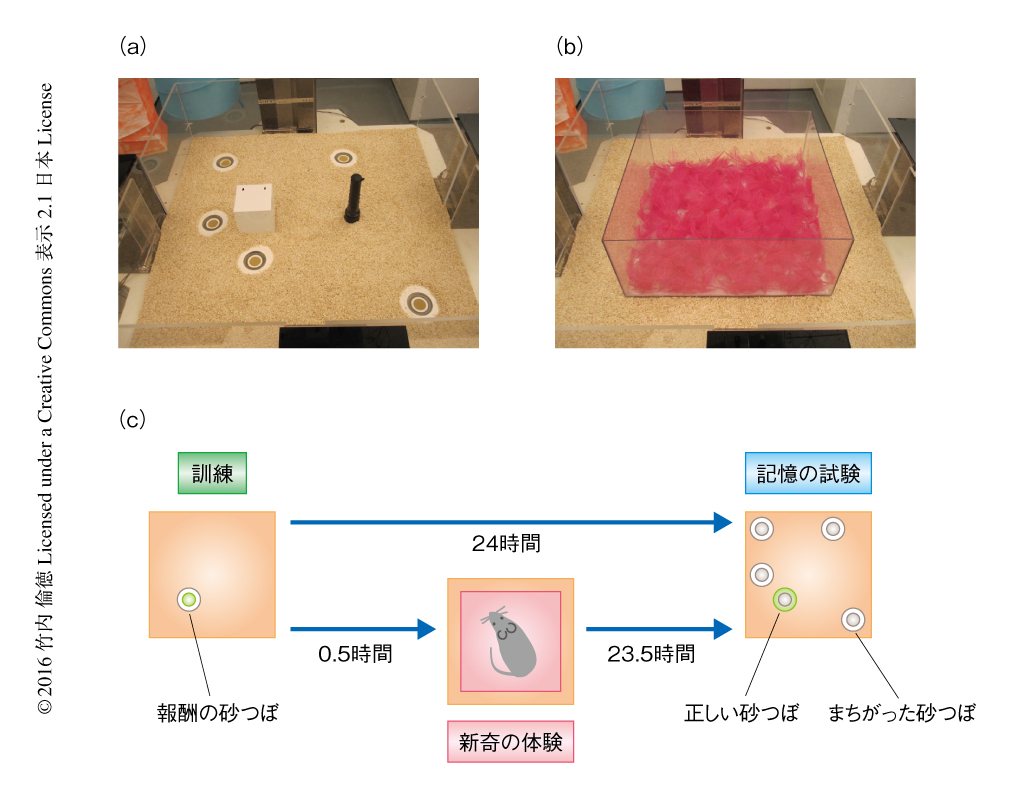
マウスにおいて新奇な体験が日常の記憶を向上させる効果を調べる日常記憶課題を確立した4).この行動試験の大きな特色として,すべてのマウス(被験動物)がすべての実験条件を経験する被験動物内デザインの適応が可能で,動物実験において問題になる個体間のバラつきを最小限にすることが可能になった.日常記憶課題においては,イベントアリーナ装置とよばれるオープンフィールドにおいて,マウスに報酬の餌が底に隠されている砂つぼの場所を記憶させる(図1a).報酬を含む砂つぼの場所は毎日変わるため,マウスはその日その日の特定の砂つぼの場所を記憶する必要がある.報酬量の少ない“弱い訓練”を行ったマウスは,24時間のちには報酬の砂つぼの場所を忘れていた.一方,弱い訓練の30分のちに,新奇な素材を床に敷きつめた新奇体験ボックスを5分間にわたり探索させると(図1b),報酬の砂つぼの場所の記憶は24時間のちにも保持されていた(図1c).背側の海馬に受容体の阻害薬を投与する薬理学的な阻害実験において,この新奇性による記憶の保持の強化は海馬におけるドーパミンD1受容体の阻害に対し感受性を示したが,βアドレナリン受容体の阻害には影響されなかった.これらの結果により,マウスの新奇な体験による記憶の保持の強化には海馬のドーパミンD1受容体の活性化が重要であるという,ラットを用いた先行研究と同様の結果が得られた1).
腹側被蓋野のチロシン水酸化酵素陽性ニューロンあるいは青斑核のチロシン水酸化酵素の陽性ニューロンにおける神経活動が新奇な体験により活性化するかどうか調べた.チロシン水酸化酵素陽性ニューロンを光遺伝学的に同定するため,チロシン水酸化酵素陽性ニューロンにおいてDNA組換え酵素Creを特異的に発現するトランスジェニックマウスの腹側被蓋野あるいは青斑核に,Creに依存的なウイルスベクターを感染させチロシン水酸化酵素陽性ニューロンにおいて光感受性イオンチャネルであるチャネルロドプシンを特異的に発現させた.脳に埋め込んだ光ファイバーを介してチャネルロドプシン陽性ニューロンに光を照射すると,ニューロンの外部から内部へと陽イオンが流入しニューロンは活性化される.多点電極の同時記録によりチャネルロドプシン陽性ニューロンすなわちチロシン水酸化酵素陽性ニューロンが同定される.
同定された腹側被蓋野のチロシン水酸化酵素陽性ニューロンあるいは青斑核のチロシン水酸化酵素陽性ニューロンが,なじみのある環境あるいは新奇の環境をそれぞれ5分間にわたり体験しているあいだどのような発火パターンを示すかを調べた.その結果,新奇な環境を体験しているあいだ腹側被蓋野のチロシン水酸化酵素陽性ニューロンおよび青斑核のチロシン水酸化酵素陽性ニューロンの神経活動は上昇した.ホームケージにおいて記録した神経活動を用いて標準化し比較したところ,新奇な体験による神経活動の上昇の度合いは青斑核のチロシン水酸化酵素陽性ニューロンのほうが腹側被蓋野のチロシン水酸化酵素陽性ニューロンよりも大きかった.新奇な体験に対する青斑核のチロシン水酸化酵素陽性ニューロンの応答は時間の経過とともに減少する馴化がみられた.以上のことから,青斑核のチロシン水酸化酵素陽性ニューロンの発火は新奇な体験に対しとくに感受性の高いことがわかった.
腹側被蓋野のチロシン水酸化酵素陽性ニューロンあるいは青斑核のチロシン水酸化酵素陽性ニューロンから背側の海馬への投射について,特定のニューロンに限定した順行性標識法を用いて検討した.チロシン水酸化酵素陽性ニューロンの軸索を特異的に標識するため,チロシン水酸化酵素陽性ニューロンにおいて蛍光タンパク質eYFPを特異的に発現させた.さらに,海馬に投射する腹側被蓋野のチロシン水酸化酵素陽性ニューロンあるいは青斑核のチロシン水酸化酵素陽性ニューロンに由来する軸索を抗GFP抗体および抗チロシン水酸化酵素抗体による蛍光二重免疫染色法により検出した.GFP-チロシン水酸化酵素二重陽性ニューロンの軸索を定量し,海馬の全体に入力するチロシン水酸化酵素陽性ニューロンの軸索との割合を算出した.その結果,海馬の全体に入力するチロシン水酸化酵素陽性ニューロンの軸索の約97%は青斑核のチロシン水酸化酵素陽性ニューロンに由来し,残りの数%が腹側被蓋野のチロシン水酸化酵素陽性ニューロンに由来した.
青斑核のチロシン水酸化酵素陽性ニューロンが腹側被蓋野のチロシン水酸化酵素陽性ニューロンと比べより多く海馬に投射することは,逆行性の蛍光トレーサーを海馬に投与する逆行性標識法においても確認された.
青斑核のチロシン水酸化酵素陽性ニューロンが新奇な体験による記憶の保持の強化にかかわるのかどうかを検証するため,青斑核のチロシン水酸化酵素陽性ニューロンを光遺伝学的に活性化させることにより新奇な体験による記憶の増強の効果が模倣されるかどうか調べた.チロシン水酸化酵素陽性ニューロンにおいてチャネルロドプシンを特異的に発現させ,青斑核および腹側被蓋野に光ファイバー,背側の海馬に薬剤の注入のためのカニューレを埋め込んだ.日常記憶課題を用いて弱い訓練を行い,その30分のちに,青斑核のチロシン水酸化酵素陽性ニューロンを5分間にわたり光遺伝学的に活性化した.その結果,ふだんは数時間で忘れてしまう報酬を含む砂つぼの場所の記憶が24時間のちにも保持されていた.弱い訓練ののち青斑核のチロシン水酸化酵素陽性ニューロンを活性化しない場合には24時間のちには報酬を含む砂つぼの場所は忘れられていた.一方,チャネルロドプシンを発現させた同じマウスを用いて,弱い訓練ののち腹側被蓋野のチロシン水酸化酵素陽性ニューロンを光遺伝学的に活性化した際には,新奇な体験による記憶の保持の強化は模倣されなかった.
ここで,ノルアドレナリン作動性の青斑核のチロシン水酸化酵素陽性ニューロンの光遺伝学的な活性化により記憶の保持が強化されたが,その一方で,新奇な体験による記憶の増強の効果においては海馬におけるドーパミンD1受容体の活性化が重要である,というパラドックスに直面した.この問題を解決するため,日常記憶課題において弱い訓練ののち背側の海馬に受容体の阻害薬を投与し,そののち,青斑核のチロシン水酸化酵素陽性ニューロンを光遺伝学的に活性化して記憶の保持の強化に対する影響について調べた.その結果,青斑核のチロシン水酸化酵素陽性ニューロンの活性化による記憶の保持の強化は,海馬におけるドーパミンD1受容体の阻害に対し感受性を示したが,βアドレナリン受容体の阻害による影響はなかった.すなわち,青斑核のチロシン水酸化酵素陽性ニューロンから海馬へのドーパミンの出力が記憶の保持の強化に関与することが強く示唆された.
青斑核のチロシン水酸化酵素陽性ニューロンの光遺伝学的な活性化による記憶の保持の強化の機構について調べるため,海馬の急性切片を用いて電気生理学的な実験を行った.海馬CA1野の錐体細胞の細胞体からパッチクランプ記録を行い,青斑核のチロシン水酸化酵素陽性ニューロンの軸索の光遺伝学的な活性化によるCA3-CA1シナプスの興奮性シナプス後電流への影響について調べた.海馬に投射する青斑核のチロシン水酸化酵素陽性ニューロンの軸索を活性化することにより,CA3-CA1シナプスの興奮性シナプス後電流は徐々に増大した.この増大はαアドレナリン受容体あるいはβアドレナリン受容体の阻害による影響はまったくみられなかった.一方,ドーパミンD1受容体の阻害剤の存在下においては,青斑核に由来する軸索の活性化によるCA3-CA1シナプスの興奮性シナプス後電流の増大は消失した.
青斑核のチロシン水酸化酵素陽性ニューロンの軸索の光遺伝学的な活性化によるCA3-CA1シナプスの長期増強に対する影響について調べた.長期増強の指標として,海馬CA1野の錐体細胞のフィールド興奮性シナプス後電位の傾斜を測定した.Schaffer側枝へのθバースト刺激により長期増強が誘導されたが,この長期増強は海馬に投射する青斑核のチロシン水酸化酵素陽性ニューロンの軸索を活性化することにより増大した.しかし,青斑核に由来する軸索の活性化による長期増強の増大は,海馬におけるドーパミンD1受容体の阻害により消失した.以上の結果より,青斑核のチロシン水酸化酵素陽性ニューロンの軸索の活性化を介したCA3-CA1シナプス伝達の増強は,青斑核のチロシン水酸化酵素陽性ニューロンの軸索からのドーパミンの共放出を介した機構である可能性が強く示唆された.
新奇な体験の最中に青斑核のチロシン水酸化酵素陽性ニューロンを不活性化することにより新奇な体験による記憶の保持の強化が消失するかどうか検討した.光遺伝学的な不活性化は適さないと判断されたことから,薬理学的な手法を用いて青斑核のニューロンを不活性化した.日常記憶課題を用いて弱い訓練を行い,新奇な体験の20分まえにα2アドレナリン受容体の作用薬であるクロニジンを腹腔内投与した.クロニジンの腹腔内投与は,青斑核のニューロンの神経活動を抑制するが5),腹側被蓋野のニューロンの神経活動には影響しないこと6) が報告されている.その結果,新奇な体験による記憶の増強の効果が消失した.
新奇な体験の最中に腹側被蓋野のニューロンを不活性化することにより,新奇な体験による記憶の増強の効果はどのように影響されるか検討した.日常記憶課題を用いて弱い訓練を行い,新奇な体験の直前に局所の麻酔薬であるリドカインを腹側被蓋野に注入して不活性化した.予想どおり,新奇な体験の最中の腹側被蓋野のニューロンの不活性化は,新奇な体験による記憶の保持の強化に影響しなかった.以上の結果から,新奇な体験による記憶の保持の強化には,新奇な体験の際の青斑核のニューロンの神経活動が必要であることが強く示唆された.
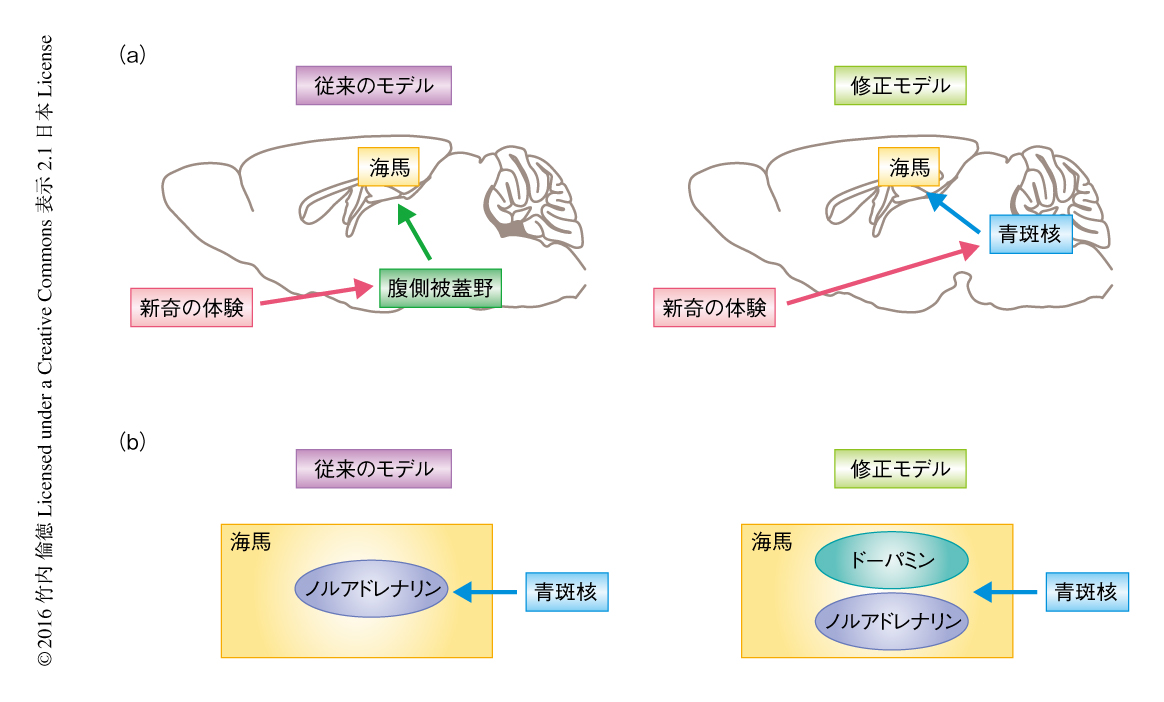
今回,筆者らは,脳において新奇な体験による記憶の保持の強化を担う部位の同定を試み,青斑核のチロシン水酸化酵素陽性ニューロンから海馬へのドーパミンの出力が新奇な体験による記憶の保持の強化に関与することを強く示唆する結果が得られた.その主要な意義のひとつは,青斑核のチロシン水酸化酵素陽性ニューロンから海馬への入力が新奇な体験の情報を伝達しているという点である.このことから,海馬-腹側被蓋野ループモデル2) という有力な仮説とは異なる,新奇な体験の情報を伝達する経路の存在が示唆される(図2a).もうひとつは,青斑核のチロシン水酸化酵素陽性ニューロンの出力としてドーパミンが重要であるという点である.このことから,青斑核のチロシン水酸化酵素陽性ニューロンがノルアドレナリンを放出して神経活動を修飾するという教科書的な基本概念の修正が提案される(図2b).しかしながら,青斑核のチロシン水酸化酵素陽性ニューロンがドーパミンの出力を介して海馬に新奇な体験の情報を伝達し記憶の保持の強化に関与することについての検証はまだ十分ではなく,この仮説が広く認知されるためにはさらなる実験が必要である.
日常の記憶の保持の強化は新奇な体験のみならず,報酬量の増加によってもひき起こされる.新奇な体験と報酬の量や予測の変化はまったく違う情報であり,異なる神経修飾の経路が使われている可能性が考えられる.これまでの研究により,黒質緻密部や腹側被蓋野のチロシン水酸化酵素陽性ニューロンは報酬にもとづく学習や動機づけなどに重要な役割をはたすと考えられている7).最近,海馬に投射する腹側被蓋野のチロシン水酸化酵素陽性ニューロンの軸索の光遺伝学的な活性化によりCA3-CA1シナプスの伝達が双方向性に修飾されることが報告された8).しかしながら,筆者らの日常記憶課題を用いた実験においては,弱い訓練の30分のちにおける腹側被蓋野のチロシン水酸化酵素陽性ニューロンの光遺伝学的な活性化は24時間のちの記憶の保持の強化を示さなかった.一方で,別の研究グループにより,空間学習の獲得の最中に海馬に投射する腹側被蓋野のチロシン水酸化酵素陽性ニューロンの軸索を光遺伝学的に活性化することにより,1時間のちの記憶の保持が強化されることが示されている9).これらの結果から,記憶の獲得と腹側被蓋野のチロシン水酸化酵素陽性ニューロンの活性化のタイミングが,そのほかの強化学習と同様に,腹側被蓋野を介したドーパミンに依存的な記憶の保持の強化の重要な要素である可能性が示唆された.
新奇な体験とは,ある行動の最中に報酬とは無関係な予期しない体験をすることである.今回の結果から,新奇な体験は青斑核のチロシン水酸化酵素陽性ニューロンを活性化し,海馬へのドーパミンの出力を介して可塑性に関連するタンパク質の発現を誘導して,記憶の固定化を促進することにより記憶の保持を強化すると考えられる.多くの領域からの入力をうけ,非常に広範な領域に分散的に投射するノルアドレナリン作動性の青斑核のチロシン水酸化酵素陽性ニューロンは10),新奇性,覚醒と睡眠,注意,ストレスなどに関与すると考えられてきた.青斑核のチロシン水酸化酵素陽性ニューロンが記憶の保持の強化を発揮する時間枠は,腹側被蓋野のチロシン水酸化酵素陽性ニューロンと比較して精度が低く,記憶の固定化の神経機構を説明する“シナプスタグ仮説”11,12) に非常によくあてはまる.今回の筆者らの研究により,われわれの日常の記憶が直前あるいは直後の新奇な体験により修飾され,その保持が強化される神経機構の一端が明らかにされた.今後,この分子基盤を明らかにすることをつうじて,日常の記憶に障害がみられる健忘症を予防または改善する新たな創薬への貢献が期待される.
略歴:2000年 東京大学大学院医学系研究科博士後期課程 修了,同年 同 助手を経て,2008年より英国Edinburgh大学 博士後研究員.
研究テーマ:新奇性および知識に依存した迅速な学習の神経機構および分子機構.
© 2016 竹内 倫徳 Licensed under CC 表示 2.1 日本
(英国Edinburgh大学Centre for Cognitive and Neural Systems)
email:竹内倫徳
DOI: 10.7875/first.author.2016.107
Locus coeruleus and dopaminergic consolidation of everyday memory.
Tomonori Takeuchi, Adrian J. Duszkiewicz, Alex Sonneborn, Patrick A. Spooner, Miwako Yamasaki, Masahiko Watanabe, Caroline C. Smith, Guillén Fernández, Karl Deisseroth, Robert W. Greene, Richard G. M. Morris
Nature, 537, 357-362 (2016)
要 約
ヒトや動物において,日常の記憶は記憶の獲得の直前あるいは直後に何か新奇な体験があるとその保持が強化されることが知られている.これまで,この記憶の保持の強化には海馬のドーパミンが重要であり,腹側被蓋野のチロシン水酸化酵素陽性ニューロンがかかわると考えられてきた.筆者らは,マウスにおいて日常の記憶を調べる行動試験を用いて新奇な体験による記憶の保持の強化にかかわるニューロンを探索した.その結果,青斑核のチロシン水酸化酵素陽性ニューロンの発火が新奇な体験に対しとくに感受性の高いこと,青斑核のチロシン水酸化酵素陽性ニューロンは腹側被蓋野のチロシン水酸化酵素陽性ニューロンと比べ海馬により多く投射していること,青斑核のチロシン水酸化酵素陽性ニューロンを光遺伝学的に活性化させると新奇な体験による記憶の保持の強化が模倣されること,新奇な体験による記憶の保持の強化は腹側被蓋野の薬理学的な不活性化に影響されないこと,が明らかにされた.青斑核のチロシン水酸化酵素陽性ニューロンの光遺伝学的な活性化による記憶の保持の強化および海馬CA1領域におけるシナプス伝達の増強は,海馬におけるドーパミンD1受容体の阻害に感受性を示したが,アドレナリン受容体の阻害には影響されなかった.したがって,ノルアドレナリン作動性の青斑核のチロシン水酸化酵素陽性ニューロンは,海馬におけるドーパミンの共放出を介して新奇な体験による記憶の保持の強化を担うことが示唆された.
はじめに
“晩ごはんにどこで何を食べたか”などのささいな日常の記憶は海馬において無意識に形成され,その多くは1日のあいだに忘れられることが知られている.一方で,“晩ごはんに行く途中に1万円札を拾った”など新奇で思いがけない出来事を直前あるいは直後にともなう場合,ささいな日常の記憶が長期にわたり保持される現象が知られている.その脳における機構を調べるため,近年,新奇の環境の探索をともなうことにより記憶の固定化が促進され,ささいな記憶が長期にわたり保持される動物をモデルとした行動試験が開発された.薬理学的な阻害実験より,この新奇な体験による記憶の保持の強化には海馬におけるドーパミンD1受容体の活性化が必要であることが明らかにされた1).そして,腹側被蓋野のチロシン水酸化酵素陽性ニューロンから海馬への投射がこのドーパミンに依存的な新奇の体験による記憶の保持の強化に関与するとの仮説が提唱された2).一方,海馬の急性切片を用いた電気生理学的な実験により,ノルアドレナリン作動性の青斑核のチロシン水酸化酵素陽性ニューロンが海馬においてドーパミンを放出する可能性が示唆された3).
筆者らは,マウスにおいて日常の記憶を調べる行動試験として日常記憶課題を開発した.そして,腹側被蓋野のチロシン水酸化酵素陽性ニューロンおよび青斑核のチロシン水酸化酵素陽性ニューロンに着目し,日常記憶課題と薬理学的な手法,電気生理学的な手法,解剖学的な手法,光遺伝学的な手法を統合的に適応することにより,脳において新奇性による記憶の保持の強化を担う部位の同定を試みた.
1.新奇な体験による記憶の保持の強化
マウスにおいて新奇な体験が日常の記憶を向上させる効果を調べる日常記憶課題を確立した4).この行動試験の大きな特色として,すべてのマウス(被験動物)がすべての実験条件を経験する被験動物内デザインの適応が可能で,動物実験において問題になる個体間のバラつきを最小限にすることが可能になった.日常記憶課題においては,イベントアリーナ装置とよばれるオープンフィールドにおいて,マウスに報酬の餌が底に隠されている砂つぼの場所を記憶させる(図1a).報酬を含む砂つぼの場所は毎日変わるため,マウスはその日その日の特定の砂つぼの場所を記憶する必要がある.報酬量の少ない“弱い訓練”を行ったマウスは,24時間のちには報酬の砂つぼの場所を忘れていた.一方,弱い訓練の30分のちに,新奇な素材を床に敷きつめた新奇体験ボックスを5分間にわたり探索させると(図1b),報酬の砂つぼの場所の記憶は24時間のちにも保持されていた(図1c).背側の海馬に受容体の阻害薬を投与する薬理学的な阻害実験において,この新奇性による記憶の保持の強化は海馬におけるドーパミンD1受容体の阻害に対し感受性を示したが,βアドレナリン受容体の阻害には影響されなかった.これらの結果により,マウスの新奇な体験による記憶の保持の強化には海馬のドーパミンD1受容体の活性化が重要であるという,ラットを用いた先行研究と同様の結果が得られた1).
2.新奇な体験による青斑核における神経活動の上昇
腹側被蓋野のチロシン水酸化酵素陽性ニューロンあるいは青斑核のチロシン水酸化酵素の陽性ニューロンにおける神経活動が新奇な体験により活性化するかどうか調べた.チロシン水酸化酵素陽性ニューロンを光遺伝学的に同定するため,チロシン水酸化酵素陽性ニューロンにおいてDNA組換え酵素Creを特異的に発現するトランスジェニックマウスの腹側被蓋野あるいは青斑核に,Creに依存的なウイルスベクターを感染させチロシン水酸化酵素陽性ニューロンにおいて光感受性イオンチャネルであるチャネルロドプシンを特異的に発現させた.脳に埋め込んだ光ファイバーを介してチャネルロドプシン陽性ニューロンに光を照射すると,ニューロンの外部から内部へと陽イオンが流入しニューロンは活性化される.多点電極の同時記録によりチャネルロドプシン陽性ニューロンすなわちチロシン水酸化酵素陽性ニューロンが同定される.
同定された腹側被蓋野のチロシン水酸化酵素陽性ニューロンあるいは青斑核のチロシン水酸化酵素陽性ニューロンが,なじみのある環境あるいは新奇の環境をそれぞれ5分間にわたり体験しているあいだどのような発火パターンを示すかを調べた.その結果,新奇な環境を体験しているあいだ腹側被蓋野のチロシン水酸化酵素陽性ニューロンおよび青斑核のチロシン水酸化酵素陽性ニューロンの神経活動は上昇した.ホームケージにおいて記録した神経活動を用いて標準化し比較したところ,新奇な体験による神経活動の上昇の度合いは青斑核のチロシン水酸化酵素陽性ニューロンのほうが腹側被蓋野のチロシン水酸化酵素陽性ニューロンよりも大きかった.新奇な体験に対する青斑核のチロシン水酸化酵素陽性ニューロンの応答は時間の経過とともに減少する馴化がみられた.以上のことから,青斑核のチロシン水酸化酵素陽性ニューロンの発火は新奇な体験に対しとくに感受性の高いことがわかった.
3.青斑核からの海馬への広範な投射
腹側被蓋野のチロシン水酸化酵素陽性ニューロンあるいは青斑核のチロシン水酸化酵素陽性ニューロンから背側の海馬への投射について,特定のニューロンに限定した順行性標識法を用いて検討した.チロシン水酸化酵素陽性ニューロンの軸索を特異的に標識するため,チロシン水酸化酵素陽性ニューロンにおいて蛍光タンパク質eYFPを特異的に発現させた.さらに,海馬に投射する腹側被蓋野のチロシン水酸化酵素陽性ニューロンあるいは青斑核のチロシン水酸化酵素陽性ニューロンに由来する軸索を抗GFP抗体および抗チロシン水酸化酵素抗体による蛍光二重免疫染色法により検出した.GFP-チロシン水酸化酵素二重陽性ニューロンの軸索を定量し,海馬の全体に入力するチロシン水酸化酵素陽性ニューロンの軸索との割合を算出した.その結果,海馬の全体に入力するチロシン水酸化酵素陽性ニューロンの軸索の約97%は青斑核のチロシン水酸化酵素陽性ニューロンに由来し,残りの数%が腹側被蓋野のチロシン水酸化酵素陽性ニューロンに由来した.
青斑核のチロシン水酸化酵素陽性ニューロンが腹側被蓋野のチロシン水酸化酵素陽性ニューロンと比べより多く海馬に投射することは,逆行性の蛍光トレーサーを海馬に投与する逆行性標識法においても確認された.
4.青斑核の光遺伝学的な活性化による記憶の保持の強化
青斑核のチロシン水酸化酵素陽性ニューロンが新奇な体験による記憶の保持の強化にかかわるのかどうかを検証するため,青斑核のチロシン水酸化酵素陽性ニューロンを光遺伝学的に活性化させることにより新奇な体験による記憶の増強の効果が模倣されるかどうか調べた.チロシン水酸化酵素陽性ニューロンにおいてチャネルロドプシンを特異的に発現させ,青斑核および腹側被蓋野に光ファイバー,背側の海馬に薬剤の注入のためのカニューレを埋め込んだ.日常記憶課題を用いて弱い訓練を行い,その30分のちに,青斑核のチロシン水酸化酵素陽性ニューロンを5分間にわたり光遺伝学的に活性化した.その結果,ふだんは数時間で忘れてしまう報酬を含む砂つぼの場所の記憶が24時間のちにも保持されていた.弱い訓練ののち青斑核のチロシン水酸化酵素陽性ニューロンを活性化しない場合には24時間のちには報酬を含む砂つぼの場所は忘れられていた.一方,チャネルロドプシンを発現させた同じマウスを用いて,弱い訓練ののち腹側被蓋野のチロシン水酸化酵素陽性ニューロンを光遺伝学的に活性化した際には,新奇な体験による記憶の保持の強化は模倣されなかった.
ここで,ノルアドレナリン作動性の青斑核のチロシン水酸化酵素陽性ニューロンの光遺伝学的な活性化により記憶の保持が強化されたが,その一方で,新奇な体験による記憶の増強の効果においては海馬におけるドーパミンD1受容体の活性化が重要である,というパラドックスに直面した.この問題を解決するため,日常記憶課題において弱い訓練ののち背側の海馬に受容体の阻害薬を投与し,そののち,青斑核のチロシン水酸化酵素陽性ニューロンを光遺伝学的に活性化して記憶の保持の強化に対する影響について調べた.その結果,青斑核のチロシン水酸化酵素陽性ニューロンの活性化による記憶の保持の強化は,海馬におけるドーパミンD1受容体の阻害に対し感受性を示したが,βアドレナリン受容体の阻害による影響はなかった.すなわち,青斑核のチロシン水酸化酵素陽性ニューロンから海馬へのドーパミンの出力が記憶の保持の強化に関与することが強く示唆された.
5.青斑核の光遺伝学的な活性化によるシナプス伝達の増強
青斑核のチロシン水酸化酵素陽性ニューロンの光遺伝学的な活性化による記憶の保持の強化の機構について調べるため,海馬の急性切片を用いて電気生理学的な実験を行った.海馬CA1野の錐体細胞の細胞体からパッチクランプ記録を行い,青斑核のチロシン水酸化酵素陽性ニューロンの軸索の光遺伝学的な活性化によるCA3-CA1シナプスの興奮性シナプス後電流への影響について調べた.海馬に投射する青斑核のチロシン水酸化酵素陽性ニューロンの軸索を活性化することにより,CA3-CA1シナプスの興奮性シナプス後電流は徐々に増大した.この増大はαアドレナリン受容体あるいはβアドレナリン受容体の阻害による影響はまったくみられなかった.一方,ドーパミンD1受容体の阻害剤の存在下においては,青斑核に由来する軸索の活性化によるCA3-CA1シナプスの興奮性シナプス後電流の増大は消失した.
青斑核のチロシン水酸化酵素陽性ニューロンの軸索の光遺伝学的な活性化によるCA3-CA1シナプスの長期増強に対する影響について調べた.長期増強の指標として,海馬CA1野の錐体細胞のフィールド興奮性シナプス後電位の傾斜を測定した.Schaffer側枝へのθバースト刺激により長期増強が誘導されたが,この長期増強は海馬に投射する青斑核のチロシン水酸化酵素陽性ニューロンの軸索を活性化することにより増大した.しかし,青斑核に由来する軸索の活性化による長期増強の増大は,海馬におけるドーパミンD1受容体の阻害により消失した.以上の結果より,青斑核のチロシン水酸化酵素陽性ニューロンの軸索の活性化を介したCA3-CA1シナプス伝達の増強は,青斑核のチロシン水酸化酵素陽性ニューロンの軸索からのドーパミンの共放出を介した機構である可能性が強く示唆された.
6.新奇な体験による記憶の増強の効果は腹側被蓋野の不活性化に影響されない
新奇な体験の最中に青斑核のチロシン水酸化酵素陽性ニューロンを不活性化することにより新奇な体験による記憶の保持の強化が消失するかどうか検討した.光遺伝学的な不活性化は適さないと判断されたことから,薬理学的な手法を用いて青斑核のニューロンを不活性化した.日常記憶課題を用いて弱い訓練を行い,新奇な体験の20分まえにα2アドレナリン受容体の作用薬であるクロニジンを腹腔内投与した.クロニジンの腹腔内投与は,青斑核のニューロンの神経活動を抑制するが5),腹側被蓋野のニューロンの神経活動には影響しないこと6) が報告されている.その結果,新奇な体験による記憶の増強の効果が消失した.
新奇な体験の最中に腹側被蓋野のニューロンを不活性化することにより,新奇な体験による記憶の増強の効果はどのように影響されるか検討した.日常記憶課題を用いて弱い訓練を行い,新奇な体験の直前に局所の麻酔薬であるリドカインを腹側被蓋野に注入して不活性化した.予想どおり,新奇な体験の最中の腹側被蓋野のニューロンの不活性化は,新奇な体験による記憶の保持の強化に影響しなかった.以上の結果から,新奇な体験による記憶の保持の強化には,新奇な体験の際の青斑核のニューロンの神経活動が必要であることが強く示唆された.
おわりに
今回,筆者らは,脳において新奇な体験による記憶の保持の強化を担う部位の同定を試み,青斑核のチロシン水酸化酵素陽性ニューロンから海馬へのドーパミンの出力が新奇な体験による記憶の保持の強化に関与することを強く示唆する結果が得られた.その主要な意義のひとつは,青斑核のチロシン水酸化酵素陽性ニューロンから海馬への入力が新奇な体験の情報を伝達しているという点である.このことから,海馬-腹側被蓋野ループモデル2) という有力な仮説とは異なる,新奇な体験の情報を伝達する経路の存在が示唆される(図2a).もうひとつは,青斑核のチロシン水酸化酵素陽性ニューロンの出力としてドーパミンが重要であるという点である.このことから,青斑核のチロシン水酸化酵素陽性ニューロンがノルアドレナリンを放出して神経活動を修飾するという教科書的な基本概念の修正が提案される(図2b).しかしながら,青斑核のチロシン水酸化酵素陽性ニューロンがドーパミンの出力を介して海馬に新奇な体験の情報を伝達し記憶の保持の強化に関与することについての検証はまだ十分ではなく,この仮説が広く認知されるためにはさらなる実験が必要である.
日常の記憶の保持の強化は新奇な体験のみならず,報酬量の増加によってもひき起こされる.新奇な体験と報酬の量や予測の変化はまったく違う情報であり,異なる神経修飾の経路が使われている可能性が考えられる.これまでの研究により,黒質緻密部や腹側被蓋野のチロシン水酸化酵素陽性ニューロンは報酬にもとづく学習や動機づけなどに重要な役割をはたすと考えられている7).最近,海馬に投射する腹側被蓋野のチロシン水酸化酵素陽性ニューロンの軸索の光遺伝学的な活性化によりCA3-CA1シナプスの伝達が双方向性に修飾されることが報告された8).しかしながら,筆者らの日常記憶課題を用いた実験においては,弱い訓練の30分のちにおける腹側被蓋野のチロシン水酸化酵素陽性ニューロンの光遺伝学的な活性化は24時間のちの記憶の保持の強化を示さなかった.一方で,別の研究グループにより,空間学習の獲得の最中に海馬に投射する腹側被蓋野のチロシン水酸化酵素陽性ニューロンの軸索を光遺伝学的に活性化することにより,1時間のちの記憶の保持が強化されることが示されている9).これらの結果から,記憶の獲得と腹側被蓋野のチロシン水酸化酵素陽性ニューロンの活性化のタイミングが,そのほかの強化学習と同様に,腹側被蓋野を介したドーパミンに依存的な記憶の保持の強化の重要な要素である可能性が示唆された.
新奇な体験とは,ある行動の最中に報酬とは無関係な予期しない体験をすることである.今回の結果から,新奇な体験は青斑核のチロシン水酸化酵素陽性ニューロンを活性化し,海馬へのドーパミンの出力を介して可塑性に関連するタンパク質の発現を誘導して,記憶の固定化を促進することにより記憶の保持を強化すると考えられる.多くの領域からの入力をうけ,非常に広範な領域に分散的に投射するノルアドレナリン作動性の青斑核のチロシン水酸化酵素陽性ニューロンは10),新奇性,覚醒と睡眠,注意,ストレスなどに関与すると考えられてきた.青斑核のチロシン水酸化酵素陽性ニューロンが記憶の保持の強化を発揮する時間枠は,腹側被蓋野のチロシン水酸化酵素陽性ニューロンと比較して精度が低く,記憶の固定化の神経機構を説明する“シナプスタグ仮説”11,12) に非常によくあてはまる.今回の筆者らの研究により,われわれの日常の記憶が直前あるいは直後の新奇な体験により修飾され,その保持が強化される神経機構の一端が明らかにされた.今後,この分子基盤を明らかにすることをつうじて,日常の記憶に障害がみられる健忘症を予防または改善する新たな創薬への貢献が期待される.
文 献
- Wang, S. H., Redondo, R. L. & Morris, R. G. M.: Relevance of synaptic tagging and capture to the persistence of long-term potentiation and everyday spatial memory. Proc. Natl. Acad. Sci. USA, 107, 19537-19542 (2010)[PubMed]
- Lisman, J. E. & Grace, A. A.: The hippocampal-VTA loop: controlling the entry of information into long-term memory. Neuron, 46, 703-713 (2005)[PubMed]
- Smith, C. C. & Greene, R. W.: CNS dopamine transmission mediated by noradrenergic innervation. J. Neurosci., 32, 6072-6080 (2012)[PubMed]
- Takeuchi, T., Duszkiewicz, A. J. & Morris, R. G. M.: The synaptic plasticity and memory hypothesis: encoding, storage and persistence. Philos. Trans. R. Soc. Lond. B Biol. Sci., 369, 20130288 (2014)[PubMed]
- Wu, M. F., Gulyani, S. A., Yau, E. et al.: Locus coeruleus neurons: cessation of activity during cataplexy. Neuroscience, 91, 1389-1399 (1999)[PubMed]
- Grenhoff, J. & Svensson, T. H.: Clonidine modulates dopamine cell firing in rat ventral tegmental area. Eur. J. Pharmacol., 165, 11-18 (1989)[PubMed]
- Schultz, W. & Dickinson, A.: Neuronal coding of prediction errors. Annu. Rev. Neurosci., 23, 473-500 (2000)[PubMed]
- Rosen, Z. B., Cheung, S. & Siegelbaum, S. A.: Midbrain dopamine neurons bidirectionally regulate CA3-CA1 synaptic drive. Nat. Neurosci., 18, 1763-1771 (2015)[PubMed]
- McNamara, C. G., Tejero-Cantero, A., Trouche, S. et al.: Dopaminergic neurons promote hippocampal reactivation and spatial memory persistence. Nat. Neurosci., 17, 1658-1660 (2014)[PubMed]
- Schwarz, L. A., Miyamichi, K., Gao, X. J. et al.: Viral-genetic tracing of the input-output organization of a central noradrenaline circuit. Nature, 524, 88-92 (2015)[PubMed] [新着論文レビュー]
- Frey, U. & Morris, R. G.: Synaptic tagging and long-term potentiation. Nature, 385, 533-536 (1997)[PubMed]
- Redondo, R. L. & Morris, R. G.: Making memories last: the synaptic tagging and capture hypothesis. Nat. Rev. Neurosci., 12, 17-30 (2011)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2000年 東京大学大学院医学系研究科博士後期課程 修了,同年 同 助手を経て,2008年より英国Edinburgh大学 博士後研究員.
研究テーマ:新奇性および知識に依存した迅速な学習の神経機構および分子機構.
© 2016 竹内 倫徳 Licensed under CC 表示 2.1 日本
