小鳥の運動前野は歌を連続的にコードする
大久保達夫・Galen F. Lynch・Michale S. Fee
(米国Massachusetts Institute of Technology,Department of Brain and Cognitive Sciences,McGovern Institute for Brain Research)
email:大久保達夫
DOI: 10.7875/first.author.2016.056
Rhythmic continuous-time coding in the songbird analog of vocal motor cortex.
Galen F. Lynch, Tatsuo S. Okubo, Alexander Hanuschkin, Richard H.R. Hahnloser, Michale S. Fee
Neuron, 90, 877-892 (2016)
脳の運動野や運動前野はどのように複雑な運動を生み出すかについては,長年にわたり議論が続いている.小鳥の歌は複雑な運動の例であるが,運動前野であるHVCとよばれる部位がどのように歌を生み出すかについては,おもに2つのモデルが提案されている.すなわち,連続時間モデルはHVCの活動は歌のどの部分でも起こると主張し,ジェスチャーモデルはHVCの活動はジェスチャーの区切りのみで起こると主張する.この研究においては,この2つのモデルを比較するため,HVCから数百にわたるニューロンの活動を記録した.その結果,HVCのニューロンの活動はジェスチャーの区切りのみならず歌のどの部分においても観察された.また,HVCの活動はシラブルに同期して10 Hzのリズムで変調しており,これは歌の学習の途中である幼い小鳥において顕著であった.これらの結果から,HVCは歌の学習の初期にはリズミックな活動を示すが,成鳥になるにつれこのリズムは弱まり,より連続的に歌をコードするようになることが示された.
ヒトを含む多くの動物の行動は,複雑な運動がつぎからつぎへと生み出されることにより成り立つ.これらの複雑な運動は脳においてどのように生成されるのだろうか? これまでの研究により,複雑な運動を行うにあたっては脳のなかでも運動野や運動前野とよばれる部位の活動が重要であり1),これらの部位のニューロンは運動の特定のパラメーター(たとえば,運動の方向2))に関連した活動を示すものが多いことがわかっている.一方,最近では,運動野や運動前野のニューロンは集団としてダイナミクスにしたがい発展し,このダイナミクスが複雑な運動パターンを生成するための基底になりうるという見方も広がりつつある3).このように,運動野や運動前野の機能に関しては複数の説が存在し,現在も議論が続いている.
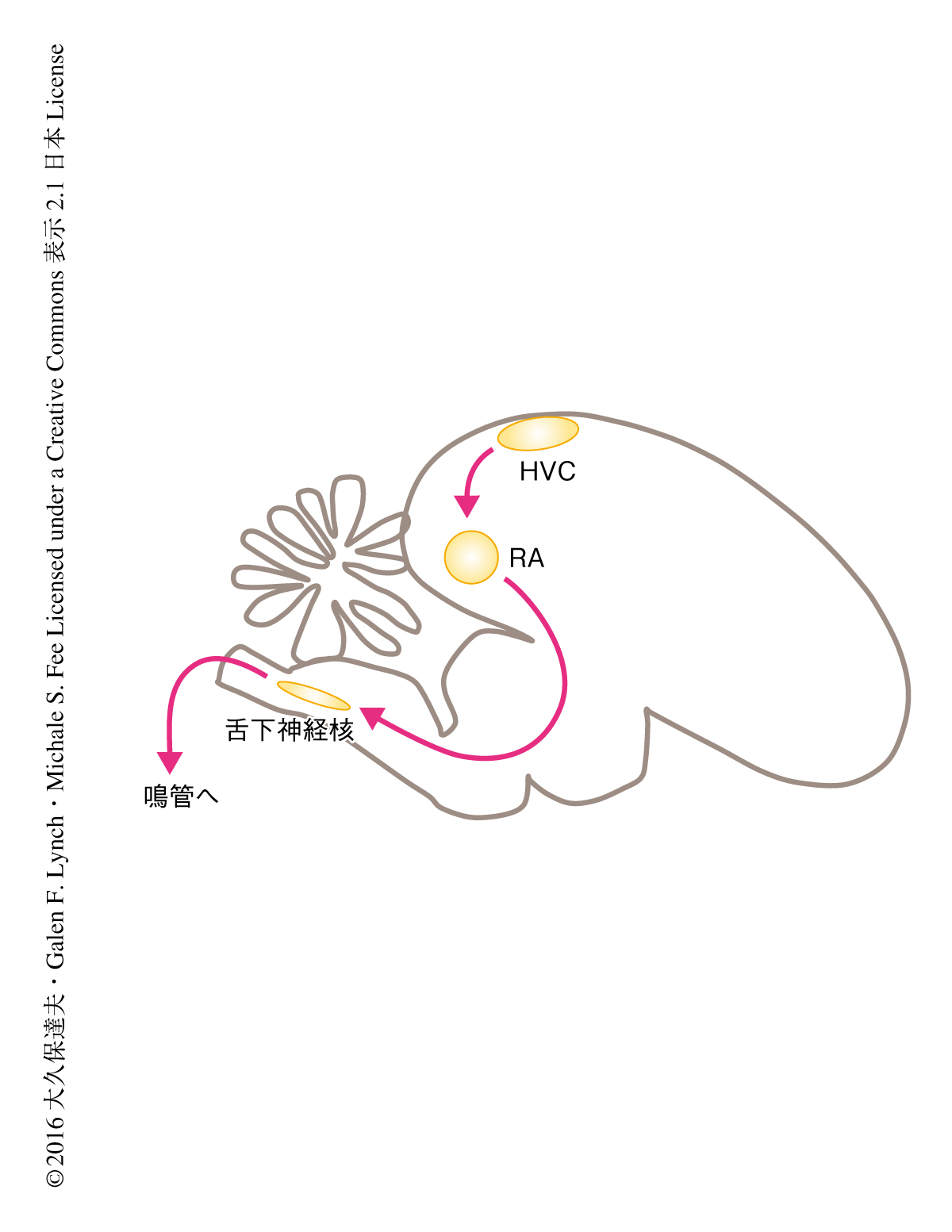
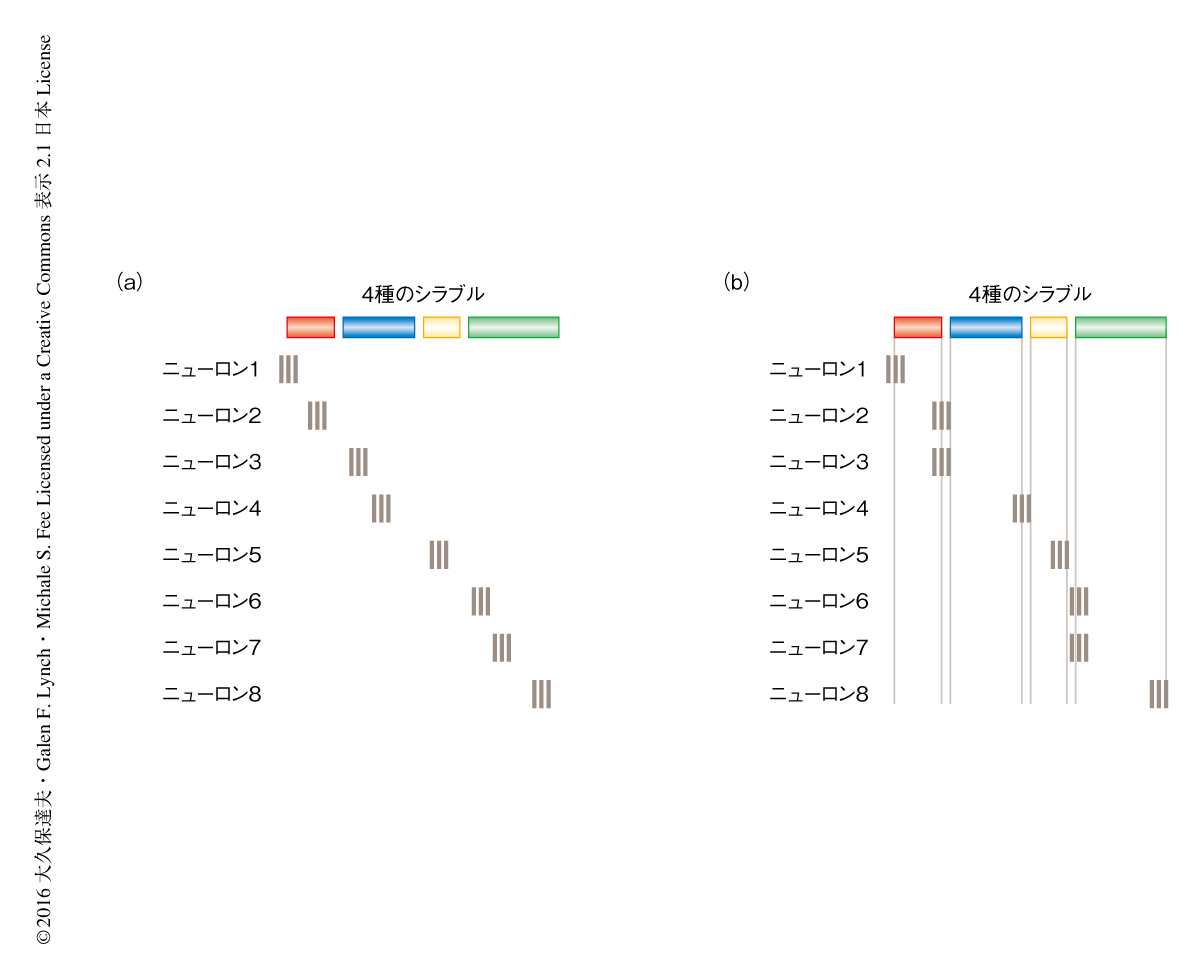
小鳥の歌は複雑な運動のよい例であるが,ここでも,運動前野の役割については意見が分かれている4,5).小鳥の脳には歌を生成するための専用の経路(図1)が存在し,この経路は運動前野であるHVCとよばれる部位にはじまり,運動野であるRAとよばれる部位をへて,鳴管の筋肉を制御する舌下神経核へとつづく.HVCの投射ニューロンは歌のごく一部において数ミリ秒のバースト状の活動を示し,それ以外は活動しない6).また,異なるニューロンは歌の異なる部分において活動する.これらの結果をもとに,HVCの個々の投射ニューロンは歌の短い部分において活動するが,全体としては歌のあらゆる部分において活動する,という連続時間モデルが提唱された6,7)(図2a).さらには,HVCの集団的な活動のパターンは複雑な歌を生成するための時間的な基底になりうるという提案もなされた8).この連続時間モデルはHVCの機能に関する重要な仮説であるが,HVCのニューロンが集団として本当に歌の全体をカバーしているかどうかは直接的には検証されていない.
一方で,近年,連続時間モデルに代わる新たなモデルとして,ジェスチャーモデルが提唱された9).このジェスチャーモデルにおいては,HVCの投射ニューロンはジェスチャーの区切りのみで活動し,それ以外は活動しないと考える(図2b).ジェスチャーの区切りは,歌のシラブルの開始時や終了時にくわえ,シラブルの途中で音響学的な特性が不連続に変わる点において起こる.ジェスチャーモデルにおいては,HVCの活動はジェスチャーの開始や終了といった運動のパラメーターに関連すると考える.
これらの2つの異なるモデルを区別するためには,同じ個体からHVCのニューロンの活動を数多く記録する必要があるが,過去の研究は,いずれも活動の記録されたニューロンの数が少なく,2つのモデルのどちらとも解釈しうるものであり,この論争に決着をつけるには不十分であった4).そこで,この研究においては,数百個にわたるニューロンの活動を記録し,その活動の様式を詳細に解析することにより,HVCがどのように歌をコードするかを明らかにすることを試みた.
HVCのニューロンが集団としてどのように活動するかを調べるため,歌っている小鳥からの細胞外記録により,計5羽の小鳥から投射ニューロンと介在ニューロンをあわせて450個のニューロンの活動を記録した.このデータを用いて,連続時間モデルのもっとも重要な仮定であるHVCの投射ニューロンは集団としては歌のどの部分においても活動することの妥当性について調べた.その結果,記録されたHVCの投射ニューロンの数が十分であった4羽の小鳥において,歌の9割以上の部分においてHVCの活動が観察された.この結果は,連続時間モデルの仮定を支持した.
HVCの投射ニューロンの集団としてのバースト状の活動がどのように分布するかについて調べた.ジェスチャーモデルが正しいのであれば,バースト状の活動はジェスチャーの区切りに密集するので同期することが多くなり,逆に,ジェスチャーの区切りのあいだにはバースト状の活動は起こらないため,長いあいだバースト状の活動がみられない部分も多くなると予想された(図2b).しかし,実際のデータにおいてはそのような傾向は観察されず,HVCのバースト状の活動は歌のあらゆる部分にわたり存在すると解釈するほうが自然であった.
HVCの投射ニューロンが活動するタイミングと歌の構造との関係を詳細に調べた.ジェスチャーモデルはHVCの活動はジェスチャーの区切りのみで起こると考えるが,この仮説の妥当性を調べるため,HVCのニューロンが活動するタイミングとジェスチャーの区切りとの関係について調べた.その結果,HVCの活動がジェスチャーの区切りのみで選択的に起こるという結果は得られなかった.
それでは,HVCの活動は歌の構造とはまったく関係がないのだろうか? この問いに答えるため,さまざまな歌のパラメーターとHVCのニューロンとの関係について解析した.その結果,HVCの介在ニューロンの活動はシラブルの開始の直前に上昇,シラブルの終了の直前に低下し,なおかつ,この活動の高低は10 Hzのリズムにより変調した.また,HVCの投射ニューロンの活動も弱いながらもこの10 Hzのリズムにより変調する傾向がみられた.このことは,HVCの活動はジェスチャーの区切りのみで起こるというジェスチャーモデルの強い命題を支持するものではなかったが,HVCの活動はシラブルの開始や終了と関係するという点においては,ジェスチャーモデルの考え方に近い結果といえた.
成鳥において観察された10 Hzのリズムはなにを意味するのだろうか? HVCの活動のパターンは歌の学習にともない大きく変わることがわかっており10)(新着論文レビュー でも掲載),成鳥において観察された10 Hzのリズムも歌の学習の結果を反映している可能性があった.その可能性を探るため,歌の学習の途中である幼い小鳥においてHVCの活動を記録した.その結果,HVCの介在ニューロンは10 Hzのリズムを示し,その活動はシラブルの開始の直前に上昇しシラブルの終了の直前に低下するという成鳥と同様のものであった.さらに,HVCの投射ニューロンにおいては成鳥よりもはるかに強い10 Hzのリズムによる変調が観察された.また,これらの10 Hzのリズムは,幼い小鳥のくり返す約100ミリ秒の短いシラブル10) と同期していた.
これらの結果から,HVCは歌の学習の初期にはシラブルに同期した10 Hzのリズミックな活動を示すが,成鳥になるにつれこのリズムの度合いは弱まり,HVCのニューロンは歌をより連続的にコードするようになることが示唆された.
小鳥の運動前野であるHVCとよばれる部位は,歌の生成および学習にかかわる脳の高次の部位ということもあり,これまで,さまざまな手法を用いてその機能が徹底的に調べられてきた.連続時間モデルはHVCの機能に関するモデルとして長きにわたりこの分野に影響をおよぼした.一方,近年,提案されたジェスチャーモデルは連続時間モデルとは大きく異なり,発表の当初から注目をあび議論をまき起こした.この論文においては,先行研究よりも大量のデータを用いてこの2つのモデルの妥当性を吟味した.その結果,HVCのバースト状の活動は歌のあらゆる部分において観察されたため,連続時間モデルの基本的な仮定が確認された.ただし,HVCの介在ニューロンにおいてシラブルに同期した活動の高低がみられたり,さらには,幼い小鳥ではその傾向が投射ニューロンにおいてもみられたりするなど,ジェスチャーモデルの考え方に近い要素も観察された.したがって,今後は,この2つのモデルをどのように統合していくかを考えていく必要がある.
なお,Neuron誌にこの論文と同時に掲載されたほかのグループの論文においては,同じ問題に相補的な手法により取り組み,頭部を固定された状態で歌っている小鳥における2光子顕微鏡を用いたイメージング法により,HVCの活動はジェスチャーの区切りのほかでも起こることを示した.さらに,HVCの投射ニューロンからの細胞内記録により,活動電位だけでなく閾値化のシナプス電位もジェスチャーの区切りのほかでも起こるとし,連続時間モデルを支持する結果が得られたと結論づけた11).
これまで,HVCの活動の様式は,小鳥の歌の学習のみならず,神経科学の全般に影響をおよぼすような知見を提供してきた.今後も,最新の技術を用いてHVCを徹底的に調べていくことにより,脳がどのように複雑な運動を生み出すかについて普遍的な原則が明らかにされることを期待したい.
略歴:2015年 米国Massachusetts Institute of Technology修了,同年より米国Harvard Medical School博士研究員.
研究テーマ:システム神経科学,神経行動学.
抱負:脳がどのようにして動物の自然な行動を生み出すかを,電気生理学をはじめとしたさまざまな方法を駆使することにより総合的に理解していきたい.
Galen F. Lynch
米国Massachusetts Institute of TechnologyにてPh.D Student.
Michale S. Fee
米国Massachusetts Institute of Technology教授.
研究室URL:http://web.mit.edu/feelab/
© 2016 大久保達夫・Galen F. Lynch・Michale S. Fee Licensed under CC 表示 2.1 日本
(米国Massachusetts Institute of Technology,Department of Brain and Cognitive Sciences,McGovern Institute for Brain Research)
email:大久保達夫
DOI: 10.7875/first.author.2016.056
Rhythmic continuous-time coding in the songbird analog of vocal motor cortex.
Galen F. Lynch, Tatsuo S. Okubo, Alexander Hanuschkin, Richard H.R. Hahnloser, Michale S. Fee
Neuron, 90, 877-892 (2016)
要 約
脳の運動野や運動前野はどのように複雑な運動を生み出すかについては,長年にわたり議論が続いている.小鳥の歌は複雑な運動の例であるが,運動前野であるHVCとよばれる部位がどのように歌を生み出すかについては,おもに2つのモデルが提案されている.すなわち,連続時間モデルはHVCの活動は歌のどの部分でも起こると主張し,ジェスチャーモデルはHVCの活動はジェスチャーの区切りのみで起こると主張する.この研究においては,この2つのモデルを比較するため,HVCから数百にわたるニューロンの活動を記録した.その結果,HVCのニューロンの活動はジェスチャーの区切りのみならず歌のどの部分においても観察された.また,HVCの活動はシラブルに同期して10 Hzのリズムで変調しており,これは歌の学習の途中である幼い小鳥において顕著であった.これらの結果から,HVCは歌の学習の初期にはリズミックな活動を示すが,成鳥になるにつれこのリズムは弱まり,より連続的に歌をコードするようになることが示された.
はじめに
ヒトを含む多くの動物の行動は,複雑な運動がつぎからつぎへと生み出されることにより成り立つ.これらの複雑な運動は脳においてどのように生成されるのだろうか? これまでの研究により,複雑な運動を行うにあたっては脳のなかでも運動野や運動前野とよばれる部位の活動が重要であり1),これらの部位のニューロンは運動の特定のパラメーター(たとえば,運動の方向2))に関連した活動を示すものが多いことがわかっている.一方,最近では,運動野や運動前野のニューロンは集団としてダイナミクスにしたがい発展し,このダイナミクスが複雑な運動パターンを生成するための基底になりうるという見方も広がりつつある3).このように,運動野や運動前野の機能に関しては複数の説が存在し,現在も議論が続いている.
小鳥の歌は複雑な運動のよい例であるが,ここでも,運動前野の役割については意見が分かれている4,5).小鳥の脳には歌を生成するための専用の経路(図1)が存在し,この経路は運動前野であるHVCとよばれる部位にはじまり,運動野であるRAとよばれる部位をへて,鳴管の筋肉を制御する舌下神経核へとつづく.HVCの投射ニューロンは歌のごく一部において数ミリ秒のバースト状の活動を示し,それ以外は活動しない6).また,異なるニューロンは歌の異なる部分において活動する.これらの結果をもとに,HVCの個々の投射ニューロンは歌の短い部分において活動するが,全体としては歌のあらゆる部分において活動する,という連続時間モデルが提唱された6,7)(図2a).さらには,HVCの集団的な活動のパターンは複雑な歌を生成するための時間的な基底になりうるという提案もなされた8).この連続時間モデルはHVCの機能に関する重要な仮説であるが,HVCのニューロンが集団として本当に歌の全体をカバーしているかどうかは直接的には検証されていない.
一方で,近年,連続時間モデルに代わる新たなモデルとして,ジェスチャーモデルが提唱された9).このジェスチャーモデルにおいては,HVCの投射ニューロンはジェスチャーの区切りのみで活動し,それ以外は活動しないと考える(図2b).ジェスチャーの区切りは,歌のシラブルの開始時や終了時にくわえ,シラブルの途中で音響学的な特性が不連続に変わる点において起こる.ジェスチャーモデルにおいては,HVCの活動はジェスチャーの開始や終了といった運動のパラメーターに関連すると考える.
これらの2つの異なるモデルを区別するためには,同じ個体からHVCのニューロンの活動を数多く記録する必要があるが,過去の研究は,いずれも活動の記録されたニューロンの数が少なく,2つのモデルのどちらとも解釈しうるものであり,この論争に決着をつけるには不十分であった4).そこで,この研究においては,数百個にわたるニューロンの活動を記録し,その活動の様式を詳細に解析することにより,HVCがどのように歌をコードするかを明らかにすることを試みた.
1.成鳥のHVCの投射ニューロンは歌のどの部分においても活動する
HVCのニューロンが集団としてどのように活動するかを調べるため,歌っている小鳥からの細胞外記録により,計5羽の小鳥から投射ニューロンと介在ニューロンをあわせて450個のニューロンの活動を記録した.このデータを用いて,連続時間モデルのもっとも重要な仮定であるHVCの投射ニューロンは集団としては歌のどの部分においても活動することの妥当性について調べた.その結果,記録されたHVCの投射ニューロンの数が十分であった4羽の小鳥において,歌の9割以上の部分においてHVCの活動が観察された.この結果は,連続時間モデルの仮定を支持した.
HVCの投射ニューロンの集団としてのバースト状の活動がどのように分布するかについて調べた.ジェスチャーモデルが正しいのであれば,バースト状の活動はジェスチャーの区切りに密集するので同期することが多くなり,逆に,ジェスチャーの区切りのあいだにはバースト状の活動は起こらないため,長いあいだバースト状の活動がみられない部分も多くなると予想された(図2b).しかし,実際のデータにおいてはそのような傾向は観察されず,HVCのバースト状の活動は歌のあらゆる部分にわたり存在すると解釈するほうが自然であった.
2.HVCの活動と歌の構造との関係
HVCの投射ニューロンが活動するタイミングと歌の構造との関係を詳細に調べた.ジェスチャーモデルはHVCの活動はジェスチャーの区切りのみで起こると考えるが,この仮説の妥当性を調べるため,HVCのニューロンが活動するタイミングとジェスチャーの区切りとの関係について調べた.その結果,HVCの活動がジェスチャーの区切りのみで選択的に起こるという結果は得られなかった.
それでは,HVCの活動は歌の構造とはまったく関係がないのだろうか? この問いに答えるため,さまざまな歌のパラメーターとHVCのニューロンとの関係について解析した.その結果,HVCの介在ニューロンの活動はシラブルの開始の直前に上昇,シラブルの終了の直前に低下し,なおかつ,この活動の高低は10 Hzのリズムにより変調した.また,HVCの投射ニューロンの活動も弱いながらもこの10 Hzのリズムにより変調する傾向がみられた.このことは,HVCの活動はジェスチャーの区切りのみで起こるというジェスチャーモデルの強い命題を支持するものではなかったが,HVCの活動はシラブルの開始や終了と関係するという点においては,ジェスチャーモデルの考え方に近い結果といえた.
3.幼い小鳥のHVCはシラブルに同期したリズミカルな活動を示す
成鳥において観察された10 Hzのリズムはなにを意味するのだろうか? HVCの活動のパターンは歌の学習にともない大きく変わることがわかっており10)(新着論文レビュー でも掲載),成鳥において観察された10 Hzのリズムも歌の学習の結果を反映している可能性があった.その可能性を探るため,歌の学習の途中である幼い小鳥においてHVCの活動を記録した.その結果,HVCの介在ニューロンは10 Hzのリズムを示し,その活動はシラブルの開始の直前に上昇しシラブルの終了の直前に低下するという成鳥と同様のものであった.さらに,HVCの投射ニューロンにおいては成鳥よりもはるかに強い10 Hzのリズムによる変調が観察された.また,これらの10 Hzのリズムは,幼い小鳥のくり返す約100ミリ秒の短いシラブル10) と同期していた.
これらの結果から,HVCは歌の学習の初期にはシラブルに同期した10 Hzのリズミックな活動を示すが,成鳥になるにつれこのリズムの度合いは弱まり,HVCのニューロンは歌をより連続的にコードするようになることが示唆された.
おわりに
小鳥の運動前野であるHVCとよばれる部位は,歌の生成および学習にかかわる脳の高次の部位ということもあり,これまで,さまざまな手法を用いてその機能が徹底的に調べられてきた.連続時間モデルはHVCの機能に関するモデルとして長きにわたりこの分野に影響をおよぼした.一方,近年,提案されたジェスチャーモデルは連続時間モデルとは大きく異なり,発表の当初から注目をあび議論をまき起こした.この論文においては,先行研究よりも大量のデータを用いてこの2つのモデルの妥当性を吟味した.その結果,HVCのバースト状の活動は歌のあらゆる部分において観察されたため,連続時間モデルの基本的な仮定が確認された.ただし,HVCの介在ニューロンにおいてシラブルに同期した活動の高低がみられたり,さらには,幼い小鳥ではその傾向が投射ニューロンにおいてもみられたりするなど,ジェスチャーモデルの考え方に近い要素も観察された.したがって,今後は,この2つのモデルをどのように統合していくかを考えていく必要がある.
なお,Neuron誌にこの論文と同時に掲載されたほかのグループの論文においては,同じ問題に相補的な手法により取り組み,頭部を固定された状態で歌っている小鳥における2光子顕微鏡を用いたイメージング法により,HVCの活動はジェスチャーの区切りのほかでも起こることを示した.さらに,HVCの投射ニューロンからの細胞内記録により,活動電位だけでなく閾値化のシナプス電位もジェスチャーの区切りのほかでも起こるとし,連続時間モデルを支持する結果が得られたと結論づけた11).
これまで,HVCの活動の様式は,小鳥の歌の学習のみならず,神経科学の全般に影響をおよぼすような知見を提供してきた.今後も,最新の技術を用いてHVCを徹底的に調べていくことにより,脳がどのように複雑な運動を生み出すかについて普遍的な原則が明らかにされることを期待したい.
文 献
- Tanji, J.: Sequential organization of multiple movements: involvement of cortical motor areas. Annu. Rev. Neurosci., 24, 631-651 (2001)[PubMed]
- Georgopoulos, A. P., Schwartz, A. B. & Kettner, R. E.: Neuronal population coding of movement direction. Science, 233, 1416-1419 (1986)[PubMed]
- Shenoy, K. V, Sahani, M. & Churchland, M. M.: Cortical control of arm movements: a dynamical systems perspective. Annu. Rev. Neurosci., 36, 337-359 (2013)[PubMed]
- Troyer, T. W.: The units of a song. Nature, 495, 56-57 (2013)[PubMed]
- Troyer, T. W.: Continuous time representations of song in zebra finches. Neuron, 90, 672-674 (2016)[PubMed]
- Hahnloser, R. H. R., Kozhevnikov, A. A. & Fee, M. S.: An ultra-sparse code underlies the generation of neural sequences in a songbird. Nature, 419, 65-70 (2002)[PubMed]
- Fee, M. S., Kozhevnikov, A. A. & Hahnloser, R. H. R.: Neural mechanisms of vocal sequence: generation in the songbird. Ann. N. Y. Acad. Sci., 1016, 153-170 (2004)[PubMed]
- Leonardo, A. & Fee, M. S.: Ensemble coding of vocal control in birdsong. J. Neurosci., 25, 652-661 (2005)[PubMed]
- Amador, A., Perl, Y. S., Mindlin, G. B. et al.: Elemental gesture dynamics are encoded by song premotor cortical neurons. Nature, 495, 59-64 (2014)[PubMed]
- Okubo, T. S., Mackevicius, E. L., Payne, H. L. et al.: Growth and splitting of neural sequences in songbird vocal development. Nature, 528, 352-357 (2015)[PubMed] [新着論文レビュー]
- Picardo, M. A., Merel, J., Katlowitz, K. A. et al.: Population-level representation of a temporal sequence underlying song production in the zebra finch. Neuron, 90, 866-876 (2016)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2015年 米国Massachusetts Institute of Technology修了,同年より米国Harvard Medical School博士研究員.
研究テーマ:システム神経科学,神経行動学.
抱負:脳がどのようにして動物の自然な行動を生み出すかを,電気生理学をはじめとしたさまざまな方法を駆使することにより総合的に理解していきたい.
Galen F. Lynch
米国Massachusetts Institute of TechnologyにてPh.D Student.
Michale S. Fee
米国Massachusetts Institute of Technology教授.
研究室URL:http://web.mit.edu/feelab/
© 2016 大久保達夫・Galen F. Lynch・Michale S. Fee Licensed under CC 表示 2.1 日本
