恐怖記憶の形成および消去には異なるタイプの青斑核のノルアドレナリンニューロンが関与する
植松 朗・Bao Zhen Tan・Joshua P. Johansen
(理化学研究所脳科学総合研究センター 記憶神経回路研究チーム)
email:植松 朗
DOI: 10.7875/first.author.2017.114
Modular organization of the brainstem noradrenaline system coordinates opposing learning states.
Akira Uematsu, Bao Zhen Tan, Edgar A. Ycu, Jessica Sulkes Cuevas, Jenny Koivumaa, Felix Junyent, Eric J. Kremer, Ilana B. Witten, Karl Deisseroth, Joshua P. Johansen
Nature Neuroscience, 20, 1602-1611 (2017)
恐怖記憶の形成および消去にはそれぞれ扁桃体および内側前頭前野が関与し,ノルアドレナリンシグナルが重要であることが知られている.ノルアドレナリンニューロンはおもに青斑核のノルアドレナリンニューロンから脳のさまざまな領域に投射しており,青斑核のノルアドレナリンニューロンは機能的に一様であると考えられていた.しかしながら,青斑核のノルアドレナリンニューロンの恐怖記憶の形成および消去における役割は明らかではなかった.筆者らは,扁桃体あるいは内側前頭前野に投射する青斑核のニューロンに着目し,それぞれは解剖学的に異なるタイプのニューロンであることを見い出した.光遺伝学的な手法により,扁桃体に投射するニューロンは恐怖記憶の形成に,内側前頭前野に投射するニューロンは恐怖記憶の消去に関与することが明らかにされた.さらに,扁桃体に投射するニューロンおよび内側前頭前野に投射するニューロンは特定の領域に選択的に投射していた.以上のことから,青斑核のノルアドレナリンニューロンは一様の機能をもつと考えられてきたが,恐怖記憶の形成および消去においては投射する部位により異なる機能を示すことが明らかにされた.
恐怖の体験を記憶することは危険の予知などにつながることから,われわれの生活に必要な生存本能の一部である.一方で,恐怖記憶が不要になると消去学習により上書きされ,恐怖記憶は弱まることが知られている.先行研究により,恐怖条件づけにおいては扁桃体が,また,消去学習においては内側前頭前野の腹側部が重要であることが明らかにされている1,2).また,薬理学的な実験により,神経伝達物質であるノルアドレナリンは,恐怖条件づけにおいては扁桃体の基底外側核で,また,消去学習においては内側前頭前野の腹側部で重要な役割をはたすことが示されている3-5).
ノルアドレナリンは脳幹にある青斑核のノルアドレナリンニューロンからおもに分泌されることが知られている.青斑核に存在するニューロンはほとんどがノルアドレナリンニューロンであるが,このノルアドレナリンニューロンのひとつひとつがさまざまな領域に一様に分布し,音などの外部からの刺激に対し一様な応答を示すと考えられていた6-8).ノルアドレナリンニューロンは不安や睡眠といったさまざまなことに関係することが示されていたものの9,10),恐怖条件づけあるいは消去学習における青斑核のノルアドレナリンニューロンのはたらきについてはほとんど明らかにされていなかった.この研究においては,青斑核のノルアドレナリンニューロンの恐怖記憶の形成および消去における役割を光遺伝学的な手法および電気生理学的な手法を用いて明らかにすることを目的とした.
カテコールアミンニューロンにおいてCreを発現する遺伝子改変ラットを使用した11).光遺伝学的な操作のため,Creに特異的にアーキロドプシンTを発現させるアデノ随伴ウイルスベクターを青斑核に投与し,青斑核の上部に光ファイバーを留置した.アーキロドプシンTは光感受性のプロトンポンプで,波長570 nm付近の光を照射することにより神経活動を抑制することができる.このラットを手術から3週間後に行動実験に供試した.恐怖条件づけにおいては,恐怖を誘発しない音の刺激を提示したのち,恐怖体験として脚に電気ショックをあたえ,翌日に5回の音の刺激を提示した.消去学習においては,恐怖体験として電気ショックをあたえた翌日に22回の音の刺激を提示し,さらにその翌日に5回の音の刺激を提示した.
恐怖条件づけにおいて音の刺激を提示したとき青斑核に光を照射して神経活動を抑制したところ,翌日の音の刺激に対するすくみ応答には対照と比べて差はみられなかった.一方,恐怖条件づけにおいて電気ショックをあたえたとき青斑核に光を照射して神経活動を抑制したところ,翌日の音の刺激に対するすくみ応答は対照と比べ有意に減少した.以上のことから,恐怖記憶の形成においては,恐怖体験のあいだの青斑核のノルアドレナリンニューロンの活動が重要であることが明らかにされた.また,消去学習において音の刺激を提示したとき青斑核に光を照射して神経活動を抑制したところ,翌日の音の刺激に対するすくみ応答は対照と比べ有意に増加した.このことから,恐怖記憶の消去においては,音の刺激に対する青斑核のノルアドレナリンニューロンの活動が必要であることが示唆された.
ラットの青斑核に電極を留置することによりノルアドレナリンニューロンの活動を電気生理学的に測定した.青斑核のニューロンはほとんどがノルアドレナリンニューロンであるため,測定ののち組織切片において電極の位置を確かめ,青斑核に位置していた場合にはノルアドレナリンニューロンから記録されていると仮定した.その結果,電気ショックに対してはほとんどのノルアドレナリンニューロンが強い応答を示した.その一方,恐怖条件づけののち音の刺激に対するすくみ応答が高いときには,記録したニューロンのうち一部のニューロンのみが応答した(恐怖細胞).そして,消去学習が進むと恐怖細胞とは異なるニューロンが応答するようになった(消去細胞).以上のことから,これまで考えられていたように,ノルアドレナリンニューロンは一様の応答を示すわけではなく,状況により異なるノルアドレナリンニューロンが応答を示すことが明らかにされた.
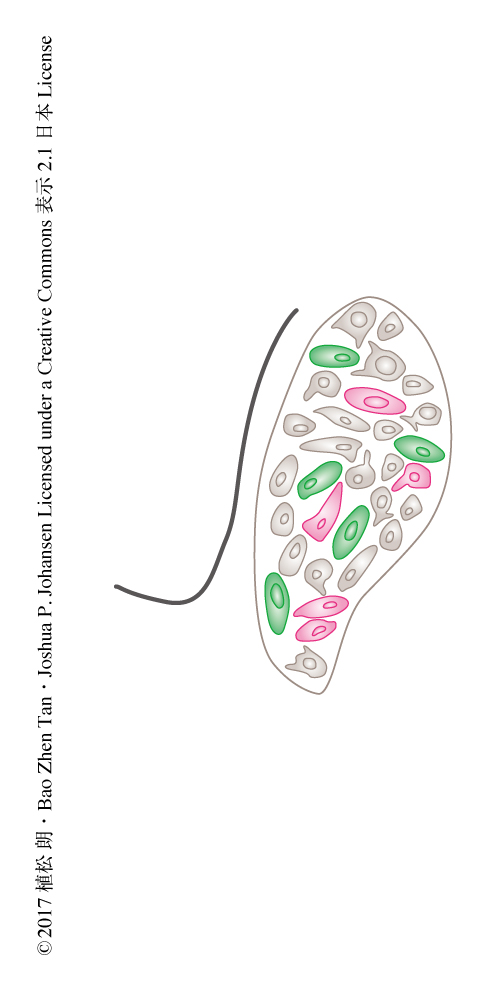
ノルアドレナリンは扁桃体の基底外側核においては恐怖記憶の形成に,内側前頭前野の腹側部においては恐怖記憶の消去に関与する.そこで,恐怖を誘発しない刺激に対する異なる神経活動は投射先により異なるという仮説をたてた.青斑核には投射先により異なるニューロンがあるかどうか検証するため,扁桃体の基底外側核および内側前頭前野の腹側部に異なる逆行性の蛍光色素を投与した.これらの逆行性の蛍光色素は軸索より取り込まれ細胞体へと輸送される.その結果,青斑核のニューロンにおいてそれぞれの蛍光はほとんどが重なることはなく,扁桃体の基底外側核に投射するニューロンと内側前頭前野の腹側部に投射するニューロンは異なる独立したニューロンであることが明らかにされた(図1).
逆行性の蛍光色素により扁桃体の基底外側核に投射するニューロンおよび内側前頭前野の腹側部に投射するニューロンを標識し,恐怖条件づけの翌日に5回あるいは22回の恐怖を誘発しない音の刺激を提示したのち,神経活動のマーカーであるc-Fosを抗体染色した.その結果,恐怖条件づけののち,どちらにも同等のc-Fos陽性ニューロンが発現していた.しかし,刺激を5回提示し恐怖を想起させたラットにおいては,c-Fos陽性のニューロンは内側前頭前野の腹側部に投射するニューロンに比べ扁桃体の基底外側核へと投射するニューロンにおいて有意に多かった.一方で,刺激を22回提示し恐怖を消去したラットにおいては,c-Fos陽性のニューロンは内側前頭前野の腹側部に投射するニューロンにおいて有意に多かった.c-Fosの免疫染色は時間分解能が低いため,どの時点でこれらのニューロンが応答したのかは定かではないが,これらの結果から,電気生理学的な実験においてみられた恐怖細胞および消去細胞の活動との関係性が示唆された.
扁桃体あるいは内側前頭前野に投射するニューロンの機能をみるために光遺伝学的な手法を用いた.ラットの扁桃体あるいは内側前頭前野に逆行性に輸送される組換え狂犬病ウイルスを投与し,青斑核の上部に光ファイバーを留置した.この組換え狂犬病ウイルスはウイルス複製因子の遺伝子を除いた代わりにアーキロドプシンTの遺伝子を挿入しているため,投与した部位に投射するニューロンの軸索より逆行性に輸送され細胞体にアーキロドプシンTを発現させる.ウイルス複製因子をもたないため,そののち増殖あるいは転移することはない.その結果,青斑核の周囲においてアーキロドプシンTを発現するニューロンはほとんどがノルアドレナリンニューロンであった.恐怖条件づけにおいて電気ショックをあたえたときのみ光を照射して神経活動を抑制したところ,扁桃体に投射するニューロンの活動を抑制したときには翌日の音の刺激に対するすくみ応答は対照と比べ有意に減少したが,内側前頭前野に投射するニューロンの活動を抑制したときには変化はみられなかった.一方,消去学習において音の刺激を提示したときに光を照射して神経活動を抑制したところ,扁桃体に投射するニューロンの活動を抑制したときには翌日の音の刺激に対するすくみ応答が対照と比べ有意に減少したが,内側前頭前野に投射するニューロンの活動を抑制したときには翌日の音の刺激に対するすくみ応答は対照に比べ有意に増加した.これらの結果から,恐怖記憶の形成においては扁桃体に投射するノルアドレナリンニューロンが重要であること,恐怖記憶の消去においては扁桃体に投射するニューロンと内側前頭前野に投射するニューロンは正反対の役割をもつことが明らかにされた.
青斑核のノルアドレナリンニューロンについては1つのタイプのニューロンが脳のさまざまな領域に投射すると考えられていた.しかしながら,逆行性の蛍光色素による標識により,青斑核には投射先により異なるタイプのニューロンのあることが見い出されたことから,これらのニューロンのさまざまな部位への投射について,多くの領域に一様に投射するのか,それとも,選択的な部位への投射があるのか観察した.逆行性に輸送されるイヌアデノウイルスを扁桃体の基底外側核あるいは内側前頭前野の腹側部に投与し,青斑核にCreに特異的にシナプトフィジンを発現させるアデノウイルスベクターを投与した.シナプトフィジンは神経伝達物質の放出に関係しシナプス終末に多く存在するタンパク質であり,これに蛍光タンパク質を融合したものを発現させることにより脳のさまざまな部位において軸索末端を定量した.その結果,扁桃体の基底外側核に投射するニューロンあるいは内側前頭前野の腹側部に投射するニューロンは,脳のさまざまな部位に軸索末端が認められたものの,扁桃体の基底外側核に投射するニューロンは基底外側核,中心核,内側核といった扁桃体の全体に軸索末端が多く確認され,一方で,内側前頭前野の腹側部に投射するニューロンは腹側部にも背側部にも軸索末端が多くあることが明らかにされた.以上のことから,扁桃体あるいは内側前頭前野に投射するニューロンは,どちらも脳のさまざまな領域に広く投射するものの,選択的に多く投射する部位のあることが明らかにされた.
この研究においては,青斑核のノルアドレナリンニューロンは恐怖記憶の形成および消去のどちらにも重要であること,青斑核のノルアドレナリンニューロンは状況により異なる神経活動を示すこと,扁桃体あるいは内側前頭前野に投射する異なるタイプのニューロンの存在することが見い出された.さらに,扁桃体に投射するニューロンは恐怖記憶の形成に必要で恐怖記憶の消去を阻害する役割のあること,また,内側前頭前野に投射するニューロンは恐怖記憶の消去に重要な役割をはたすことが明らかにされた.
過度な恐怖は心的外傷後ストレス障害といった不安障害をひき起こす.また,不安障害などの精神疾患に対して,一部でノルアドレナリンを標的とした治療薬が用いられている.今後,この研究を発展させ,この研究において見い出された異なるタイプのノルアドレナリンニューロンに特異的な遺伝子マーカーを同定し選択的に標的とすることができれば,不安障害などの精神疾患に対しより効果的な治療法の開発につながるものと期待される.
略歴:2011年 東京大学大学院農学生命科学研究科 修了,味の素 研究員を経て,2013年より理化学研究所脳科学総合研究センター 研究員.
研究テーマ:学習におけるカテコールアミンニューロンの役割.
Bao Zhen Tan
理化学研究所脳科学総合研究センター 研究員.
Joshua P. Johansen
理化学研究所脳科学総合研究センター チームリーダー.
研究室URL:http://jlab.brain.riken.jp/
© 2017 植松 朗・Bao Zhen Tan・Joshua P. Johansen Licensed under CC 表示 2.1 日本
(理化学研究所脳科学総合研究センター 記憶神経回路研究チーム)
email:植松 朗
DOI: 10.7875/first.author.2017.114
Modular organization of the brainstem noradrenaline system coordinates opposing learning states.
Akira Uematsu, Bao Zhen Tan, Edgar A. Ycu, Jessica Sulkes Cuevas, Jenny Koivumaa, Felix Junyent, Eric J. Kremer, Ilana B. Witten, Karl Deisseroth, Joshua P. Johansen
Nature Neuroscience, 20, 1602-1611 (2017)
要 約
恐怖記憶の形成および消去にはそれぞれ扁桃体および内側前頭前野が関与し,ノルアドレナリンシグナルが重要であることが知られている.ノルアドレナリンニューロンはおもに青斑核のノルアドレナリンニューロンから脳のさまざまな領域に投射しており,青斑核のノルアドレナリンニューロンは機能的に一様であると考えられていた.しかしながら,青斑核のノルアドレナリンニューロンの恐怖記憶の形成および消去における役割は明らかではなかった.筆者らは,扁桃体あるいは内側前頭前野に投射する青斑核のニューロンに着目し,それぞれは解剖学的に異なるタイプのニューロンであることを見い出した.光遺伝学的な手法により,扁桃体に投射するニューロンは恐怖記憶の形成に,内側前頭前野に投射するニューロンは恐怖記憶の消去に関与することが明らかにされた.さらに,扁桃体に投射するニューロンおよび内側前頭前野に投射するニューロンは特定の領域に選択的に投射していた.以上のことから,青斑核のノルアドレナリンニューロンは一様の機能をもつと考えられてきたが,恐怖記憶の形成および消去においては投射する部位により異なる機能を示すことが明らかにされた.
はじめに
恐怖の体験を記憶することは危険の予知などにつながることから,われわれの生活に必要な生存本能の一部である.一方で,恐怖記憶が不要になると消去学習により上書きされ,恐怖記憶は弱まることが知られている.先行研究により,恐怖条件づけにおいては扁桃体が,また,消去学習においては内側前頭前野の腹側部が重要であることが明らかにされている1,2).また,薬理学的な実験により,神経伝達物質であるノルアドレナリンは,恐怖条件づけにおいては扁桃体の基底外側核で,また,消去学習においては内側前頭前野の腹側部で重要な役割をはたすことが示されている3-5).
ノルアドレナリンは脳幹にある青斑核のノルアドレナリンニューロンからおもに分泌されることが知られている.青斑核に存在するニューロンはほとんどがノルアドレナリンニューロンであるが,このノルアドレナリンニューロンのひとつひとつがさまざまな領域に一様に分布し,音などの外部からの刺激に対し一様な応答を示すと考えられていた6-8).ノルアドレナリンニューロンは不安や睡眠といったさまざまなことに関係することが示されていたものの9,10),恐怖条件づけあるいは消去学習における青斑核のノルアドレナリンニューロンのはたらきについてはほとんど明らかにされていなかった.この研究においては,青斑核のノルアドレナリンニューロンの恐怖記憶の形成および消去における役割を光遺伝学的な手法および電気生理学的な手法を用いて明らかにすることを目的とした.
1.恐怖記憶の形成および消去には青斑核のノルアドレナリンニューロンが重要である
カテコールアミンニューロンにおいてCreを発現する遺伝子改変ラットを使用した11).光遺伝学的な操作のため,Creに特異的にアーキロドプシンTを発現させるアデノ随伴ウイルスベクターを青斑核に投与し,青斑核の上部に光ファイバーを留置した.アーキロドプシンTは光感受性のプロトンポンプで,波長570 nm付近の光を照射することにより神経活動を抑制することができる.このラットを手術から3週間後に行動実験に供試した.恐怖条件づけにおいては,恐怖を誘発しない音の刺激を提示したのち,恐怖体験として脚に電気ショックをあたえ,翌日に5回の音の刺激を提示した.消去学習においては,恐怖体験として電気ショックをあたえた翌日に22回の音の刺激を提示し,さらにその翌日に5回の音の刺激を提示した.
恐怖条件づけにおいて音の刺激を提示したとき青斑核に光を照射して神経活動を抑制したところ,翌日の音の刺激に対するすくみ応答には対照と比べて差はみられなかった.一方,恐怖条件づけにおいて電気ショックをあたえたとき青斑核に光を照射して神経活動を抑制したところ,翌日の音の刺激に対するすくみ応答は対照と比べ有意に減少した.以上のことから,恐怖記憶の形成においては,恐怖体験のあいだの青斑核のノルアドレナリンニューロンの活動が重要であることが明らかにされた.また,消去学習において音の刺激を提示したとき青斑核に光を照射して神経活動を抑制したところ,翌日の音の刺激に対するすくみ応答は対照と比べ有意に増加した.このことから,恐怖記憶の消去においては,音の刺激に対する青斑核のノルアドレナリンニューロンの活動が必要であることが示唆された.
2.恐怖記憶の形成および消去においては異なるノルアドレナリンニューロンが活動する
ラットの青斑核に電極を留置することによりノルアドレナリンニューロンの活動を電気生理学的に測定した.青斑核のニューロンはほとんどがノルアドレナリンニューロンであるため,測定ののち組織切片において電極の位置を確かめ,青斑核に位置していた場合にはノルアドレナリンニューロンから記録されていると仮定した.その結果,電気ショックに対してはほとんどのノルアドレナリンニューロンが強い応答を示した.その一方,恐怖条件づけののち音の刺激に対するすくみ応答が高いときには,記録したニューロンのうち一部のニューロンのみが応答した(恐怖細胞).そして,消去学習が進むと恐怖細胞とは異なるニューロンが応答するようになった(消去細胞).以上のことから,これまで考えられていたように,ノルアドレナリンニューロンは一様の応答を示すわけではなく,状況により異なるノルアドレナリンニューロンが応答を示すことが明らかにされた.
3.投射先により異なるノルアドレナリンニューロン
ノルアドレナリンは扁桃体の基底外側核においては恐怖記憶の形成に,内側前頭前野の腹側部においては恐怖記憶の消去に関与する.そこで,恐怖を誘発しない刺激に対する異なる神経活動は投射先により異なるという仮説をたてた.青斑核には投射先により異なるニューロンがあるかどうか検証するため,扁桃体の基底外側核および内側前頭前野の腹側部に異なる逆行性の蛍光色素を投与した.これらの逆行性の蛍光色素は軸索より取り込まれ細胞体へと輸送される.その結果,青斑核のニューロンにおいてそれぞれの蛍光はほとんどが重なることはなく,扁桃体の基底外側核に投射するニューロンと内側前頭前野の腹側部に投射するニューロンは異なる独立したニューロンであることが明らかにされた(図1).
逆行性の蛍光色素により扁桃体の基底外側核に投射するニューロンおよび内側前頭前野の腹側部に投射するニューロンを標識し,恐怖条件づけの翌日に5回あるいは22回の恐怖を誘発しない音の刺激を提示したのち,神経活動のマーカーであるc-Fosを抗体染色した.その結果,恐怖条件づけののち,どちらにも同等のc-Fos陽性ニューロンが発現していた.しかし,刺激を5回提示し恐怖を想起させたラットにおいては,c-Fos陽性のニューロンは内側前頭前野の腹側部に投射するニューロンに比べ扁桃体の基底外側核へと投射するニューロンにおいて有意に多かった.一方で,刺激を22回提示し恐怖を消去したラットにおいては,c-Fos陽性のニューロンは内側前頭前野の腹側部に投射するニューロンにおいて有意に多かった.c-Fosの免疫染色は時間分解能が低いため,どの時点でこれらのニューロンが応答したのかは定かではないが,これらの結果から,電気生理学的な実験においてみられた恐怖細胞および消去細胞の活動との関係性が示唆された.
4.投射先により異なる機能をもつノルアドレナリンニューロン
扁桃体あるいは内側前頭前野に投射するニューロンの機能をみるために光遺伝学的な手法を用いた.ラットの扁桃体あるいは内側前頭前野に逆行性に輸送される組換え狂犬病ウイルスを投与し,青斑核の上部に光ファイバーを留置した.この組換え狂犬病ウイルスはウイルス複製因子の遺伝子を除いた代わりにアーキロドプシンTの遺伝子を挿入しているため,投与した部位に投射するニューロンの軸索より逆行性に輸送され細胞体にアーキロドプシンTを発現させる.ウイルス複製因子をもたないため,そののち増殖あるいは転移することはない.その結果,青斑核の周囲においてアーキロドプシンTを発現するニューロンはほとんどがノルアドレナリンニューロンであった.恐怖条件づけにおいて電気ショックをあたえたときのみ光を照射して神経活動を抑制したところ,扁桃体に投射するニューロンの活動を抑制したときには翌日の音の刺激に対するすくみ応答は対照と比べ有意に減少したが,内側前頭前野に投射するニューロンの活動を抑制したときには変化はみられなかった.一方,消去学習において音の刺激を提示したときに光を照射して神経活動を抑制したところ,扁桃体に投射するニューロンの活動を抑制したときには翌日の音の刺激に対するすくみ応答が対照と比べ有意に減少したが,内側前頭前野に投射するニューロンの活動を抑制したときには翌日の音の刺激に対するすくみ応答は対照に比べ有意に増加した.これらの結果から,恐怖記憶の形成においては扁桃体に投射するノルアドレナリンニューロンが重要であること,恐怖記憶の消去においては扁桃体に投射するニューロンと内側前頭前野に投射するニューロンは正反対の役割をもつことが明らかにされた.
5.特定のノルアドレナリンニューロンの選択的な領域への投射
青斑核のノルアドレナリンニューロンについては1つのタイプのニューロンが脳のさまざまな領域に投射すると考えられていた.しかしながら,逆行性の蛍光色素による標識により,青斑核には投射先により異なるタイプのニューロンのあることが見い出されたことから,これらのニューロンのさまざまな部位への投射について,多くの領域に一様に投射するのか,それとも,選択的な部位への投射があるのか観察した.逆行性に輸送されるイヌアデノウイルスを扁桃体の基底外側核あるいは内側前頭前野の腹側部に投与し,青斑核にCreに特異的にシナプトフィジンを発現させるアデノウイルスベクターを投与した.シナプトフィジンは神経伝達物質の放出に関係しシナプス終末に多く存在するタンパク質であり,これに蛍光タンパク質を融合したものを発現させることにより脳のさまざまな部位において軸索末端を定量した.その結果,扁桃体の基底外側核に投射するニューロンあるいは内側前頭前野の腹側部に投射するニューロンは,脳のさまざまな部位に軸索末端が認められたものの,扁桃体の基底外側核に投射するニューロンは基底外側核,中心核,内側核といった扁桃体の全体に軸索末端が多く確認され,一方で,内側前頭前野の腹側部に投射するニューロンは腹側部にも背側部にも軸索末端が多くあることが明らかにされた.以上のことから,扁桃体あるいは内側前頭前野に投射するニューロンは,どちらも脳のさまざまな領域に広く投射するものの,選択的に多く投射する部位のあることが明らかにされた.
おわりに
この研究においては,青斑核のノルアドレナリンニューロンは恐怖記憶の形成および消去のどちらにも重要であること,青斑核のノルアドレナリンニューロンは状況により異なる神経活動を示すこと,扁桃体あるいは内側前頭前野に投射する異なるタイプのニューロンの存在することが見い出された.さらに,扁桃体に投射するニューロンは恐怖記憶の形成に必要で恐怖記憶の消去を阻害する役割のあること,また,内側前頭前野に投射するニューロンは恐怖記憶の消去に重要な役割をはたすことが明らかにされた.
過度な恐怖は心的外傷後ストレス障害といった不安障害をひき起こす.また,不安障害などの精神疾患に対して,一部でノルアドレナリンを標的とした治療薬が用いられている.今後,この研究を発展させ,この研究において見い出された異なるタイプのノルアドレナリンニューロンに特異的な遺伝子マーカーを同定し選択的に標的とすることができれば,不安障害などの精神疾患に対しより効果的な治療法の開発につながるものと期待される.
文 献
- LeDoux, J. E.: Emotion circuits in the brain. Annu. Rev. Neurosci., 23, 155-184 (2000)[PubMed]
- Herry, C. & Johansen, J. P.: Encoding of fear learning and memory in distributed neuronal circuits. Nat. Neurosci., 17, 1644-1654 (2014)[PubMed]
- Bush, D.E. Caparosa, E. M., Gekker, A. et al.: Beta-adrenergic receptors in the lateral nucleus of the amygdala contribute to the acquisition but not the consolidation of auditory fear conditioning. Front. Behav. Neurosci., 4, 154 (2010)[PubMed]
- Johansen, J. P., Diaz-Mataix, L., Hamanaka, H. et al.: Hebbian and neuromodulatory mechanisms interact to trigger associative memory formation. Proc. Natl. Acad. Sci. USA, 111, E5584-E5592 (2014)[PubMed]
- Mueller, D., Porter, J. T. & Quirk, G. J.: Noradrenergic signaling in infralimbic cortex increases cell excitability and strengthens memory for fear extinction. J. Neurosci., 28, 369-375 (2008)[PubMed]
- Sara, S. J. & Bouret, S.: Orienting and reorienting: the locus coeruleus mediates cognition through arousal. Neuron, 76, 130-141 (2012)[PubMed]
- Aston-Jones, G. & Cohen, J. D.: An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu. Rev. Neurosci., 28, 403-450 (2005)[PubMed]
- Schwarz, L. A., Miyamichi, K., Gao, X. J. et al.: Viral-genetic tracing of the input-output organization of a central noradrenaline circuit. Nature, 524, 88-92 (2015)[PubMed] [新着論文レビュー]
- Arnsten, A. F.: Stress signalling pathways that impair prefrontal cortex structure and function. Nat. Rev. Neurosci., 10, 410-422 (2009)[PubMed]
- Uematsu, A., Tan, B. Z. & Johansen, J. P.: Projection specificity in heterogeneous locus coeruleus cell populations: implications for learning and memory. Learn. Mem., 22, 444-451 (2015)[PubMed]
- Witten, I. B., Steinberg, E. E., Lee, S. Y. et al.: Recombinase-driver rat lines: tools, techniques, and optogenetic application to dopamine-mediated reinforcement. Neuron, 72, 721-733 (2011)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2011年 東京大学大学院農学生命科学研究科 修了,味の素 研究員を経て,2013年より理化学研究所脳科学総合研究センター 研究員.
研究テーマ:学習におけるカテコールアミンニューロンの役割.
Bao Zhen Tan
理化学研究所脳科学総合研究センター 研究員.
Joshua P. Johansen
理化学研究所脳科学総合研究センター チームリーダー.
研究室URL:http://jlab.brain.riken.jp/
© 2017 植松 朗・Bao Zhen Tan・Joshua P. Johansen Licensed under CC 表示 2.1 日本
