アブラナ科植物において自他を認識する遺伝子の遺伝子重複と相互の機能喪失により生じた新たな生殖障壁
高田美信1・鈴木 剛2・渡辺正夫1
(1東北大学大学院生命科学研究科 生態システム生命科学専攻植物生殖遺伝分野,2大阪教育大学 理数情報講座植物分子遺伝学研究室)
email:高田美信,鈴木 剛,渡辺正夫
DOI: 10.7875/first.author.2017.074
Duplicated pollen-pistil recognition loci control intraspecific unilateral incompatibility in Brassica rapa.
Yoshinobu Takada, Kohji Murase, Hiroko Shimosato-Asano, Takahiro Sato, Honoka Nakanishi, Keita Suwabe, Kentaro K. Shimizu, Yong Pyo Lim, Seiji Takayama, Go Suzuki, Masao Watanabe
Nature Plants, 3, 17096 (2017)
植物の有性生殖においては受粉から受精までのさまざまなステップに存在する雌雄の認識機構が解明されつつある.今回,筆者らは,日本とトルコという地理的に離れた地域に由来するアブラナの交配実験から,同一の種であるにもかかわらず受精にいたらない不和合性のあることを見い出し,その制御タンパク質をつきとめた.柱頭の側の制御タンパク質SUI1および花粉の側の制御タンパク質PUI1は,それぞれ,自家不和合性を制御する自他認識タンパク質SRKおよびSP11と相同な受容体型キナーゼおよびそのリガンドであり,この2つのタンパク質が結合することにより不和合性のシグナルが伝達されるものと推察された.このSUI1およびPUI1による一側性不和合性の機構は,SRK遺伝子およびSP11遺伝子がセットで遺伝子重複したのち,日本およびトルコにおいて相互に機能喪失したことにより顕在化したと考えられた.現在は同一の種として扱われている日本とトルコのアブラナが,集団のあいだではすでに潜在的な生殖障壁をもつともいえる.この研究により,植物のゲノムに多く存在するSRK様の遺伝子およびSP11様の遺伝子が種間不和合性における他者の認識に関与する可能性が示された.
植物にとり野外で無作為にはこばれてくる花粉のなかから適切な交雑相手を選択することは,種内の遺伝的な多様性を維持しながらも種の同一性を保つために重要である.そのため,植物は生殖の相手を選別するためのさまざまな機構を発達させてきた1).なかでも,自己の花粉と非自己の花粉を識別し,自己の花粉を拒絶することにより近親交雑を避け,種内の遺伝的な多様性を維持する機構として知られる自家不和合性はさまざまな種にみられ,これまで多くの研究がなされてきた1,2).アブラナ科植物の自家不和合性はひとつの遺伝子座により制御される.このS遺伝子座領域には,柱頭の側のタンパク質である受容体型キナーゼをコードするSRK遺伝子と,花粉の側のタンパク質であるリガンドペプチドをコードするSP11遺伝子がセットで存在する.自己の花粉が受粉した場合,柱頭のSRKと花粉のSP11が結合して花粉の拒絶のシグナルが伝達され不和合反応がひき起こされる1,2).一方で,近縁な他種の花粉を識別する機構である種間不和合性もまた,種の概念の根幹にかかわる重要な形質である.この種間不和合性は,多くの場合,一方向の交雑のみが不和合性となる一側性不和合性として観察され,めしべと花粉の不和合性として古くから知られている1).この一側性不和合性においては,互いに近縁な自家和合性の種と自家不和合性の種のあいだの交雑において,自家和合性の種のめしべに自家不和合性の種の花粉を交配した場合には花粉管がめしべの内部に侵入し伸長するものの,逆の組合せではめしべの柱頭あるいはめしべの内部で花粉管は停止し受精にはいたらないという現象が知られている1,3).また,このルールには例外も報告されている1).一側性不和合性はナス科植物において多く観察されており,種間不和合性の分子機構にナス科植物のもつ協調的な非自己認識モデルによる自家不和合性の機構そのものがかかわることが明らかにされつつある4-7)(文献4) は新着論文レビュー でも掲載).一方,アブラナ科植物においては,受粉の際の一側性不和合性の分子機構について,自家不和合性の分子機構となんらかの関連のあることは示されていたものの3),その実体はまったく解明されていない.
これまでに,筆者らは,アブラナ科植物Brassica rapaを材料とし,日本とトルコという異なる地域に由来する自家不和合性をもつ個体のあいだに新規な一側性不和合性を見い出し,これを遺伝学的に解析してきた.この一側性不和合性は日本に由来する個体の柱頭にトルコに由来する個体の花粉を交配した場合にみられるもので,逆の組合せでは交雑が成立した.分離世代を用いた遺伝学的な解析により,この一側性不和合性は柱頭の側も花粉の側もひとつの遺伝子座により制御され,両者は強く連鎖していること,他者の認識ののちのシグナル伝達の機構には自家不和合性の機構と同じ遺伝子が関与することが示されていた8,9).
ポジショナルクローニングからアブラナ科植物に見い出された新規な種内の一側性不和合性の原因遺伝子の探索を開始した.柱頭の側および花粉の側の原因遺伝子が強く連鎖するという先行研究をうけ9),柱頭の側および花粉の側の形質が分離する世代を用いた遺伝学的な解析の結果,この一側性不和合性を制御するそれぞれの遺伝子をB. rapaのA04染色体の約50 kbの領域に絞り込んだ.この領域には12個の遺伝子が推定されており,それぞれの配列の特徴からBr017016遺伝子に着目した.Br017016遺伝子はめしべの柱頭に特異的に発現するSRK様の遺伝子として,筆者らが2001年に報告していたBcRK6遺伝子と対応していた10).Br017016遺伝子は下流の遺伝子と融合した状態でデータベースに登録されていたが,柱頭の側の一側性不和合性の表現型をもつ個体の柱頭から得たRNAからBr017016遺伝子に対応するcDNAをクローニングし,新たにSUI1遺伝子と名づけた(図1a).SUI1遺伝子はSRK遺伝子と同様の膜貫通型の受容体型キナーゼをコードしており,この一側性不和合性の柱頭の側の原因遺伝子の候補と考えた.
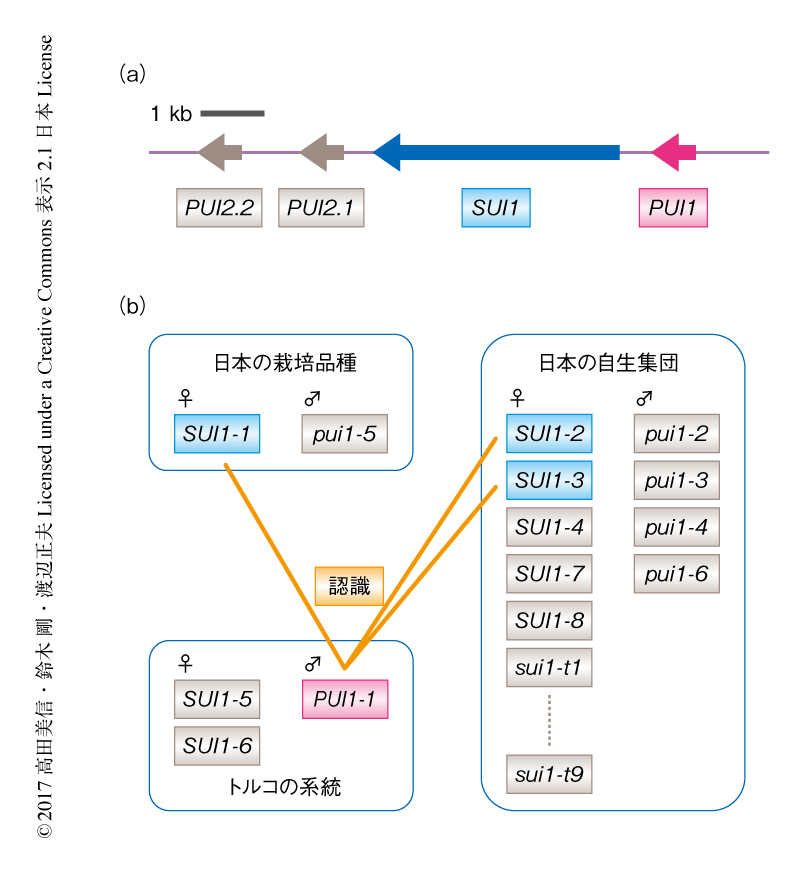
また,Br017016遺伝子の周辺の塩基配列を詳細に解析した結果,SP11遺伝子と類似した配列をもち多数のシステイン残基を含む小型の分泌型のペプチドをコードする複数の遺伝子が見い出された(図1a).そこで,柱頭の側と同様に,花粉の側の一側性不和合性の表現型をもつ個体の葯からcDNAをクローニングし,それぞれ,PUI1遺伝子,PUI2.1遺伝子,PUI2.2遺伝子と名づけた.PUI2.1遺伝子とPUI2.2遺伝子はまったく同じアミノ酸配列をコードしていた.花粉の側の原因遺伝子の候補を絞り込むため,RNAゲルブロット法により発現を解析した結果,PUI1遺伝子はSP11遺伝子と同様に葯に特異的な発現を示し,未成熟の葯においてもっとも高いRNAの蓄積を示した.一方,PUI2.1遺伝子およびPUI2.2遺伝子は転写産物が確認されなかったことから,PUI1遺伝子を花粉の側の原因遺伝子の第1候補とした.
SUI1遺伝子およびPUI1遺伝子について,B. rapaの日本およびトルコに由来する自生集団および栽培品種を用いて遺伝子多型について調べた.SUI1遺伝子については,受容体型キナーゼとしての全長をもつと考えられるSUI1-1からSUI1-8の8つのアリル,フレームシフトによりC末端側の切れた活性をもたないと考えられるsui1-t1からsui1-t9の9つのアリルがクローニングされた(図1b).系統解析の結果,機能型と考えられるSUI1-1アリルの細胞外受容体領域において非常に高い相同性をもつSUI1-2アリルおよびSUI1-3アリルが見い出された.これらのアリルは日本に由来する自生集団から見い出されたものであり,これら植物体を用いた検定交配の結果,SUI1-2アリルあるいはSUI1-3アリルをもつ個体の柱頭は,花粉の側の一側性不和合性をもつ花粉に対し不和合性を示した.つまり,栽培品種のほかにも日本の自生集団にはトルコに由来する花粉を拒絶する一側性不和合性の表現型をもつ個体が存在することが示された.
PUI1遺伝子については,トルコに由来するすべての系統は機能型のPUI1-1アリルをホモにもつ一方,日本に由来するすべての系統は機能型のPUI1-1アリルはもたなかった(図1b).アリルの塩基配列は相互によく類似しており,交配実験から一側性不和合性をひき起こさないことが明らかにされたpui1-2と機能型のPUI1-1の推定アミノ酸配列の違いはわずかに3アミノ酸残基であった.
一側性不和合性の柱頭の側の原因遺伝子がSUI1遺伝子であることを証明するため,遺伝子導入による相補試験を行った.柱頭の側の一側性不和合性の表現型を示さない個体に,柱頭に特異的に発現するSRK遺伝子のプロモーターの制御下においたSUI1遺伝子を導入したところ,3つの形質転換体すべてにおいて導入されたSUI1遺伝子の転写産物の発現が検出された.さらに,これら3つの形質転換体の柱頭にトルコに由来する花粉の側の一側性不和合性の表現型を示す個体の花粉を受粉させたところ,自家不和合性の観点からは非自己の組合せであるにもかかわらず,柱頭において不和合性を示した.一方で,日本に由来する一側性不和合性をもたない花粉はこの形質転換体の柱頭において和合性を示した.このことから,SUI1遺伝子がこの一側性不和合性の柱頭の側の原因遺伝子であることが証明された.
一側性不和合性の花粉の側の原因遺伝子がPUI1遺伝子であることを証明するために,2つの独立した手法を用いた.ひとつは,化学合成したPUI1を用いたバイオアッセイである.化学合成ののち,SP11のジスルフィド結合の状態に関する情報11) をもとにリフォールディングしたPUI1を柱頭の側の一側性不和合性の表現型を示す個体の柱頭の先端の乳頭細胞部に塗布したのち,花粉の側の一側性不和合性の表現型を示さない個体の花粉を受粉させた.その結果,塗布したPUI1の濃度に依存した不和合性がみられた.もうひとつは,SUI1遺伝子と同様な遺伝子導入による相補試験である.花粉の側の一側性不和合性の表現型を示さない個体に,葯のタペート組織に特異的に発現するSP11遺伝子のプロモーターの制御下においたPUI1遺伝子あるいはPUI2.1遺伝子を導入した.その結果,PUI1遺伝子を導入した6つの形質転換体の花粉は柱頭の側の一側性不和合性の表現型を示す個体の柱頭に対し不和合性を示した一方,PUI2.1遺伝子を導入した6つの形質転換体においては花粉の表現型に変化はみられなかった.以上の結果から,PUI1遺伝子がこの一側性不和合性の花粉の側の原因遺伝子であることが証明された.
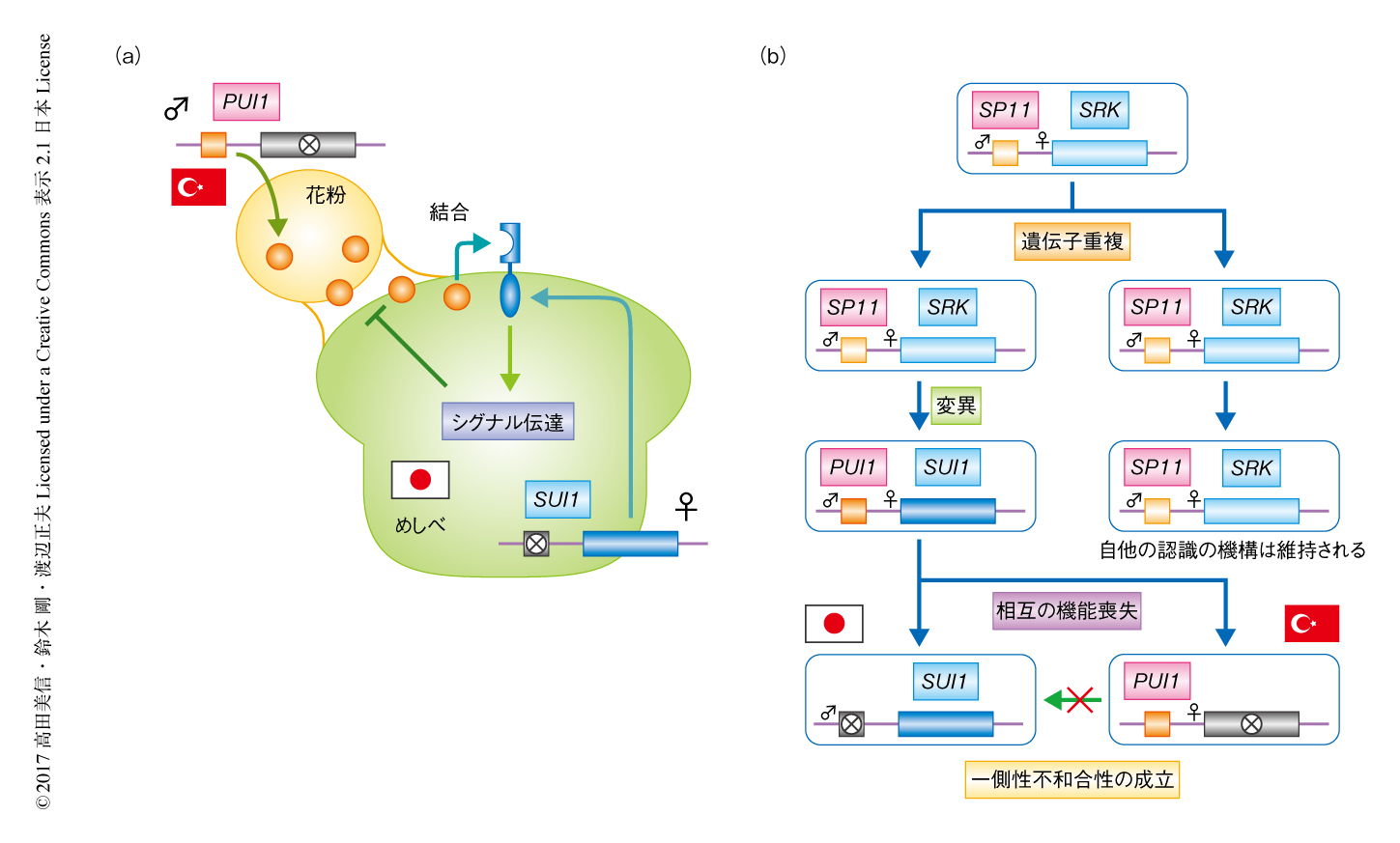
柱頭の側のSUI1遺伝子および花粉の側のPUI1遺伝子は,自家不和合性の機構におけるSRK遺伝子およびSP11遺伝子と相同な受容体型キナーゼおよびそのリガンドペプチドをコードしていた.すなわち,花粉に由来するPUI1と乳頭細胞のSUI1とが結合し不和合性のシグナルを伝達することにより花粉の拒絶にいたる機構が考えられた(図2a).また,相同性および系統解析の結果から,SUI1遺伝子およびPUI1遺伝子はSRK遺伝子およびSP11遺伝子の存在するS遺伝子座領域における遺伝子重複に起源すると考えられた.それぞれの遺伝子のアリルの分布の結果をあわせて考えると,遺伝子重複を起こしたSRK遺伝子およびSP11遺伝子がトルコと日本という離れた地域において相互に機能を喪失することにより,この新たな生殖障壁が生じたものと推察された(図2b).また,日本およびトルコのそれぞれの地域においては,花粉の側のタンパク質あるいは柱頭の側のタンパク質を欠失しているため一側性不和合性は起こらないものと考えられた.
ゲノム情報の公開されているB. rapaの近縁種であるBrassica oleraceaを対象としてゲノム比較解析をしたところ,B. oleraceaにおいては対応する領域はみつかったものの,SUI1遺伝子にはフレームシフトによる変異がみられたことから機能を失っているものと考えられた.また,PUI2遺伝子は存在したものの,PUI1遺伝子の同祖遺伝子は欠失していた.Arabidopsis属にはSUI1-PUI1領域それ自体が存在しなかった.これらの結果から,SUI1遺伝子およびPUI1遺伝子の誕生にかかわるS遺伝子座領域における遺伝子重複は,Arabidopsis属とBrassica属が分化したのちに起こったと考えられた.
この研究において解析した一側性不和合性は,種内で起こること,両親ともに自家不和合性であること,の2点において,種間で起こる一側性不和合性とは異なる.つまり,今回,明らかにされたSUI1およびPUI1による一側性不和合性により,アブラナ科植物における種間の不和合性の機構が解明されたわけではない.しかしながら,自家不和合性における自他認識遺伝子であるSRK遺伝子およびSP11遺伝子はそれぞれ大きなファミリーを形成しており10),なかにはSRK様の遺伝子とSP11様の遺伝子がこの研究の場合と同様にゲノムにおいて近接して存在する場合のあること12),および,自家不和合性と種間の一側性不和合性とのあいだには関連性のあることを考えると1,3),S遺伝子座領域の遺伝子重複,多型の獲得,機能喪失が種間の生殖障壁にどのように関連するか興味深い.
筆者らは,これまでまったく知られていなかったアブラナ科植物における一側性不和合性の解明をつうじて,他者認識による不和合性の機構の一端を明らかにした.この研究が植物の生殖機構の研究において新たな展開をもたらすことを期待する.
略歴:2005年 岩手大学大学院連合農学研究科 修了,同年 大阪教育大学 研究員,東北大学大学院生命科学研究科 研究員を経て,2007年より同 技術職員.
研究テーマ:アブラナ科植物における受粉の際の不和合性の認識の機構および花粉の拒絶の機構.
抱負:菜の花の受粉のときに起こる認識,拒絶,許容の機構のほんの少しをのぞきみたい.
鈴木 剛(Go Suzuki)
大阪教育大学 教授.
研究室URL:http://web.nsc.osaka-kyoiku.ac.jp/life/suzuki/suzuki.html
渡辺 正夫(Masao Watanabe)
東北大学大学院生命科学研究科 教授.
研究室URL:http://www.ige.tohoku.ac.jp/prg/watanabe/
? 2017 高田美信・鈴木 剛・渡辺正夫 Licensed under CC 表示 2.1 日本
(1東北大学大学院生命科学研究科 生態システム生命科学専攻植物生殖遺伝分野,2大阪教育大学 理数情報講座植物分子遺伝学研究室)
email:高田美信,鈴木 剛,渡辺正夫
DOI: 10.7875/first.author.2017.074
Duplicated pollen-pistil recognition loci control intraspecific unilateral incompatibility in Brassica rapa.
Yoshinobu Takada, Kohji Murase, Hiroko Shimosato-Asano, Takahiro Sato, Honoka Nakanishi, Keita Suwabe, Kentaro K. Shimizu, Yong Pyo Lim, Seiji Takayama, Go Suzuki, Masao Watanabe
Nature Plants, 3, 17096 (2017)
要 約
植物の有性生殖においては受粉から受精までのさまざまなステップに存在する雌雄の認識機構が解明されつつある.今回,筆者らは,日本とトルコという地理的に離れた地域に由来するアブラナの交配実験から,同一の種であるにもかかわらず受精にいたらない不和合性のあることを見い出し,その制御タンパク質をつきとめた.柱頭の側の制御タンパク質SUI1および花粉の側の制御タンパク質PUI1は,それぞれ,自家不和合性を制御する自他認識タンパク質SRKおよびSP11と相同な受容体型キナーゼおよびそのリガンドであり,この2つのタンパク質が結合することにより不和合性のシグナルが伝達されるものと推察された.このSUI1およびPUI1による一側性不和合性の機構は,SRK遺伝子およびSP11遺伝子がセットで遺伝子重複したのち,日本およびトルコにおいて相互に機能喪失したことにより顕在化したと考えられた.現在は同一の種として扱われている日本とトルコのアブラナが,集団のあいだではすでに潜在的な生殖障壁をもつともいえる.この研究により,植物のゲノムに多く存在するSRK様の遺伝子およびSP11様の遺伝子が種間不和合性における他者の認識に関与する可能性が示された.
はじめに
植物にとり野外で無作為にはこばれてくる花粉のなかから適切な交雑相手を選択することは,種内の遺伝的な多様性を維持しながらも種の同一性を保つために重要である.そのため,植物は生殖の相手を選別するためのさまざまな機構を発達させてきた1).なかでも,自己の花粉と非自己の花粉を識別し,自己の花粉を拒絶することにより近親交雑を避け,種内の遺伝的な多様性を維持する機構として知られる自家不和合性はさまざまな種にみられ,これまで多くの研究がなされてきた1,2).アブラナ科植物の自家不和合性はひとつの遺伝子座により制御される.このS遺伝子座領域には,柱頭の側のタンパク質である受容体型キナーゼをコードするSRK遺伝子と,花粉の側のタンパク質であるリガンドペプチドをコードするSP11遺伝子がセットで存在する.自己の花粉が受粉した場合,柱頭のSRKと花粉のSP11が結合して花粉の拒絶のシグナルが伝達され不和合反応がひき起こされる1,2).一方で,近縁な他種の花粉を識別する機構である種間不和合性もまた,種の概念の根幹にかかわる重要な形質である.この種間不和合性は,多くの場合,一方向の交雑のみが不和合性となる一側性不和合性として観察され,めしべと花粉の不和合性として古くから知られている1).この一側性不和合性においては,互いに近縁な自家和合性の種と自家不和合性の種のあいだの交雑において,自家和合性の種のめしべに自家不和合性の種の花粉を交配した場合には花粉管がめしべの内部に侵入し伸長するものの,逆の組合せではめしべの柱頭あるいはめしべの内部で花粉管は停止し受精にはいたらないという現象が知られている1,3).また,このルールには例外も報告されている1).一側性不和合性はナス科植物において多く観察されており,種間不和合性の分子機構にナス科植物のもつ協調的な非自己認識モデルによる自家不和合性の機構そのものがかかわることが明らかにされつつある4-7)(文献4) は新着論文レビュー でも掲載).一方,アブラナ科植物においては,受粉の際の一側性不和合性の分子機構について,自家不和合性の分子機構となんらかの関連のあることは示されていたものの3),その実体はまったく解明されていない.
これまでに,筆者らは,アブラナ科植物Brassica rapaを材料とし,日本とトルコという異なる地域に由来する自家不和合性をもつ個体のあいだに新規な一側性不和合性を見い出し,これを遺伝学的に解析してきた.この一側性不和合性は日本に由来する個体の柱頭にトルコに由来する個体の花粉を交配した場合にみられるもので,逆の組合せでは交雑が成立した.分離世代を用いた遺伝学的な解析により,この一側性不和合性は柱頭の側も花粉の側もひとつの遺伝子座により制御され,両者は強く連鎖していること,他者の認識ののちのシグナル伝達の機構には自家不和合性の機構と同じ遺伝子が関与することが示されていた8,9).
1.新規の一側性不和合性の柱頭の側および花粉の側における原因遺伝子の探索
ポジショナルクローニングからアブラナ科植物に見い出された新規な種内の一側性不和合性の原因遺伝子の探索を開始した.柱頭の側および花粉の側の原因遺伝子が強く連鎖するという先行研究をうけ9),柱頭の側および花粉の側の形質が分離する世代を用いた遺伝学的な解析の結果,この一側性不和合性を制御するそれぞれの遺伝子をB. rapaのA04染色体の約50 kbの領域に絞り込んだ.この領域には12個の遺伝子が推定されており,それぞれの配列の特徴からBr017016遺伝子に着目した.Br017016遺伝子はめしべの柱頭に特異的に発現するSRK様の遺伝子として,筆者らが2001年に報告していたBcRK6遺伝子と対応していた10).Br017016遺伝子は下流の遺伝子と融合した状態でデータベースに登録されていたが,柱頭の側の一側性不和合性の表現型をもつ個体の柱頭から得たRNAからBr017016遺伝子に対応するcDNAをクローニングし,新たにSUI1遺伝子と名づけた(図1a).SUI1遺伝子はSRK遺伝子と同様の膜貫通型の受容体型キナーゼをコードしており,この一側性不和合性の柱頭の側の原因遺伝子の候補と考えた.
また,Br017016遺伝子の周辺の塩基配列を詳細に解析した結果,SP11遺伝子と類似した配列をもち多数のシステイン残基を含む小型の分泌型のペプチドをコードする複数の遺伝子が見い出された(図1a).そこで,柱頭の側と同様に,花粉の側の一側性不和合性の表現型をもつ個体の葯からcDNAをクローニングし,それぞれ,PUI1遺伝子,PUI2.1遺伝子,PUI2.2遺伝子と名づけた.PUI2.1遺伝子とPUI2.2遺伝子はまったく同じアミノ酸配列をコードしていた.花粉の側の原因遺伝子の候補を絞り込むため,RNAゲルブロット法により発現を解析した結果,PUI1遺伝子はSP11遺伝子と同様に葯に特異的な発現を示し,未成熟の葯においてもっとも高いRNAの蓄積を示した.一方,PUI2.1遺伝子およびPUI2.2遺伝子は転写産物が確認されなかったことから,PUI1遺伝子を花粉の側の原因遺伝子の第1候補とした.
2.SUI1遺伝子およびPUI1遺伝子のアリルの分布
SUI1遺伝子およびPUI1遺伝子について,B. rapaの日本およびトルコに由来する自生集団および栽培品種を用いて遺伝子多型について調べた.SUI1遺伝子については,受容体型キナーゼとしての全長をもつと考えられるSUI1-1からSUI1-8の8つのアリル,フレームシフトによりC末端側の切れた活性をもたないと考えられるsui1-t1からsui1-t9の9つのアリルがクローニングされた(図1b).系統解析の結果,機能型と考えられるSUI1-1アリルの細胞外受容体領域において非常に高い相同性をもつSUI1-2アリルおよびSUI1-3アリルが見い出された.これらのアリルは日本に由来する自生集団から見い出されたものであり,これら植物体を用いた検定交配の結果,SUI1-2アリルあるいはSUI1-3アリルをもつ個体の柱頭は,花粉の側の一側性不和合性をもつ花粉に対し不和合性を示した.つまり,栽培品種のほかにも日本の自生集団にはトルコに由来する花粉を拒絶する一側性不和合性の表現型をもつ個体が存在することが示された.
PUI1遺伝子については,トルコに由来するすべての系統は機能型のPUI1-1アリルをホモにもつ一方,日本に由来するすべての系統は機能型のPUI1-1アリルはもたなかった(図1b).アリルの塩基配列は相互によく類似しており,交配実験から一側性不和合性をひき起こさないことが明らかにされたpui1-2と機能型のPUI1-1の推定アミノ酸配列の違いはわずかに3アミノ酸残基であった.
3.SUI1遺伝子は一側性不和合性の柱頭の側の原因遺伝子である
一側性不和合性の柱頭の側の原因遺伝子がSUI1遺伝子であることを証明するため,遺伝子導入による相補試験を行った.柱頭の側の一側性不和合性の表現型を示さない個体に,柱頭に特異的に発現するSRK遺伝子のプロモーターの制御下においたSUI1遺伝子を導入したところ,3つの形質転換体すべてにおいて導入されたSUI1遺伝子の転写産物の発現が検出された.さらに,これら3つの形質転換体の柱頭にトルコに由来する花粉の側の一側性不和合性の表現型を示す個体の花粉を受粉させたところ,自家不和合性の観点からは非自己の組合せであるにもかかわらず,柱頭において不和合性を示した.一方で,日本に由来する一側性不和合性をもたない花粉はこの形質転換体の柱頭において和合性を示した.このことから,SUI1遺伝子がこの一側性不和合性の柱頭の側の原因遺伝子であることが証明された.
4.PUI1遺伝子は一側性不和合性の花粉の側の原因遺伝子である
一側性不和合性の花粉の側の原因遺伝子がPUI1遺伝子であることを証明するために,2つの独立した手法を用いた.ひとつは,化学合成したPUI1を用いたバイオアッセイである.化学合成ののち,SP11のジスルフィド結合の状態に関する情報11) をもとにリフォールディングしたPUI1を柱頭の側の一側性不和合性の表現型を示す個体の柱頭の先端の乳頭細胞部に塗布したのち,花粉の側の一側性不和合性の表現型を示さない個体の花粉を受粉させた.その結果,塗布したPUI1の濃度に依存した不和合性がみられた.もうひとつは,SUI1遺伝子と同様な遺伝子導入による相補試験である.花粉の側の一側性不和合性の表現型を示さない個体に,葯のタペート組織に特異的に発現するSP11遺伝子のプロモーターの制御下においたPUI1遺伝子あるいはPUI2.1遺伝子を導入した.その結果,PUI1遺伝子を導入した6つの形質転換体の花粉は柱頭の側の一側性不和合性の表現型を示す個体の柱頭に対し不和合性を示した一方,PUI2.1遺伝子を導入した6つの形質転換体においては花粉の表現型に変化はみられなかった.以上の結果から,PUI1遺伝子がこの一側性不和合性の花粉の側の原因遺伝子であることが証明された.
5.SUI1およびPUI1による一側性不和合性の機構とその進化
柱頭の側のSUI1遺伝子および花粉の側のPUI1遺伝子は,自家不和合性の機構におけるSRK遺伝子およびSP11遺伝子と相同な受容体型キナーゼおよびそのリガンドペプチドをコードしていた.すなわち,花粉に由来するPUI1と乳頭細胞のSUI1とが結合し不和合性のシグナルを伝達することにより花粉の拒絶にいたる機構が考えられた(図2a).また,相同性および系統解析の結果から,SUI1遺伝子およびPUI1遺伝子はSRK遺伝子およびSP11遺伝子の存在するS遺伝子座領域における遺伝子重複に起源すると考えられた.それぞれの遺伝子のアリルの分布の結果をあわせて考えると,遺伝子重複を起こしたSRK遺伝子およびSP11遺伝子がトルコと日本という離れた地域において相互に機能を喪失することにより,この新たな生殖障壁が生じたものと推察された(図2b).また,日本およびトルコのそれぞれの地域においては,花粉の側のタンパク質あるいは柱頭の側のタンパク質を欠失しているため一側性不和合性は起こらないものと考えられた.
ゲノム情報の公開されているB. rapaの近縁種であるBrassica oleraceaを対象としてゲノム比較解析をしたところ,B. oleraceaにおいては対応する領域はみつかったものの,SUI1遺伝子にはフレームシフトによる変異がみられたことから機能を失っているものと考えられた.また,PUI2遺伝子は存在したものの,PUI1遺伝子の同祖遺伝子は欠失していた.Arabidopsis属にはSUI1-PUI1領域それ自体が存在しなかった.これらの結果から,SUI1遺伝子およびPUI1遺伝子の誕生にかかわるS遺伝子座領域における遺伝子重複は,Arabidopsis属とBrassica属が分化したのちに起こったと考えられた.
おわりに
この研究において解析した一側性不和合性は,種内で起こること,両親ともに自家不和合性であること,の2点において,種間で起こる一側性不和合性とは異なる.つまり,今回,明らかにされたSUI1およびPUI1による一側性不和合性により,アブラナ科植物における種間の不和合性の機構が解明されたわけではない.しかしながら,自家不和合性における自他認識遺伝子であるSRK遺伝子およびSP11遺伝子はそれぞれ大きなファミリーを形成しており10),なかにはSRK様の遺伝子とSP11様の遺伝子がこの研究の場合と同様にゲノムにおいて近接して存在する場合のあること12),および,自家不和合性と種間の一側性不和合性とのあいだには関連性のあることを考えると1,3),S遺伝子座領域の遺伝子重複,多型の獲得,機能喪失が種間の生殖障壁にどのように関連するか興味深い.
筆者らは,これまでまったく知られていなかったアブラナ科植物における一側性不和合性の解明をつうじて,他者認識による不和合性の機構の一端を明らかにした.この研究が植物の生殖機構の研究において新たな展開をもたらすことを期待する.
文 献
- de Nettancourt, D.: Incompatibility and Incongruity in Wild and Cultivated Plants. Springer-Verlag, Berlin (2001)
- Watanabe, M., Suwabe, K. & Suzuki, G.: Molecular genetics, physiology and biology of self-incompatibility in Brassicaceae. Proc. Jpn. Acad. Ser. B, 88, 519-535 (2012)[PubMed]
- Hiscock, S. J. & Dickinson, H. G.: Unilateral incompatibility within the Brassicaceae: further evidence for the involvement of the self-incompatibility (S)-locus. Theor. Appl. Genet., 86, 744-753 (1993)[PubMed]
- Kubo, K., Entani, E. & Takara, A.: Collaborative non-self recognition system in S-RNase-based self-incompatibility. Science, 330, 796-799 (2010)[PubMed] [新着論文レビュー]
- Kubo, K., Paape, T., Hatakeyama, M. et al.: Gene duplication and genetic exchange drive the evolution of S-RNase-based self-incompatibility in Petunia. Nature Plants, 1, 14005 (2015)[PubMed]
- Li, W. & Chetelat, R. T.: Unilateral incompatibility gene ui1.1 encodes an S-locus F-box protein expressed in pollen of Solanum species. Proc. Natl Acad. Sci. USA, 112, 4417-4422 (2015)[PubMed]
- Fujii, S., Kubo, K. & Takayama, S.: Non-self- and self-recognition models in plant self-incompatibility. Nature Plants, 2, 16130 (2016)[PubMed]
- Takada, Y., Nakanowatari, T., Sato, J. et al.: Genetic analysis of novel intra-species unilateral incompatibility in Brassica rapa (syn. campestris) L. Sex. Plant Reprod., 17, 211-217 (2005)
- Takada, Y., Sato, T., Suzuki, G. et al.: Involvement of MLPK pathway in intraspecies unilateral incompatibility regulated by a single locus with stigma and pollen factors. G3, 3, 719-726 (2013)[PubMed]
- Kai, N., Suzuki, G., Watanabe, M. et al.: Sequence comparisons among dispersed members of the Brassica S multigene family in an S9 genome. Mol. Genet. Genomics, 265, 526-534 (2001)[PubMed]
- Takayama, S., Shimosato, H., Shiba, H. et al.: Direct ligand-receptor complex interaction controls Brassica self-incompatibility. Nature, 413, 534-538 (2001)[PubMed]
- Zhang, X., Wang, L., Yuan, Y. et al.: Rapid copy number expansion and recent recruitment of domains in S-receptor kinase-like genes contribute to the origin of self-incompatibility. FEBS J., 278, 4323-4337 (2011)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2005年 岩手大学大学院連合農学研究科 修了,同年 大阪教育大学 研究員,東北大学大学院生命科学研究科 研究員を経て,2007年より同 技術職員.
研究テーマ:アブラナ科植物における受粉の際の不和合性の認識の機構および花粉の拒絶の機構.
抱負:菜の花の受粉のときに起こる認識,拒絶,許容の機構のほんの少しをのぞきみたい.
鈴木 剛(Go Suzuki)
大阪教育大学 教授.
研究室URL:http://web.nsc.osaka-kyoiku.ac.jp/life/suzuki/suzuki.html
渡辺 正夫(Masao Watanabe)
東北大学大学院生命科学研究科 教授.
研究室URL:http://www.ige.tohoku.ac.jp/prg/watanabe/
? 2017 高田美信・鈴木 剛・渡辺正夫 Licensed under CC 表示 2.1 日本
