S-RNase型の自家不和合性における協調的な非自己認識系
久保健一・円谷徹之・高山誠司
(奈良先端科学技術大学院大学バイオサイエンス研究科 細胞生物学専攻細胞間情報学講座)
email:久保健一,高山誠司
DOI: 10.7875/first.author.2010.059
Collaborative non-self recognition system in S-RNase-based self-incompatibility.
Ken-ichi Kubo, Tetsuyuki Entani, Akie Takara, Ning Wang, Allison M. Fields, Zhihua Hua, Mamiko Toyoda, Shin-ichi Kawashima, Toshio Ando, Akira Isogai, Teh-hui Kao, Seiji Takayama
Science, 330, 796-799 (2010)
顕花植物の自家不和合性は自殖をさまたげ他殖を促すことによって遺伝的な多様性を保つ遺伝的な機構である.ナス科植物ではS遺伝子座にコードされるFボックスタンパク質SLFが自家不和合性の花粉側因子である.SLFの対立遺伝子産物は雌しべ側因子であるS-RNaseの非自己の対立遺伝子産物を特異的に解毒することによって和合性受粉を可能にしているものと考えられてきた.しかしながら,SLFは対立遺伝子産物のあいだでの配列多型性がきわめて低く,どのようにして非自己のS-RNaseの膨大なレパートリーすべてを認識しているのか謎であった.筆者らは,in vivo機能アッセイおよびタンパク質相互作用アッセイにより,ナス科植物のペチュニアでは少なくとも3つのタイプの異なる遺伝子にコードされたSLFが花粉側因子として協調的に機能し,それぞれが一部の非自己のS-RNaseを分担して認識していることを明らかにし,S-RNase型の自家不和合性機構として協調的な非自己認識系を提唱した.
自家不和合性は顕花植物の種内生殖障壁であり,雌しべにおける自己(遺伝的に近縁の)花粉の排除および非自己(遺伝的に無関係の)花粉の受容を可能にしている.この自己と非自己との識別は,単一のS遺伝子座の複対立遺伝子にコードされた花粉側因子と雌しべ側因子とによって調節されている1,2)(図1).花粉側因子と雌しべ側因子をコードする遺伝子は密接に連鎖して組み換わることなく遺伝するため,これら遺伝子のセットはSハプロタイプとよばれている.ナス科,バラ科,オオバコ科の自家不和合性を示す種は雌しべ側因子として分泌型リボヌクレアーゼであるS-RNaseを用いている1,3).半数体花粉のSハプロタイプが花柱のSハプロタイプのいずれかと一致した場合,S-RNaseは自己の花粉管の内部で細胞毒性を発揮してその伸長を阻害する1,3).一方,花粉側因子はSLF(S-locus F-box)と名づけられたFボックスタンパク質であり3,4),これらはSCF(Skp1-Cullin1-F-box)型ユビキチンリガーゼ複合体の構成タンパク質であることが示唆されてきた3,4).
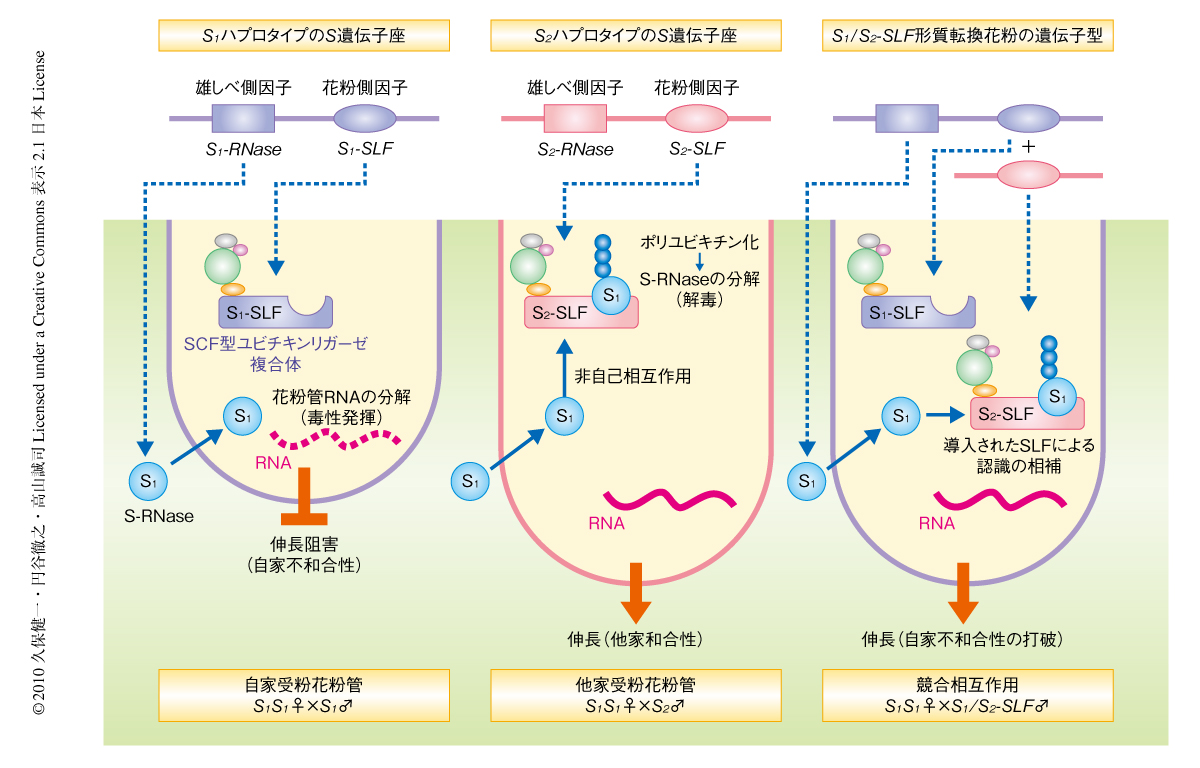
S-RNaseによる花粉管のSハプロタイプの特異的な排除を説明するためタンパク質分解モデルが提唱されてきた(図1).このモデルでは,SLFが自己以外のすべての非自己のS-RNaseを認識し,ユビキチン-プロテアソーム系を介してこれを分解し解毒することを予想する1,3,4).このモデルは古くから知られる4倍体植物が自家和合性を示す“競合相互作用”とよばれる現象,すなわち,2つの異なる花粉側因子の対立遺伝子をもつ花粉が自家和合性を示す現象を矛盾なく説明することができる5,6)(図1).2つの対立遺伝子をもつ花粉から生じる2つの異なるSLFは互いに認識特異性を相補しあうことによってすべてのS-RNaseを分解することが可能となり,結果として,花粉管はすべてのSハプロタイプの花柱に対して和合性になると説明しうる3).実際,競合相互作用にもとづいてデザインされたin vivo機能アッセイにより,ナス科のペチュニアの一種Petunia inflataのS2ハプロタイプに由来するSLFが花粉側因子として機能することが証明された.すなわち,S2ハプロタイプに由来するSLF対立遺伝子をS1S2個体およびS2S3個体に導入する実験が行われ,その結果,異なる対立遺伝子をもつS1花粉あるいはS3花粉にS2ハプロタイプに由来するSLF対立遺伝子が導入されると自家不和合性は打破されるが,異なる対立遺伝子をもたないS2花粉の場合には自家不和合性が維持されることが示された7).
この実験結果がもととなりSLFが唯一の花粉側因子であると考えられるようになった7,8).しかし,のちの実験により,SLFはその対立遺伝子産物のあいだでの多型性がS-RNaseに比べずっと低く,また,アブラナ科植物の花粉側因子と雌しべ側因子とのあいだで認められた共進化の関係もまったく認められないことが示唆されてきた.さらに,それぞれの種が数十に及ぶSハプロタイプをもつことを考えると,単一のSLFがはたして自己以外のすべてのSハプロタイプに由来するS-RNaseのレパートリーを認識しうるかどうか疑問視されるようになってきた.そこで,この研究では,ペチュニアにおいてすでに同定されたSLFが本当に自家不和合性における唯一の花粉側因子であるのかどうかをまず検証することにした.
ペチュニアのS5,S7,S9,S11のホモ接合体の花粉のcDNAから,既知のSLF対立遺伝子との配列相同性をもとに4つの新規な対立遺伝子をクローニングした.同定されたすべての対立遺伝子から推定されたSLFのアミノ酸配列はきわめて高い相同性を示し(アミノ酸同一性は86.4%~100%),相当するS-RNaseの示す低い相同性(多型性)とは対照的であった(アミノ酸同一性は40.1%~79.4%).なお,S-RNase型の自家不和合性をもつ種は一般に複数のSLF様の遺伝子をもつことが示されてきたことから8-10),混乱をさけるため,S2ハプロタイプに由来するSLF対立遺伝子と高い相同性を示すこれらのSLF対立遺伝子をタイプ1 SLFと名づけ,それぞれの対立遺伝子をSn-SLF1とよぶことにした(nはSハプロタイプの種類を表す).
驚くべきことに,S7-SLF1から推定されたアミノ酸配列はS19-SLF1 6) のものと100%同一であった.S7ハプロタイプおよびS19ハプロタイプは異なるS-RNaseをもち(アミノ酸同一性は45%),それらのホモ接合体のあいだの相互交配から異なる自他認識特異性を示すことも確認された.2つの異なるSハプロタイプにおいて同一のSLF対立遺伝子が発見されたことから,SLFが唯一の花粉側因子ではない可能性が示唆された.
この可能性を調べるため,タイプ1 SLFのそれぞれの対立遺伝子の産物が花粉側因子として機能するかどうかを形質転換実験により検証することにした.たとえば,S7-SLF1(= S19-SLF1)を用いた実験では,自己SハプロタイプをもつS7花粉およびS19花粉だけでなく,非自己SハプロタイプをもつS5花粉およびS11花粉においても競合相互作用が起こらないことが示された.一方,S7-SLF1をさらに別の非自己SハプロタイプをもつS9およびS17花粉において発現させた場合は競合相互作用が観察された.以上の結果は,S7-SLF1が一部の非自己Sハプロタイプをもつ花粉においてのみ選択的な競合相互作用を起こすことを示唆した.タイプ1 SLFのほかの対立遺伝子を用いた形質転換実験においても,同様に一部の非自己Sハプロタイプの花粉においてのみ競合相互作用が観察された.このことは,タンパク質分解モデルにもとづけば,おのおののタイプ1 SLFは非自己S-RNaseの一部を認識しそれらの分解を誘導していることを示唆していた.では,タイプ1 SLFにより認識されなかったほかの非自己S-RNaseは,S遺伝子座にコードされたいずれの因子によって認識されているのだろうか.
ペチュニアではいくつかのSLF様の遺伝子がS遺伝子座と強く連鎖していることが知られてきた8).そこで,このようなSLF様遺伝子にコードされた因子も花粉側因子として機能する可能性を推察した.この仮説を検証するため,既知のSLF様遺伝子の配列情報にもとづき,S5,S7,S9,S11,S17,S19それぞれのハプロタイプの花粉から新規のSLF様遺伝子のcDNA断片を徹底的にクローニングした.
結果として,30個のSLF様遺伝子を同定し,配列類似性にもとづいて5つのサブグループに分類しタイプ2 SLF~タイプ6 SLFと名づけた.タイプ1 SLFと同じ命名法を用いてそれらの対立遺伝子をSn-SLFxとよぶことにした(xはタイプの番号を表す).タイプ1 SLFの場合と同様に,いずれのタイプのSLFもSハプロタイプのあいだでの多型性は低いことが示された(アミノ酸同一性は70%以上).
6つのタイプのSLF遺伝子のすべてが類似した雄性器官に特異的な発現プロファイルを示した.すなわち,葯の発達過程において発現が増加し,開花前にピークに達し,成熟花粉および花粉管において減少した.また,それぞれのSLF遺伝子とS-RNase遺伝子とのあいだの連鎖を調べたところ,6つのタイプのSLF遺伝子のすべてがS-RNase遺伝子の1 cM以内に位置していることが示唆された.以上の結果は,新規の5つのタイプのSLF遺伝子が花粉側因子の遺伝子としての条件を満たすことを示した.
そこでつぎに,タイプ2およびタイプ3に属する5つのSLF対立遺伝子について,形質転換法によるin vivo機能アッセイを行った.その結果,S7-SLF3を除くすべてについて,タイプ1 SLFと同様に調べたいずれかの非自己Sハプロタイプをもつ花粉において競合相互作用が観察された.たとえば,S7-SLF2を用いた実験では,自己のS7ハプロタイプおよび非自己であるS5ハプロタイプあるいは非自己S17ハプロタイプをもつ花粉では競合相互作用はみられなかったが,非自己であるS9ハプロタイプ,S11ハプロタイプ,S19ハプロタイプをもつ花粉では競合相互作用が観察された.なお,今回のの実験で遺伝子導入したS7ハプロタイプおよびS11ハプロタイプに由来する3つのタイプのSLF1~SLF3遺伝子ではいずれもS5花粉においては競合相互作用が観察されず,この事実は,ほかの未解析のタイプのSLF遺伝子の産物がS5ハプロタイプに由来するS5-RNaseの解毒に関与していることを示唆した.さらなる解析は必要であるが,今回の形質転換実験は少なくとも3つ以上のタイプのSLFが花粉側因子として機能していることを示唆した.
複数のタイプのSLFが花粉側因子として機能していると仮定すると,タンパク質分解モデルは以下のように修正される.たとえば,S7花粉が自己のS7花柱に対して不和合性であるのはS7花粉において生産される複数のタイプのSLFのすべてがS7ハプロタイプに由来するS7-RNaseを認識しないためである.また,S7-SLF2を遺伝子導入したS5花粉あるいはS17花粉が競合相互作用を示さなかったのに対し,S9花粉,S11花粉,S19花粉は競合相互作用を示し自己の花柱に対して和合性を示すようになったというさきに述べた実験結果は,S7-SLF2がS5-RNaseあるいはS17-RNaseを認識しないのに対し,S9-RNase,S11-RNase,S19-RNaseを認識して分解するためと理解される.そこで,このモデルを検証するため,つぎにS7-SLF2を用いたタンパク質相互作用解析を行った.
まず,予備実験として,FLAGタグ配列を付加したS7-SLF2をS5S11個体の花粉に発現させ,それがS7-SLF2と同様にS11花粉での競合相互作用を誘導することを観察し,FLAGタグがS7-SLF2の機能に影響しないことを確認した.つぎに,FLAGタグを付加したS7-SLF2を含む形質転換花粉の抽出物を種々の異なるSハプロタイプの花柱抽出物と混合し,抗FLAG抗体を用いて免疫沈降した.その結果,S7-SLF2は予想どおり自己のS7-RNaseや競合相互作用を示さないS5ハプロタイプのS5-RNaseとは相互作用せず,競合相互作用を示すS9ハプロタイプおよびS11ハプロタイプに由来するS9-RNaseおよびS11-RNaseとは強く相互作用することが確認された.これらの結果は,SLFが特定の一部の非自己のS-RNaseと特異的に相互作用し,それらを解毒するというモデルをさらに強く示唆した.
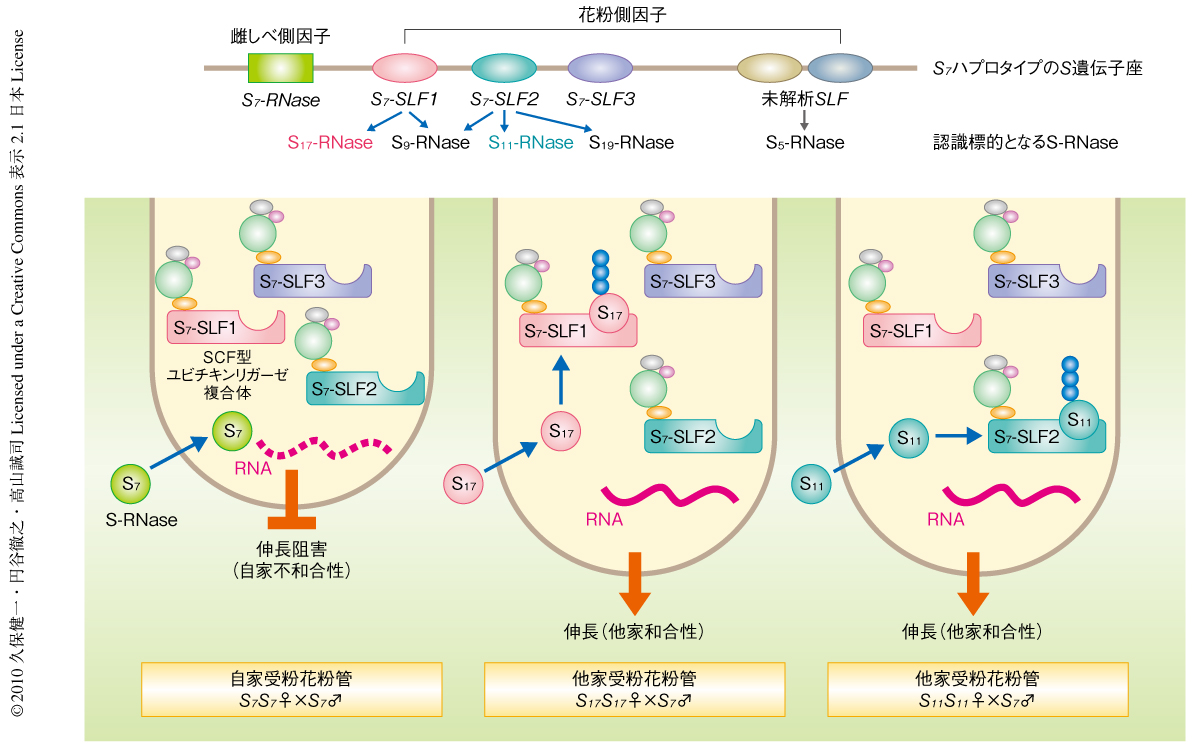
この研究にもとづきS-RNase型の自家不和合性に関して“協調的非自己認識モデル”を提唱した(図2).ペチュニアでは花粉側因子は複数のタイプのSLF遺伝子にコードされている.ひとつのSハプロタイプからつくられるおのおののタイプのSLFが相互作用できる非自己のS-RNaseは一部に限られるが,複数のタイプのSLF遺伝子の産物が協調的に機能することで非自己のS-RNaseのすべてを認識して分解し他家受粉を可能にしている,というモデルである.これまでに明らかにされてきたアブラナ科やケシ科における自家不和合性系ではいずれも雌しべ側因子と花粉側因子が単一の遺伝子によってコードされており,しかも,同一のSハプロタイプに由来する雌しべ側因子と花粉側因子との相互作用を介して“自己”認識が行われてその受精が阻害されていた.今回,解析したペチュニア(ナス科)の自家不和合性系では多数の花粉側因子を用いて異なるSハプロタイプに由来する“非自己”の雌しべ側因子の認識が行われており,根本からまったく異なる自他識別系であることが明らかになった1,11).
この研究から導かれた協調的な非自己モデルは,S-RNase型の自家不和合性の研究において長年の謎であった多型性の低いSLF対立遺伝子によりきわめて多様な非自己のS-RNaseを識別するしくみについてはじめて明確な説明をあたえた.オオバコ科やバラ科の自家不和合性を示す種も単一のS-RNaseの遺伝子と複数のSLF様のFボックスタンパク質の遺伝子をS遺伝子座にもつことが示されている9,10).たとえば,バラ科のリンゴ(Malus×domestica)では12個のSLF様のFボックスタンパク質の遺伝子がS遺伝子座に連鎖している10).ナス科植物とは系統学的に離れたバラ科植物においても協調的な非自己認識モデルが適用できるかどうか,今後,明らかにしていく必要がある.また,複数のタイプのSLF遺伝子がどのように出現し,単一のS-RNase遺伝子とともにどのように進化してきたのか,また,どのようにしてそれぞれのタイプのSLFが一部の非自己のS-RNaseを分子レベルで認識しているのか,さらには,SLFにより認識された非自己のS-RNaseはどのようにして無毒化されているのか,などの課題も,今後,明らかにしていく必要がある.
略歴:2000年 東北大学大学院工学研究科博士課程 修了(工学博士),同年 農業生物資源研究所 博士研究員を経て,2005年より奈良先端科学技術大学院大学バイオサイエンス研究科 博士研究員.
研究テーマ:ナス科植物ペチュニアにおける自家不和合性の分子機構の解明.
関心事:この研究で示した協調的な非自己認識系が,どのように獲得され,どのように進化してきたのか興味をもっている.
高山 誠司(Seiji Takayama)
略歴:1986年 東京大学大学院農学研究科博士課程 修了(農学博士),1995年 奈良先端科学技術大学院大学バイオサイエンス研究科 助教授を経て,2006年より同 教授.
研究テーマ:植物の細胞間情報伝達.
研究室URL:http://bsw3.naist.jp/takayama/index.html
© 2010 久保健一・円谷徹之・高山誠司 Licensed under CC 表示 2.1 日本
(奈良先端科学技術大学院大学バイオサイエンス研究科 細胞生物学専攻細胞間情報学講座)
email:久保健一,高山誠司
DOI: 10.7875/first.author.2010.059
Collaborative non-self recognition system in S-RNase-based self-incompatibility.
Ken-ichi Kubo, Tetsuyuki Entani, Akie Takara, Ning Wang, Allison M. Fields, Zhihua Hua, Mamiko Toyoda, Shin-ichi Kawashima, Toshio Ando, Akira Isogai, Teh-hui Kao, Seiji Takayama
Science, 330, 796-799 (2010)
要 約
顕花植物の自家不和合性は自殖をさまたげ他殖を促すことによって遺伝的な多様性を保つ遺伝的な機構である.ナス科植物ではS遺伝子座にコードされるFボックスタンパク質SLFが自家不和合性の花粉側因子である.SLFの対立遺伝子産物は雌しべ側因子であるS-RNaseの非自己の対立遺伝子産物を特異的に解毒することによって和合性受粉を可能にしているものと考えられてきた.しかしながら,SLFは対立遺伝子産物のあいだでの配列多型性がきわめて低く,どのようにして非自己のS-RNaseの膨大なレパートリーすべてを認識しているのか謎であった.筆者らは,in vivo機能アッセイおよびタンパク質相互作用アッセイにより,ナス科植物のペチュニアでは少なくとも3つのタイプの異なる遺伝子にコードされたSLFが花粉側因子として協調的に機能し,それぞれが一部の非自己のS-RNaseを分担して認識していることを明らかにし,S-RNase型の自家不和合性機構として協調的な非自己認識系を提唱した.
はじめに
自家不和合性は顕花植物の種内生殖障壁であり,雌しべにおける自己(遺伝的に近縁の)花粉の排除および非自己(遺伝的に無関係の)花粉の受容を可能にしている.この自己と非自己との識別は,単一のS遺伝子座の複対立遺伝子にコードされた花粉側因子と雌しべ側因子とによって調節されている1,2)(図1).花粉側因子と雌しべ側因子をコードする遺伝子は密接に連鎖して組み換わることなく遺伝するため,これら遺伝子のセットはSハプロタイプとよばれている.ナス科,バラ科,オオバコ科の自家不和合性を示す種は雌しべ側因子として分泌型リボヌクレアーゼであるS-RNaseを用いている1,3).半数体花粉のSハプロタイプが花柱のSハプロタイプのいずれかと一致した場合,S-RNaseは自己の花粉管の内部で細胞毒性を発揮してその伸長を阻害する1,3).一方,花粉側因子はSLF(S-locus F-box)と名づけられたFボックスタンパク質であり3,4),これらはSCF(Skp1-Cullin1-F-box)型ユビキチンリガーゼ複合体の構成タンパク質であることが示唆されてきた3,4).
S-RNaseによる花粉管のSハプロタイプの特異的な排除を説明するためタンパク質分解モデルが提唱されてきた(図1).このモデルでは,SLFが自己以外のすべての非自己のS-RNaseを認識し,ユビキチン-プロテアソーム系を介してこれを分解し解毒することを予想する1,3,4).このモデルは古くから知られる4倍体植物が自家和合性を示す“競合相互作用”とよばれる現象,すなわち,2つの異なる花粉側因子の対立遺伝子をもつ花粉が自家和合性を示す現象を矛盾なく説明することができる5,6)(図1).2つの対立遺伝子をもつ花粉から生じる2つの異なるSLFは互いに認識特異性を相補しあうことによってすべてのS-RNaseを分解することが可能となり,結果として,花粉管はすべてのSハプロタイプの花柱に対して和合性になると説明しうる3).実際,競合相互作用にもとづいてデザインされたin vivo機能アッセイにより,ナス科のペチュニアの一種Petunia inflataのS2ハプロタイプに由来するSLFが花粉側因子として機能することが証明された.すなわち,S2ハプロタイプに由来するSLF対立遺伝子をS1S2個体およびS2S3個体に導入する実験が行われ,その結果,異なる対立遺伝子をもつS1花粉あるいはS3花粉にS2ハプロタイプに由来するSLF対立遺伝子が導入されると自家不和合性は打破されるが,異なる対立遺伝子をもたないS2花粉の場合には自家不和合性が維持されることが示された7).
この実験結果がもととなりSLFが唯一の花粉側因子であると考えられるようになった7,8).しかし,のちの実験により,SLFはその対立遺伝子産物のあいだでの多型性がS-RNaseに比べずっと低く,また,アブラナ科植物の花粉側因子と雌しべ側因子とのあいだで認められた共進化の関係もまったく認められないことが示唆されてきた.さらに,それぞれの種が数十に及ぶSハプロタイプをもつことを考えると,単一のSLFがはたして自己以外のすべてのSハプロタイプに由来するS-RNaseのレパートリーを認識しうるかどうか疑問視されるようになってきた.そこで,この研究では,ペチュニアにおいてすでに同定されたSLFが本当に自家不和合性における唯一の花粉側因子であるのかどうかをまず検証することにした.
1.既知のSLFは唯一の花粉側因子ではない
ペチュニアのS5,S7,S9,S11のホモ接合体の花粉のcDNAから,既知のSLF対立遺伝子との配列相同性をもとに4つの新規な対立遺伝子をクローニングした.同定されたすべての対立遺伝子から推定されたSLFのアミノ酸配列はきわめて高い相同性を示し(アミノ酸同一性は86.4%~100%),相当するS-RNaseの示す低い相同性(多型性)とは対照的であった(アミノ酸同一性は40.1%~79.4%).なお,S-RNase型の自家不和合性をもつ種は一般に複数のSLF様の遺伝子をもつことが示されてきたことから8-10),混乱をさけるため,S2ハプロタイプに由来するSLF対立遺伝子と高い相同性を示すこれらのSLF対立遺伝子をタイプ1 SLFと名づけ,それぞれの対立遺伝子をSn-SLF1とよぶことにした(nはSハプロタイプの種類を表す).
驚くべきことに,S7-SLF1から推定されたアミノ酸配列はS19-SLF1 6) のものと100%同一であった.S7ハプロタイプおよびS19ハプロタイプは異なるS-RNaseをもち(アミノ酸同一性は45%),それらのホモ接合体のあいだの相互交配から異なる自他認識特異性を示すことも確認された.2つの異なるSハプロタイプにおいて同一のSLF対立遺伝子が発見されたことから,SLFが唯一の花粉側因子ではない可能性が示唆された.
この可能性を調べるため,タイプ1 SLFのそれぞれの対立遺伝子の産物が花粉側因子として機能するかどうかを形質転換実験により検証することにした.たとえば,S7-SLF1(= S19-SLF1)を用いた実験では,自己SハプロタイプをもつS7花粉およびS19花粉だけでなく,非自己SハプロタイプをもつS5花粉およびS11花粉においても競合相互作用が起こらないことが示された.一方,S7-SLF1をさらに別の非自己SハプロタイプをもつS9およびS17花粉において発現させた場合は競合相互作用が観察された.以上の結果は,S7-SLF1が一部の非自己Sハプロタイプをもつ花粉においてのみ選択的な競合相互作用を起こすことを示唆した.タイプ1 SLFのほかの対立遺伝子を用いた形質転換実験においても,同様に一部の非自己Sハプロタイプの花粉においてのみ競合相互作用が観察された.このことは,タンパク質分解モデルにもとづけば,おのおののタイプ1 SLFは非自己S-RNaseの一部を認識しそれらの分解を誘導していることを示唆していた.では,タイプ1 SLFにより認識されなかったほかの非自己S-RNaseは,S遺伝子座にコードされたいずれの因子によって認識されているのだろうか.
2.花粉側因子の特異性は複数の種類のSLF遺伝子によって決定されている
ペチュニアではいくつかのSLF様の遺伝子がS遺伝子座と強く連鎖していることが知られてきた8).そこで,このようなSLF様遺伝子にコードされた因子も花粉側因子として機能する可能性を推察した.この仮説を検証するため,既知のSLF様遺伝子の配列情報にもとづき,S5,S7,S9,S11,S17,S19それぞれのハプロタイプの花粉から新規のSLF様遺伝子のcDNA断片を徹底的にクローニングした.
結果として,30個のSLF様遺伝子を同定し,配列類似性にもとづいて5つのサブグループに分類しタイプ2 SLF~タイプ6 SLFと名づけた.タイプ1 SLFと同じ命名法を用いてそれらの対立遺伝子をSn-SLFxとよぶことにした(xはタイプの番号を表す).タイプ1 SLFの場合と同様に,いずれのタイプのSLFもSハプロタイプのあいだでの多型性は低いことが示された(アミノ酸同一性は70%以上).
6つのタイプのSLF遺伝子のすべてが類似した雄性器官に特異的な発現プロファイルを示した.すなわち,葯の発達過程において発現が増加し,開花前にピークに達し,成熟花粉および花粉管において減少した.また,それぞれのSLF遺伝子とS-RNase遺伝子とのあいだの連鎖を調べたところ,6つのタイプのSLF遺伝子のすべてがS-RNase遺伝子の1 cM以内に位置していることが示唆された.以上の結果は,新規の5つのタイプのSLF遺伝子が花粉側因子の遺伝子としての条件を満たすことを示した.
そこでつぎに,タイプ2およびタイプ3に属する5つのSLF対立遺伝子について,形質転換法によるin vivo機能アッセイを行った.その結果,S7-SLF3を除くすべてについて,タイプ1 SLFと同様に調べたいずれかの非自己Sハプロタイプをもつ花粉において競合相互作用が観察された.たとえば,S7-SLF2を用いた実験では,自己のS7ハプロタイプおよび非自己であるS5ハプロタイプあるいは非自己S17ハプロタイプをもつ花粉では競合相互作用はみられなかったが,非自己であるS9ハプロタイプ,S11ハプロタイプ,S19ハプロタイプをもつ花粉では競合相互作用が観察された.なお,今回のの実験で遺伝子導入したS7ハプロタイプおよびS11ハプロタイプに由来する3つのタイプのSLF1~SLF3遺伝子ではいずれもS5花粉においては競合相互作用が観察されず,この事実は,ほかの未解析のタイプのSLF遺伝子の産物がS5ハプロタイプに由来するS5-RNaseの解毒に関与していることを示唆した.さらなる解析は必要であるが,今回の形質転換実験は少なくとも3つ以上のタイプのSLFが花粉側因子として機能していることを示唆した.
3.SLFは非自己のS-RNaseの一部と相互作用する
複数のタイプのSLFが花粉側因子として機能していると仮定すると,タンパク質分解モデルは以下のように修正される.たとえば,S7花粉が自己のS7花柱に対して不和合性であるのはS7花粉において生産される複数のタイプのSLFのすべてがS7ハプロタイプに由来するS7-RNaseを認識しないためである.また,S7-SLF2を遺伝子導入したS5花粉あるいはS17花粉が競合相互作用を示さなかったのに対し,S9花粉,S11花粉,S19花粉は競合相互作用を示し自己の花柱に対して和合性を示すようになったというさきに述べた実験結果は,S7-SLF2がS5-RNaseあるいはS17-RNaseを認識しないのに対し,S9-RNase,S11-RNase,S19-RNaseを認識して分解するためと理解される.そこで,このモデルを検証するため,つぎにS7-SLF2を用いたタンパク質相互作用解析を行った.
まず,予備実験として,FLAGタグ配列を付加したS7-SLF2をS5S11個体の花粉に発現させ,それがS7-SLF2と同様にS11花粉での競合相互作用を誘導することを観察し,FLAGタグがS7-SLF2の機能に影響しないことを確認した.つぎに,FLAGタグを付加したS7-SLF2を含む形質転換花粉の抽出物を種々の異なるSハプロタイプの花柱抽出物と混合し,抗FLAG抗体を用いて免疫沈降した.その結果,S7-SLF2は予想どおり自己のS7-RNaseや競合相互作用を示さないS5ハプロタイプのS5-RNaseとは相互作用せず,競合相互作用を示すS9ハプロタイプおよびS11ハプロタイプに由来するS9-RNaseおよびS11-RNaseとは強く相互作用することが確認された.これらの結果は,SLFが特定の一部の非自己のS-RNaseと特異的に相互作用し,それらを解毒するというモデルをさらに強く示唆した.
4.協調的な非自己認識モデル
この研究にもとづきS-RNase型の自家不和合性に関して“協調的非自己認識モデル”を提唱した(図2).ペチュニアでは花粉側因子は複数のタイプのSLF遺伝子にコードされている.ひとつのSハプロタイプからつくられるおのおののタイプのSLFが相互作用できる非自己のS-RNaseは一部に限られるが,複数のタイプのSLF遺伝子の産物が協調的に機能することで非自己のS-RNaseのすべてを認識して分解し他家受粉を可能にしている,というモデルである.これまでに明らかにされてきたアブラナ科やケシ科における自家不和合性系ではいずれも雌しべ側因子と花粉側因子が単一の遺伝子によってコードされており,しかも,同一のSハプロタイプに由来する雌しべ側因子と花粉側因子との相互作用を介して“自己”認識が行われてその受精が阻害されていた.今回,解析したペチュニア(ナス科)の自家不和合性系では多数の花粉側因子を用いて異なるSハプロタイプに由来する“非自己”の雌しべ側因子の認識が行われており,根本からまったく異なる自他識別系であることが明らかになった1,11).
おわりに
この研究から導かれた協調的な非自己モデルは,S-RNase型の自家不和合性の研究において長年の謎であった多型性の低いSLF対立遺伝子によりきわめて多様な非自己のS-RNaseを識別するしくみについてはじめて明確な説明をあたえた.オオバコ科やバラ科の自家不和合性を示す種も単一のS-RNaseの遺伝子と複数のSLF様のFボックスタンパク質の遺伝子をS遺伝子座にもつことが示されている9,10).たとえば,バラ科のリンゴ(Malus×domestica)では12個のSLF様のFボックスタンパク質の遺伝子がS遺伝子座に連鎖している10).ナス科植物とは系統学的に離れたバラ科植物においても協調的な非自己認識モデルが適用できるかどうか,今後,明らかにしていく必要がある.また,複数のタイプのSLF遺伝子がどのように出現し,単一のS-RNase遺伝子とともにどのように進化してきたのか,また,どのようにしてそれぞれのタイプのSLFが一部の非自己のS-RNaseを分子レベルで認識しているのか,さらには,SLFにより認識された非自己のS-RNaseはどのようにして無毒化されているのか,などの課題も,今後,明らかにしていく必要がある.
文 献
- Takayama, S. & Isogai, A.: Self-incompatibility in plants. Annu. Rev. Plant Biol., 56, 467-489 (2005)[PubMed]
- Franklin-Tong, V. E. (Ed.): Self-Incompatibility in Flowering Plants. Springer, Heidelberg (2008)
- Hua, Z., Fields, A. & Kao, T. -h.: Biochemical models for S-RNase-based self-incompatibility. Mol. Plant, 1, 575-585 (2008)[PubMed]
- Entani, T., Iwano, M., Shiba, H. et al.: Comparative analysis of the self-incompatibility (S-) locus region of Prunus mume: identification of pollen-expressed F-box gene with allelic diversity. Genes Cells, 8, 203-213 (2003)[PubMed]
- Entani, T., Takayama, S., Iwano, M. et al.: Relationship between polyploidy and pollen self-incompatibility phenotype in Petunia hybrida Vilm. Biosci. Biotechnol. Biochem., 63, 1882-1888 (1999)[PubMed]
- Tsukamoto, T., Ando, T., Watanabe, H. et al.: Duplication of the S-locus F-box gene is associated with breakdown of pollen function in an S-haplotype identified in a natural population of self-incompatible Petunia axillaris. Plant Mol. Biol., 57, 141-153 (2005)[PubMed]
- Sijacic, P., Wang, X., Skirpan, A. L. et al.: Identification of the pollen determinant of S-RNase-mediated self-incompatibility. Nature, 429, 302-305 (2004)[PubMed]
- Hua, Z., Meng, X., & Kao, T. -h.: Comparison of Petunia inflata S-Locus F-box protein (Pi SLF) with Pi SLF like proteins reveals its unique function in S-RNase based self-incompatibility. Plant Cell, 19, 3593-3609 (2007)[PubMed]
- Zhou, J., Wang, F., Ma, W. et al.: Structural and transcriptional analysis of S-locus F-box genes in Antirrhinum. Sex. Plant Reprod., 16, 165-177 (2003)
- Minamikawa, M., Kakui, H., Wang, S. et al.: Apple S locus region represents a large cluster of related, polymorphic and pollen-specific F-box genes. Plant Mol. Biol., 74, 143-154 (2010)[PubMed]
- Wheeler, M. J., de Graaf, B. H. J., Hadjiosif, N. et al.: Identification of the pollen self-incompatibility determinant in Papaver rhoeas. Nature, 459, 992-995 (2009)[PubMed]
著者プロフィール
略歴:2000年 東北大学大学院工学研究科博士課程 修了(工学博士),同年 農業生物資源研究所 博士研究員を経て,2005年より奈良先端科学技術大学院大学バイオサイエンス研究科 博士研究員.
研究テーマ:ナス科植物ペチュニアにおける自家不和合性の分子機構の解明.
関心事:この研究で示した協調的な非自己認識系が,どのように獲得され,どのように進化してきたのか興味をもっている.
高山 誠司(Seiji Takayama)
略歴:1986年 東京大学大学院農学研究科博士課程 修了(農学博士),1995年 奈良先端科学技術大学院大学バイオサイエンス研究科 助教授を経て,2006年より同 教授.
研究テーマ:植物の細胞間情報伝達.
研究室URL:http://bsw3.naist.jp/takayama/index.html
© 2010 久保健一・円谷徹之・高山誠司 Licensed under CC 表示 2.1 日本
