根における局所的な窒素の欠乏を植物の全体に伝達するペプチドホルモン
田畑 亮・松林嘉克
(名古屋大学大学院理学研究科 生命理学専攻細胞間シグナル研究グループ)
email:田畑 亮,松林嘉克
DOI: 10.7875/first.author.2014.134
Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling.
Ryo Tabata, Kumiko Sumida, Tomoaki Yoshii, Kentaro Ohyama, Hidefumi Shinohara, Yoshikatsu Matsubayashi
Science, 346, 343-346 (2014)
植物にとり窒素はもっとも重要な無機栄養のひとつである.植物は地中から窒素をおもにNO3-として吸収しているが,自然界においてNO3-の地中における分布はきわめて不均一である.そのため,植物は根の一部が局所的な窒素の欠乏におちいったとき,その情報をほかの根に伝え,NO3-の取り込みを相補的に促進させるしくみをもつ.しかし,この全身的な窒素要求シグナル伝達とよばれる巧妙なしくみの分子機構は解明されていなかった.筆者らは,根の一部が局所的な窒素の欠乏を感知すると分泌型のペプチドホルモンであるCEPが産生され,それらが導管を移行して地上部のCEP受容体に認識されることがひき金となり,最終的に植物の全体に窒素の欠乏の情報が伝達されていることを明らかにした.この全身的なシグナル伝達により別の根におけるNO3-の取り込みが増大し,局所的な窒素の不足が補填されていた.この発見は,変動する環境に対する植物のたくみな適応能力の一端を明らかにしたものである.
近年,植物の形態形成や環境応答において,数アミノ酸残基から数十アミノ酸残基という短鎖のペプチドホルモンを介した細胞間シグナル伝達が重要な役割をはたしていることが明らかになりつつある1,2).これらのペプチドホルモンは,約100アミノ酸残基からなる前駆体が翻訳されたのち,C末端の付近が修飾およびプロセシングをうけ,10~20アミノ酸残基の活性型のペプチドとして切り出されて分泌される(図1a).筆者らのグループでは,既知のペプチドホルモンの前駆体の構造的な特徴や植物界における保存性にもとづき,シロイヌナズナのゲノムから新規のペプチドホルモンの候補となる遺伝子をいくつか見い出して解析してきた3,4).そのひとつがCEP(C-terminally encoded peptide)ファミリーであり,シロイヌナズナからは少なくとも11個の遺伝子が見い出されている.CEPの多くは根において特異的に発現がみられ,過剰に発現させた場合あるいは培地に合成ペプチドをくわえた場合に根の生育が抑制されたことから,根の成長に関与することが示唆された.しかしながら,CEPファミリーは高度に遺伝子重複しているため欠損株の作製が困難であり2,4),当初,その機能の解明は難航した.
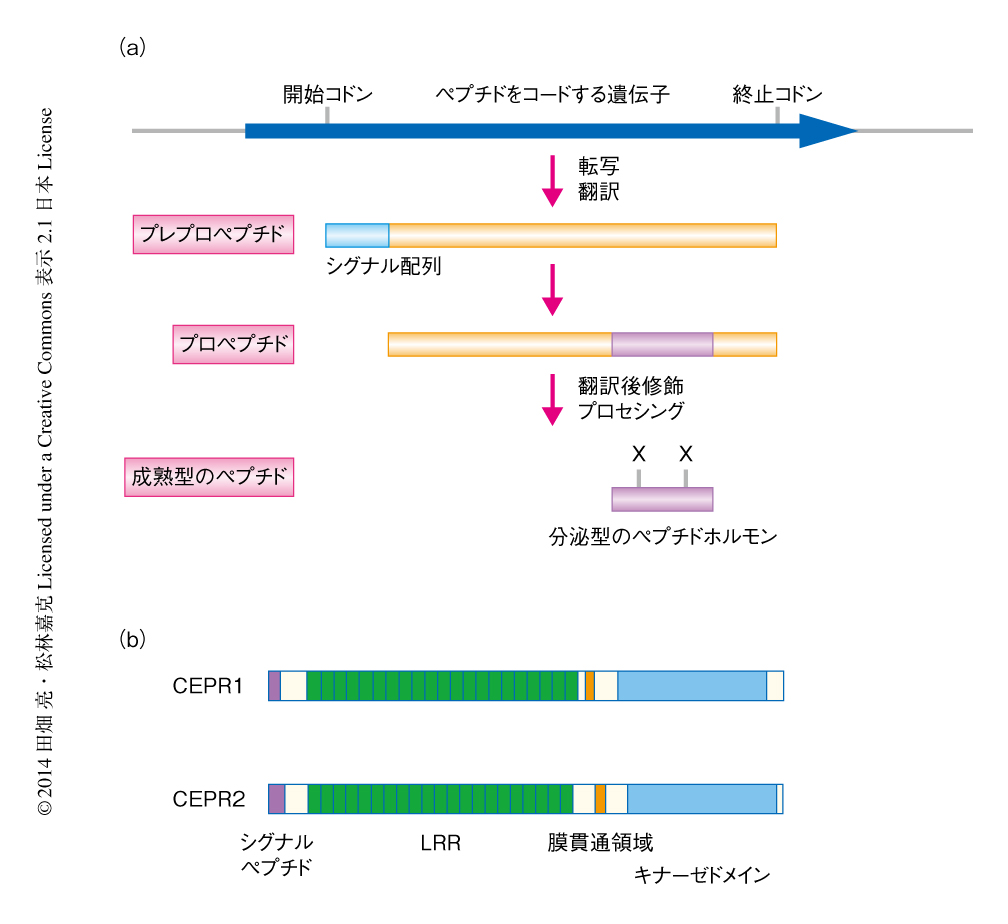
そこで,まずCEPを認識する受容体を同定し,その欠損株の解析からCEPの機能を明らかにすることを試みた.CEP受容体の候補として,短鎖のペプチドの受容体として機能するいくつかのタンパク質の含まれる,シロイヌナズナのロイシンリッチリピート型受容体キナーゼのサブファミリーXおよびサブファミリーXIの約40種類に着目した.これらを個々にタバコ培養細胞BY-2において過剰に発現させ,膜画分を回収して発現ライブラリーとした.このライブラリーに対し,光反応性のCEP1を用いた光アフィニティー標識による網羅的な結合実験を行ったところ,CEP1と直接的に結合する2つのロイシンリッチリピート型受容体キナーゼが見い出された(図1b).この2つはともにサブファミリーXIに属し5),これらの二重変異体はCEP1に対し非感受性を示したことから,CEP受容体であると結論づけCEPR1およびCEPR2と命名した.CEPR1はCEP1だけでなくCEP3やCEP5とも結合したことから,CEPファミリーに共通の受容体であると考えられた.CEPR1およびCEPR2の発現は植物の全体において観察されたが,CEPR1はとくに葉の維管束において高い発現を示した.
CEP受容体の欠損株であるcepr1 cepr2二重変異体の表現型を解析したところ,側根の伸長,葉の黄化,植物体の矮小化などの形態変化がみられた.これらの表現型は植物が窒素の欠乏におちいった際にしばしば観察されるため6),CEPが窒素応答に関与している可能性が示唆された.
CEPが窒素応答のどの過程に関与するかを調べるため,野生型とCEP受容体欠損株における遺伝子発現をマイクロアレイ法により比較した.その結果,CEP受容体欠損株の根において,主要なNO3-輸送体をコードするNRT2.1遺伝子,NRT3.1遺伝子,NRT1.1遺伝子の発現がいちじるしく低下していた.実際に,CEP受容体欠損株の根を用いて安定同位体により標識したNO3-の取り込みについて調べたところ,野生型と比べ活性は約半分に低下していた.
さらに,これらNO3-輸送体の発現が実際にCEPにより制御されているのかどうかを解析した.通常の条件において生育させた植物をCEP1の含まれる培地に移し,根においてNO3-輸送体の発現を解析したところ,CEP1の処理ののち6時間以内に発現の上昇がみられ,24時間後には発現は5倍以上に上昇することが確かめられた.ほかのCEPファミリーにもこの活性は認められた.これらの結果より,NO3-輸送体の発現はCEPにより制御されていることが示された.
NO3-輸送体であるNRT2.1,NRT3.1,NRT1.1は全身的な窒素要求シグナル伝達により制御されていることが知られている7,8).この全身的な窒素要求シグナル伝達とは,一部の根の周辺においてNO3-が不足した場合に別の根においてNO3-の取り込みを増大させてこれを相補する制御系であり,土壌においてNO3-の分布が不均一な環境で生きぬくために植物が進化させた巧妙な系である.しかし,その分子機構についてはよくわかっていなかった.全身的な窒素要求シグナル伝達の標的として知られていたほかの複数の遺伝子のいずれもがCEP受容体欠損株では発現が低下していたことや,分泌型のペプチドホルモンは組織の内部を移行しうることを考慮して,CEPが全身的な窒素要求シグナル伝達に関与しているという仮説をたてその実証を進めた.
CEPの発現が窒素の濃度に応じて制御されているかどうかを調べた.窒素が十分に含まれる培地で生育させた野生型の植物を窒素を含まない培地に移しCEPの発現について解析したところ,6時間後にはCEP1,CEP3,CEP5,CEP6,CEP7,CEP8,CEP9の発現が上昇し,24時間後には数倍から十数倍になることが示された.これに対して,リンあるいはカリウムの欠乏に対しては変化を示さなかったことから,CEPの発現は窒素の欠乏に対し特異的に誘導されることが確かめられた.
CEP受容体の欠損が全身的な窒素要求シグナル伝達にあたえる影響について調べた.これには,1本の植物から出る根を2束に分け,それぞれを異なる条件にさらすという培養系を用いた.一方の根は十分なNO3-の存在する培地,もう一方の根はNO3-の存在しない培地で生育させたところ,野生型では窒素の存在しないほうの根においてCEPの発現の上昇をともないつつ,窒素の十分に存在するもう一方の根においてNO3-輸送体NRT2.1の発現が上昇した.それに対して,CEP受容体欠損株では窒素の存在しないほうの根においてCEPの発現は上昇したものの,窒素の十分に存在するもう一方の根におけるNRT2.1の発現の上昇はみられなかった.すなわち,CEP受容体欠損株においては全身的な窒素要求シグナル伝達は起こらないことが確かめられた.
さらに,同じ培養系を用いて,両方の根を十分なNO3-の存在する培地で生育させ,一方の根のみをCEP1により処理した際のNRT2.1の発現について解析した.その結果,CEP1を処理したほうの根のみならず,もう一方の根においてもNRT2.1の発現は上昇することが明らかになった.この全身的な反応は少なくとも6時間以内に観察された.以上の結果は,窒素の欠乏により根において発現の誘導されるCEPが全身的な窒素要求シグナル伝達に関与していることを示した.
窒素の欠乏により発現の誘導された根に由来するCEPが,どこでCEP受容体に認識され,最終的に離れた根においてNO3-輸送体の発現の上昇をひき起こしているのかについて検討した.これには,接木と根を2束に分けて異なる条件にさらす培養系とを組み合わせた実験系を用いた(図2a).
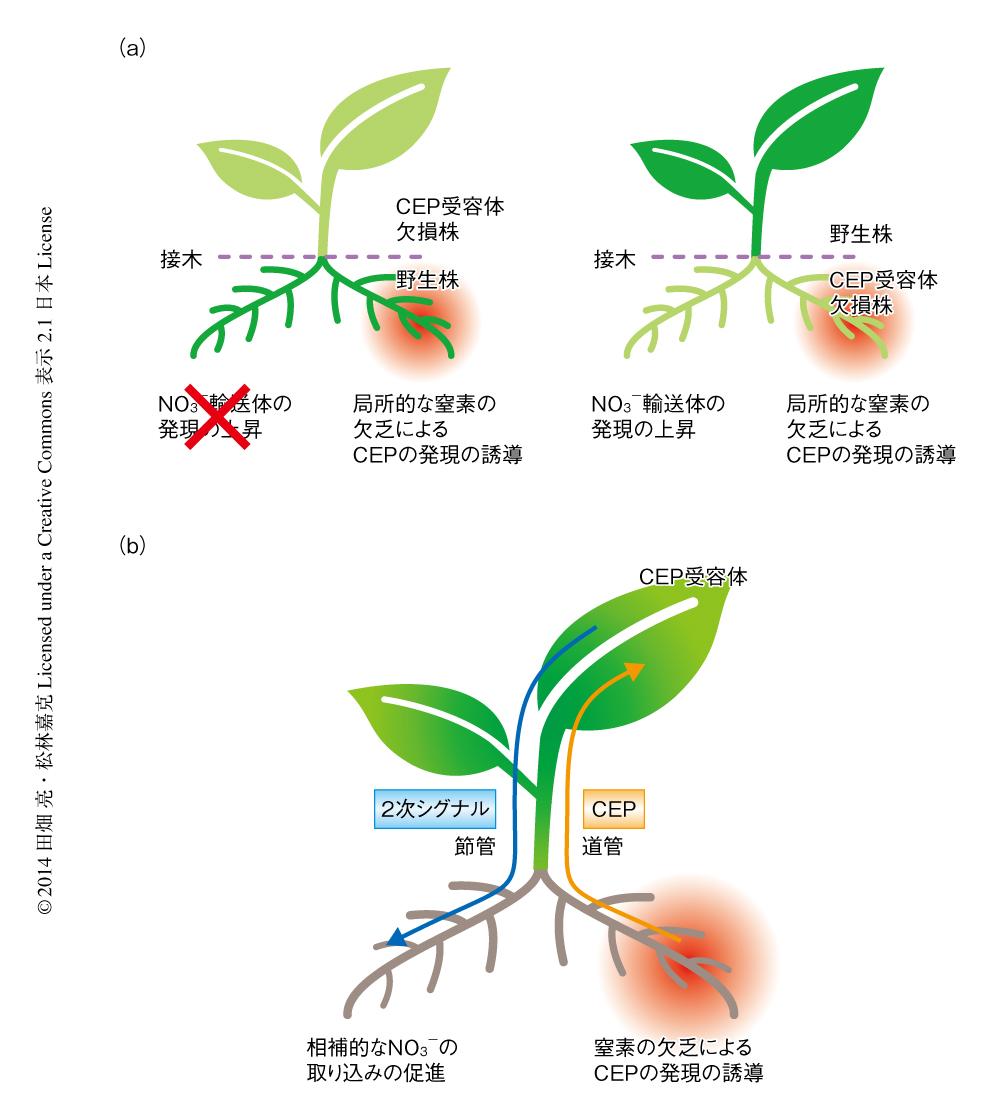
地上部の接穂にCEP受容体欠損株,地下部の台木に野生型を組み合わせて胚軸のあいだで接木した植物においては,一方の根をNO3-の存在しない培地で生育させても,もう一方の根においてNO3-輸送体NRT2.1の発現の上昇はみられなかった.また,この接木した植物の両方の根を十分なNO3-の存在する培地で生育させ,一方の根にのみCEP1を処理した場合,NRT2.1の発現はいずれの側の根でも上昇しなかった.逆に,地上部が野生型,地下部はCEP受容体欠損株の組合せにより接木した植物では,一方の根をNO3-の存在しない培地で生育させたとき,もう一方の根においてNRT2.1の発現の上昇が観察された.また,一方の根にCEPを処理した場合,処理した側だけでなくもう一方の根においてもNRT2.1の発現の上昇がみられた.これらの結果は,根に由来するCEPの地上部のCEP受容体による認識が,全身的なNRT2.1の発現の上昇のため必須であることを示した.
地上部のCEP受容体においてCEPが認識されるのならば,窒素の欠乏により根において発現の誘導されたCEPは根から地上部へと移動する必要がある.このことを確かめるため,シロイヌナズナの胚軸を切断して道管液を回収し,内生するCEPについて解析した.その結果,窒素を欠乏したときの道管液にはCEP3,CEP6,CEP9などいくつかのCEPファミリーが検出されたが,窒素を十分にあたえた植物に由来する道管液からはほとんど検出されないことが確かめられた.すなわち,根における窒素の欠乏を伝えるシグナルとして,CEPは根から地上部へ道管をとおり輸送されていた.
以上の結果をまとめると,つぎのようなモデルが考えられた(図2b).土壌において根の一部が窒素の欠乏を感知すると,分泌型のペプチドホルモンであるCEPが産生され,道管をとおり地上部へと輸送される.地上部に達したCEPは,葉の維管束において発現するロイシンリッチリピート型受容体キナーゼであるCEP受容体により認識され,全身に窒素の欠乏を伝達する未知の2次シグナルを誘導する.この2次シグナルは,おそらく篩管をとおってふたたび根に移行し,周囲にNO3-の存在するほかの根にはたらきかけてNRT2.1などNO3-輸送体の発現を上昇させる.
これまで,植物は一部の根の周辺においてNO3-が欠乏すると別の根においてNO3-の取り込みを増大させるという応答を示すことは知られていたが,その分子機構はほとんど明らかにされていなかった.今回の発見により,根において産生される分泌型のペプチドホルモンCEPを,葉においてCEP受容体が受け取ることが,全身的な窒素要求シグナル伝達の根幹をなすことが明らかにされた.これは,動きまわることのできない植物が進化させた環境応答のためのたくみな系のひとつといえるだろう.今後は,CEPがCEP受容体により認識されたのち葉において産生され根に輸送されると考えられる,未知の2次シグナルの解明に興味がもたれる.
略歴:2010年 名古屋大学大学院生命農学研究科にて博士号取得,熊本大学大学院自然科学研究科 博士研究員などを経て,2014年より名古屋大学大学院理学研究科 博士研究員.
研究テーマ:ペプチドホルモンを介した植物の形態形成および環境応答.
関心事:植物の協調的な器官形成をになう細胞間および器官間のシグナル伝達機構.釣り.広島東洋カープ.
松林 嘉克(Yoshikatsu Matsubayashi)
名古屋大学大学院理学研究科 教授.
研究室URL:http://www.bio.nagoya-u.ac.jp/~b2
© 2014 田畑 亮・松林嘉克 Licensed under CC 表示 2.1 日本
(名古屋大学大学院理学研究科 生命理学専攻細胞間シグナル研究グループ)
email:田畑 亮,松林嘉克
DOI: 10.7875/first.author.2014.134
Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling.
Ryo Tabata, Kumiko Sumida, Tomoaki Yoshii, Kentaro Ohyama, Hidefumi Shinohara, Yoshikatsu Matsubayashi
Science, 346, 343-346 (2014)
要 約
植物にとり窒素はもっとも重要な無機栄養のひとつである.植物は地中から窒素をおもにNO3-として吸収しているが,自然界においてNO3-の地中における分布はきわめて不均一である.そのため,植物は根の一部が局所的な窒素の欠乏におちいったとき,その情報をほかの根に伝え,NO3-の取り込みを相補的に促進させるしくみをもつ.しかし,この全身的な窒素要求シグナル伝達とよばれる巧妙なしくみの分子機構は解明されていなかった.筆者らは,根の一部が局所的な窒素の欠乏を感知すると分泌型のペプチドホルモンであるCEPが産生され,それらが導管を移行して地上部のCEP受容体に認識されることがひき金となり,最終的に植物の全体に窒素の欠乏の情報が伝達されていることを明らかにした.この全身的なシグナル伝達により別の根におけるNO3-の取り込みが増大し,局所的な窒素の不足が補填されていた.この発見は,変動する環境に対する植物のたくみな適応能力の一端を明らかにしたものである.
はじめに
近年,植物の形態形成や環境応答において,数アミノ酸残基から数十アミノ酸残基という短鎖のペプチドホルモンを介した細胞間シグナル伝達が重要な役割をはたしていることが明らかになりつつある1,2).これらのペプチドホルモンは,約100アミノ酸残基からなる前駆体が翻訳されたのち,C末端の付近が修飾およびプロセシングをうけ,10~20アミノ酸残基の活性型のペプチドとして切り出されて分泌される(図1a).筆者らのグループでは,既知のペプチドホルモンの前駆体の構造的な特徴や植物界における保存性にもとづき,シロイヌナズナのゲノムから新規のペプチドホルモンの候補となる遺伝子をいくつか見い出して解析してきた3,4).そのひとつがCEP(C-terminally encoded peptide)ファミリーであり,シロイヌナズナからは少なくとも11個の遺伝子が見い出されている.CEPの多くは根において特異的に発現がみられ,過剰に発現させた場合あるいは培地に合成ペプチドをくわえた場合に根の生育が抑制されたことから,根の成長に関与することが示唆された.しかしながら,CEPファミリーは高度に遺伝子重複しているため欠損株の作製が困難であり2,4),当初,その機能の解明は難航した.
1.CEP受容体の同定
そこで,まずCEPを認識する受容体を同定し,その欠損株の解析からCEPの機能を明らかにすることを試みた.CEP受容体の候補として,短鎖のペプチドの受容体として機能するいくつかのタンパク質の含まれる,シロイヌナズナのロイシンリッチリピート型受容体キナーゼのサブファミリーXおよびサブファミリーXIの約40種類に着目した.これらを個々にタバコ培養細胞BY-2において過剰に発現させ,膜画分を回収して発現ライブラリーとした.このライブラリーに対し,光反応性のCEP1を用いた光アフィニティー標識による網羅的な結合実験を行ったところ,CEP1と直接的に結合する2つのロイシンリッチリピート型受容体キナーゼが見い出された(図1b).この2つはともにサブファミリーXIに属し5),これらの二重変異体はCEP1に対し非感受性を示したことから,CEP受容体であると結論づけCEPR1およびCEPR2と命名した.CEPR1はCEP1だけでなくCEP3やCEP5とも結合したことから,CEPファミリーに共通の受容体であると考えられた.CEPR1およびCEPR2の発現は植物の全体において観察されたが,CEPR1はとくに葉の維管束において高い発現を示した.
2.CEP受容体の欠損株ではNO3-輸送体の発現が低下している
CEP受容体の欠損株であるcepr1 cepr2二重変異体の表現型を解析したところ,側根の伸長,葉の黄化,植物体の矮小化などの形態変化がみられた.これらの表現型は植物が窒素の欠乏におちいった際にしばしば観察されるため6),CEPが窒素応答に関与している可能性が示唆された.
CEPが窒素応答のどの過程に関与するかを調べるため,野生型とCEP受容体欠損株における遺伝子発現をマイクロアレイ法により比較した.その結果,CEP受容体欠損株の根において,主要なNO3-輸送体をコードするNRT2.1遺伝子,NRT3.1遺伝子,NRT1.1遺伝子の発現がいちじるしく低下していた.実際に,CEP受容体欠損株の根を用いて安定同位体により標識したNO3-の取り込みについて調べたところ,野生型と比べ活性は約半分に低下していた.
さらに,これらNO3-輸送体の発現が実際にCEPにより制御されているのかどうかを解析した.通常の条件において生育させた植物をCEP1の含まれる培地に移し,根においてNO3-輸送体の発現を解析したところ,CEP1の処理ののち6時間以内に発現の上昇がみられ,24時間後には発現は5倍以上に上昇することが確かめられた.ほかのCEPファミリーにもこの活性は認められた.これらの結果より,NO3-輸送体の発現はCEPにより制御されていることが示された.
3.CEPは全身的な窒素要求シグナル伝達に関与する
NO3-輸送体であるNRT2.1,NRT3.1,NRT1.1は全身的な窒素要求シグナル伝達により制御されていることが知られている7,8).この全身的な窒素要求シグナル伝達とは,一部の根の周辺においてNO3-が不足した場合に別の根においてNO3-の取り込みを増大させてこれを相補する制御系であり,土壌においてNO3-の分布が不均一な環境で生きぬくために植物が進化させた巧妙な系である.しかし,その分子機構についてはよくわかっていなかった.全身的な窒素要求シグナル伝達の標的として知られていたほかの複数の遺伝子のいずれもがCEP受容体欠損株では発現が低下していたことや,分泌型のペプチドホルモンは組織の内部を移行しうることを考慮して,CEPが全身的な窒素要求シグナル伝達に関与しているという仮説をたてその実証を進めた.
CEPの発現が窒素の濃度に応じて制御されているかどうかを調べた.窒素が十分に含まれる培地で生育させた野生型の植物を窒素を含まない培地に移しCEPの発現について解析したところ,6時間後にはCEP1,CEP3,CEP5,CEP6,CEP7,CEP8,CEP9の発現が上昇し,24時間後には数倍から十数倍になることが示された.これに対して,リンあるいはカリウムの欠乏に対しては変化を示さなかったことから,CEPの発現は窒素の欠乏に対し特異的に誘導されることが確かめられた.
CEP受容体の欠損が全身的な窒素要求シグナル伝達にあたえる影響について調べた.これには,1本の植物から出る根を2束に分け,それぞれを異なる条件にさらすという培養系を用いた.一方の根は十分なNO3-の存在する培地,もう一方の根はNO3-の存在しない培地で生育させたところ,野生型では窒素の存在しないほうの根においてCEPの発現の上昇をともないつつ,窒素の十分に存在するもう一方の根においてNO3-輸送体NRT2.1の発現が上昇した.それに対して,CEP受容体欠損株では窒素の存在しないほうの根においてCEPの発現は上昇したものの,窒素の十分に存在するもう一方の根におけるNRT2.1の発現の上昇はみられなかった.すなわち,CEP受容体欠損株においては全身的な窒素要求シグナル伝達は起こらないことが確かめられた.
さらに,同じ培養系を用いて,両方の根を十分なNO3-の存在する培地で生育させ,一方の根のみをCEP1により処理した際のNRT2.1の発現について解析した.その結果,CEP1を処理したほうの根のみならず,もう一方の根においてもNRT2.1の発現は上昇することが明らかになった.この全身的な反応は少なくとも6時間以内に観察された.以上の結果は,窒素の欠乏により根において発現の誘導されるCEPが全身的な窒素要求シグナル伝達に関与していることを示した.
4.根に由来するCEPは地上部のCEP受容体により認識される
窒素の欠乏により発現の誘導された根に由来するCEPが,どこでCEP受容体に認識され,最終的に離れた根においてNO3-輸送体の発現の上昇をひき起こしているのかについて検討した.これには,接木と根を2束に分けて異なる条件にさらす培養系とを組み合わせた実験系を用いた(図2a).
地上部の接穂にCEP受容体欠損株,地下部の台木に野生型を組み合わせて胚軸のあいだで接木した植物においては,一方の根をNO3-の存在しない培地で生育させても,もう一方の根においてNO3-輸送体NRT2.1の発現の上昇はみられなかった.また,この接木した植物の両方の根を十分なNO3-の存在する培地で生育させ,一方の根にのみCEP1を処理した場合,NRT2.1の発現はいずれの側の根でも上昇しなかった.逆に,地上部が野生型,地下部はCEP受容体欠損株の組合せにより接木した植物では,一方の根をNO3-の存在しない培地で生育させたとき,もう一方の根においてNRT2.1の発現の上昇が観察された.また,一方の根にCEPを処理した場合,処理した側だけでなくもう一方の根においてもNRT2.1の発現の上昇がみられた.これらの結果は,根に由来するCEPの地上部のCEP受容体による認識が,全身的なNRT2.1の発現の上昇のため必須であることを示した.
地上部のCEP受容体においてCEPが認識されるのならば,窒素の欠乏により根において発現の誘導されたCEPは根から地上部へと移動する必要がある.このことを確かめるため,シロイヌナズナの胚軸を切断して道管液を回収し,内生するCEPについて解析した.その結果,窒素を欠乏したときの道管液にはCEP3,CEP6,CEP9などいくつかのCEPファミリーが検出されたが,窒素を十分にあたえた植物に由来する道管液からはほとんど検出されないことが確かめられた.すなわち,根における窒素の欠乏を伝えるシグナルとして,CEPは根から地上部へ道管をとおり輸送されていた.
5.ペプチドホルモンCEPとCEP受容体を介した全身的な窒素要求シグナル伝達
以上の結果をまとめると,つぎのようなモデルが考えられた(図2b).土壌において根の一部が窒素の欠乏を感知すると,分泌型のペプチドホルモンであるCEPが産生され,道管をとおり地上部へと輸送される.地上部に達したCEPは,葉の維管束において発現するロイシンリッチリピート型受容体キナーゼであるCEP受容体により認識され,全身に窒素の欠乏を伝達する未知の2次シグナルを誘導する.この2次シグナルは,おそらく篩管をとおってふたたび根に移行し,周囲にNO3-の存在するほかの根にはたらきかけてNRT2.1などNO3-輸送体の発現を上昇させる.
おわりに
これまで,植物は一部の根の周辺においてNO3-が欠乏すると別の根においてNO3-の取り込みを増大させるという応答を示すことは知られていたが,その分子機構はほとんど明らかにされていなかった.今回の発見により,根において産生される分泌型のペプチドホルモンCEPを,葉においてCEP受容体が受け取ることが,全身的な窒素要求シグナル伝達の根幹をなすことが明らかにされた.これは,動きまわることのできない植物が進化させた環境応答のためのたくみな系のひとつといえるだろう.今後は,CEPがCEP受容体により認識されたのち葉において産生され根に輸送されると考えられる,未知の2次シグナルの解明に興味がもたれる.
文 献
- Butenko, M. A., Vie, A. K., Brembu, T. et al.: Plant peptides in signaling: looking for new partners. Trends Plant Sci., 14, 255-263 (2009)[PubMed]
- Matsubayashi, Y.: Posttranslationally modified small-peptide signals in plants. Annu. Rev. Plant Biol., 65, 385-413 (2014)[PubMed]
- Matsuzaki, Y., Ogawa-Ohnishi, M., Mori, A. et al.: Secreted peptide signals required for maintenance of root stem cell niche in Arabidopsis. Science, 329, 1065-1067 (2010)[PubMed] [新着論文レビュー]
- Ohyama, K., Ogawa, M. & Matsubayashi, Y.: Identification of a biologically active, small, secreted peptide in Arabidopsis by in silico gene screening, followed by LC-MS-based structure analysis. Plant J., 55, 152-160 (2008)[PubMed]
- Shiu, S. H. & Bleecker, A. B.: Expansion of the receptor-like kinase/Pelle gene family and receptor-like proteins in Arabidopsis. Plant Physiol., 132, 530-543 (2003)[PubMed]
- Vidal, E. A. & Gutierrez, R. A.: A systems view of nitrogen nutrient and metabolite responses in Arabidopsis. Curr. Opin. Plant Biol., 11, 521-529 (2008)[PubMed]
- Ruffel, S., Freixes, S., Balzergue, S. et al.: Systemic signaling of the plant nitrogen status triggers specific transcriptome responses depending on the nitrogen source in Medicago truncatula. Plant Physiol., 146, 2020-2035 (2008)[PubMed]
- Ruffel, S., Krouk, G., Ristova, D. et al.: Nitrogen economics of root foraging: transitive closure of the nitrate-cytokinin relay and distinct systemic signaling for N supply vs. demand. Proc. Natl. Acad. Sci. USA, 108, 18524-19529 (2011)[PubMed]
著者プロフィール
略歴:2010年 名古屋大学大学院生命農学研究科にて博士号取得,熊本大学大学院自然科学研究科 博士研究員などを経て,2014年より名古屋大学大学院理学研究科 博士研究員.
研究テーマ:ペプチドホルモンを介した植物の形態形成および環境応答.
関心事:植物の協調的な器官形成をになう細胞間および器官間のシグナル伝達機構.釣り.広島東洋カープ.
松林 嘉克(Yoshikatsu Matsubayashi)
名古屋大学大学院理学研究科 教授.
研究室URL:http://www.bio.nagoya-u.ac.jp/~b2
© 2014 田畑 亮・松林嘉克 Licensed under CC 表示 2.1 日本
