運動学習中に大脳皮質において起こる層に特異的な神経活動の変化
田中康裕・正水芳人・松崎政紀
(基礎生物学研究所 光脳回路研究部門)
email:田中康裕,正水芳人,松崎政紀
DOI: 10.7875/first.author.2014.095
Two distinct layer-specific dynamics of cortical ensembles during learning of a motor task.
Yoshito Masamizu, Yasuhiro R. Tanaka, Yasuyo H. Tanaka, Riichiro Hira, Fuki Ohkubo, Kazuo Kitamura, Yoshikazu Isomura, Takashi Okada, Masanori Matsuzaki
Nature Neuroscience, 17, 987-994 (2014)
1次運動野は運動出力を5b層から出すが,2つの中間層として2/3層および5a層をもつ.どちらの層もさまざまな脳部位から入力をうけ大脳皮質に投射を出し,それにくわえて,5a層は皮質下への投射をもつ.それゆえ,2/3層および5a層においては運動学習による神経回路の再編成は異なっていると考えられる.筆者らは,レバー引きの運動学習を行っているマウスにおいて1次運動野の2光子Ca2+イメージングを行い,ニューロンの集団および単一のニューロンからのレバー軌跡の予測精度を評価した.2/3層ではニューロンの集団からのレバー軌跡の予測精度は全体としてあまり変わらなかったものの,一部のニューロンは学習期間をとおして高い予測精度を示した.一方,5a層ではニューロンの集団からのレバー軌跡の予測精度は学習をつうじて改善し,皮質下に投射するニューロンを含む33%のニューロンが学習後期における集団による予測に貢献するようになった.2/3層の神経回路は学習をつうじてほかの皮質領野からの情報をうまく統合しながら変化していき,一方で,5a層ではよく学習された運動を表現する学習回路に加担するようになったものと考えられた.
大脳の表面をつつむように広がる大脳皮質はさまざまな脳部位との結合をもちながら,それ自体が精緻に組織化された6層の構造をもつ.おのおのの層はおおまかではあるが入力および出力に特異性をもち,この6層の構造,および,その入出力の特異性は哺乳類のあいだで種をこえて,また,機能の異なる皮質領野のあいだでも,ある程度は保存されている.運動を直接に支配していると考えられる1次運動野では,深層にあたる5層において2つの下位構造が知られており,そのひとつである5b層を中心に大脳皮質の主たる運動出力といえる皮質脊髄投射ニューロンが局在する.一方,5a層には皮質脊髄投射ニューロンにくわえ,反対側の線条体へ投射する交差性皮質線条体投射ニューロンが多く局在している1,2).線条体を含む大脳基底核はさまざまな神経変性疾患とかかわり,運動の制御および実行における重要な役割が知られている.一方,皮質の浅層である2/3層は大脳皮質のほかの領野から入力をうけ,それらの領野へ演算の結果を送り返すと考えられている.
近年,2光子顕微鏡技術の進展にともない急速に発展した生体内Ca2+イメージング法により,大脳皮質の2/3層を中心に神経活動の時空間的な特性が調べられるようになった3).この手法の利点は,おおよそ500μm四方に含まれる数百個のニューロンを同時に観察できることである.一方で,レーザーの光学的な特性によるため脳の深い位置での計測には制限があり,これまでの知見は2/3層のものにかたよっていた4-6).今回,筆者らは,2週間にわたる運動学習中に5a層から神経活動を継続的に記録し,学習による特異的な活動変化を検出した.
マウスは右前肢によるレバー引きを2週間にわたって学習した(図1).レバーを引くと口の前にあるノズルから水が出るため,これを報酬としてマウスは自らのリズムでレバーを引くようになる5,7).毎日1時間のセッションを行い,成功回数,成功率,成功間隔時間により学習の進行を定量した.成績は1日目から4日目までは急激に上がり,10日程度で高止まりした.以下,1日目から4日目を学習初期,11日目から14日目を学習後期とする.
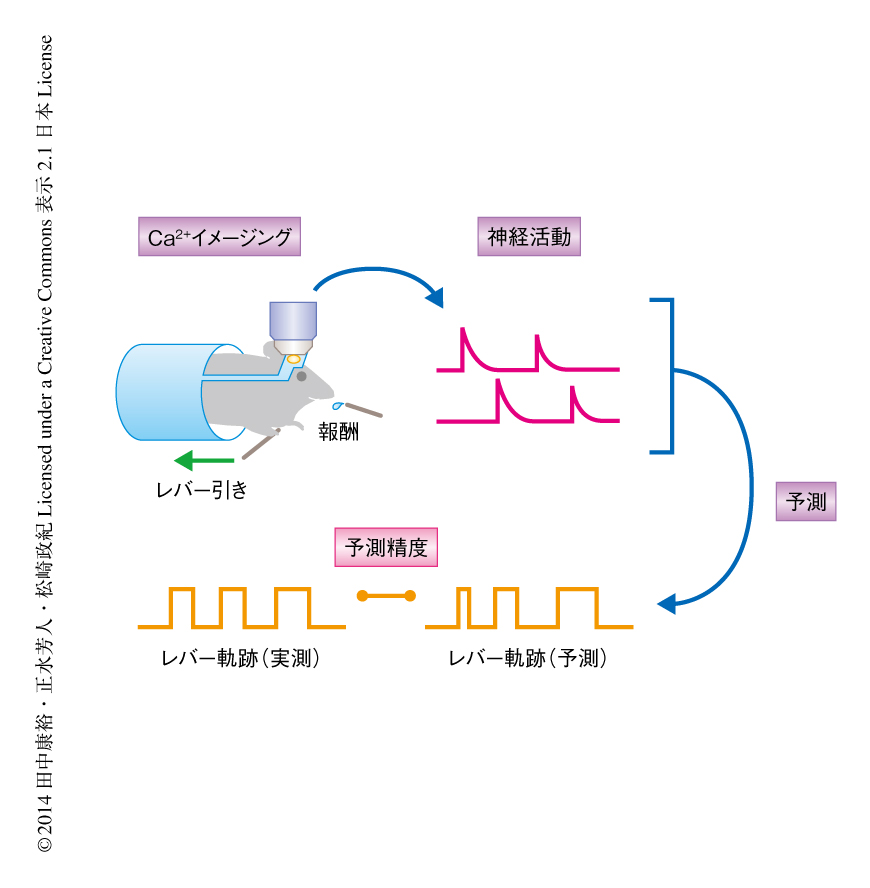
レバー引きの実行中に左脳の前肢領域においてCa2+イメージングを行った.Ca2+指示体としては蛍光タンパク質GFP,Ca2+応答タンパク質であるカルモジュリン,ミオシン軽鎖キナーゼのC末端側からつくられたGCaMP3 8) を用い,アデノ随伴ウイルスを用いて導入した.Ca2+イメージングは2光子顕微鏡を用いて3~4.5 Hzの周波数で行い,ゆれなどのアーティファクトを取り除いたのち,1視野あたり20~100個のニューロンの輝度の時系列を抽出した.これらの神経活動の時系列と同時に,レバー引きの時系列も記録しておき,神経活動からレバーの軌跡をどのくらい予測できるかにより両者の関連性を定量した.この際,予測精度を調べるため交差検定という手法を用いた9).時系列データを10分割した9/10の期間において神経活動とレバー軌跡との関係を統計的な方法により数式で定め,この定められた数式にしたがい残り1/10の期間の神経活動からレバー軌跡を予測し,予測されたレバー軌跡と実際に記録されたレバー軌跡との類似度を相互情報量という尺度で定量した.この作業を10分割したそれぞれの区間について行い,ニューロンの集団あるいは単一のニューロンからのレバー軌跡の予測精度を決定した.予測精度が高いほど神経活動とレバー軌跡との関係が保たれていたことになり,ニューロンの集団あるいは単一のニューロンがレバー引き運動の情報を保持していたと考えられる.
学習初期および学習後期において,ニューロンの集団の神経活動からどのくらいレバー軌跡を予測できるか調べた(図2).2/3層ではレバー軌跡の予測精度が上がる視野もあれば下がる視野もありさまざまであったが,5a層では調べた6匹のマウスの7つの視野のすべてで予測精度が上がった.これは,5a層ではレバー引き運動の情報が少なくともセッション内では保持されるようになったことを示唆した.このように,5a層にレバー引き運動の情報が保持されるようになったことは運動学習と関係があるのだろうか.これをみるため,セッションごとのレバー引きの成功回数や成功率と予測精度との関係を調べた.すると,5a層のニューロンの集団によるレバー引き運動の情報の保持がよくなるほど,運動学習を上手にこなしていた.一方,2/3層では5a層のように明瞭な関係は観測されなかった.
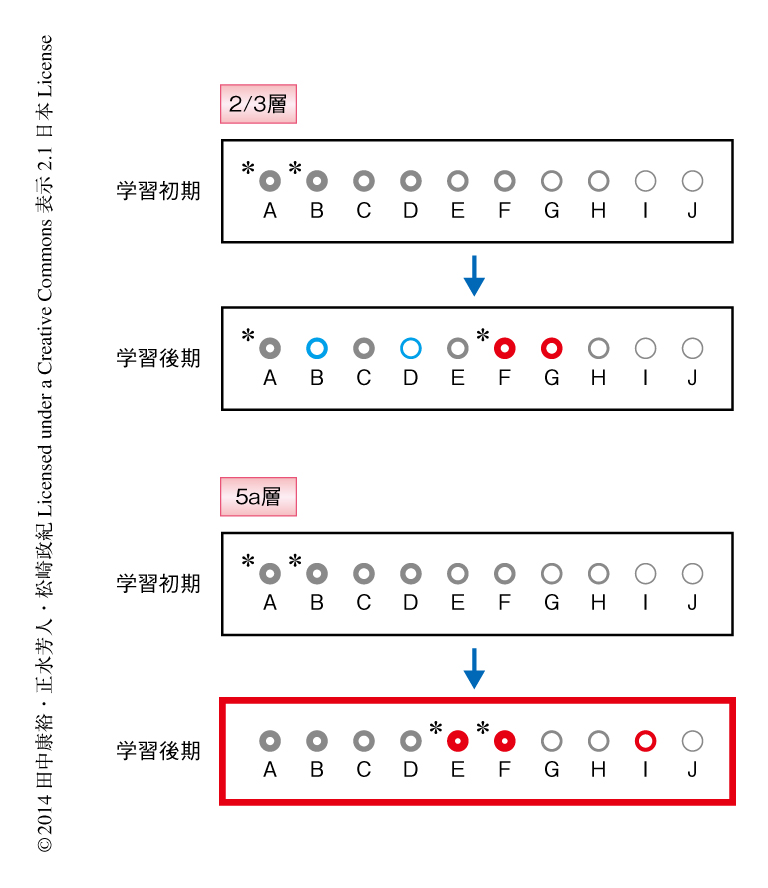
2/3層および5a層それぞれ7つの視野からのべ約8千個のニューロンの活動を記録したが,そのなかで,2/3層において126個,5a層において187個のニューロンを継続的に記録した(図2).継続的な記録とは,学習初期と学習後期,おのおの最低1日を含む8日間以上にわたり同一のニューロンを記録できたことをさす.これら継続的に記録した単一のニューロンの神経活動からレバー軌跡を予測して予測精度の変化を調べたところ,2/3層では19%のニューロンがレバー引き運動の情報の保持がよくなる一方で,17%のニューロンは情報の保持が悪くなっていた.5a層では33%のニューロンがレバー引き運動の情報の保持がよくなり,情報の保持が悪くなったニューロンは3%のみであった.5a層においてレバー引き運動の情報の保持がよくなったニューロンの活動を調べると,レバー引きのときの神経活動が学習初期に比べ学習後期において高くなっていることがわかった.
単一のニューロンの神経活動の変化は,ニューロンの集団からのレバー軌跡の予測精度にどのような影響をあたえているのだろうか.この問題を調べるため,以下の仮想実験を行った.まず,視野ごとに10個のニューロンを無作為に選び,このニューロンの集団からのレバー軌跡の予測精度をもとめる.つぎに,レバー引き運動の情報の保持がよくなったニューロンがその集団に含まれていた場合,同じ視野から記録されたニューロンに無作為に置き換え,レバー軌跡の予測精度を計算しなおした.すると,運動学習の効果とニューロンの集団からのレバー軌跡の予測精度との関係性がくずれることがわかった.このことから,単一のニューロンとしてレバー引き運動の情報を保持するようになるニューロンが,ニューロンの集団からのレバー軌跡の予測精度と運動学習の効果とを結びつけるのに必須であることがわかった.
ここまで,単一のニューロンからのレバー軌跡の予測精度が絶対値としてどのように変化するかに焦点をあててきたが,つぎに,これらの単一のニューロンが集団のなかでの位置づけをどのように変化させたかを調べた.この問題に取り組むため,継続的に記録された単一のニューロンをレバー軌跡の予測精度によりおのおのの視野においてランクづけし,そのランクの変化を調べた.2/3層では学習初期から学習後期にかけてランクの高いニューロンが比較的保たれていたのに対し,5a層ではランクの高いニューロンの多くが学習初期から学習後期にかけて入れ替わっていることがわかった.したがって,5a層ではレバー引き運動の情報を保持するようになったニューロンが集団のなかでもぬきんでることにより,ニューロンの集団からのレバー軌跡の予測精度を上げるという構図がみえてきた.一方,2/3層ではランクが高く保たれているニューロンが存在し,それらの神経活動はレバー軌跡の予測精度との関連性が学習の全期間をつうじ高かった.これらの神経活動はレバーを動かすときに必須の要素的な運動や付随する感覚と関連しているのかもしれない.
5a層は皮質下に投射するニューロンを多くもつ.なかでも,反対側の線条体に投射する交差性皮質線条体投射ニューロンおよび皮質脊髄投射ニューロンに着目し,14日間の学習期間においてどのような神経活動の変化を示すか調べた.交差性皮質線条体投射ニューロンおよび皮質脊髄投射ニューロンのいずれも30%程度が予測精度を向上させ,5%程度が予測精度を低下させた.ところが,交差性皮質線条体投射ニューロンでは学習初期から学習後期にかけて高いランクを維持しつづけるニューロンが20%程度だったのに対し,皮質脊髄投射ニューロンでは学習初期にランクの高いニューロンのすべてが学習後期にも高いランクを示した.これらの結果から,交差性皮質線条体投射ニューロンと皮質脊髄投射ニューロンのどちらも予測精度を変化させるニューロンの割合は5a層の全体と同じ程度であることがわかった.それにくわえ,新たにレバー引きを表現するようになった神経回路の一部として交差性皮質線条体投射ニューロンが組み込まれる可能性が示唆された.皮質脊髄投射ニューロンのなかで学習初期から学習後期までレバー引き運動の情報を保持できるニューロンは,レバー引き運動に必須の動きをつかさどるなど,レバー引き運動ととくに関連の高いニューロンであると考えられた.
この研究においては,ニューロンの集団あるいは単一のニューロンがレバー引き運動を表現しているかどうかに関して,神経デコーディング10) という手法を用いてアプローチした.これは,多数のニューロンの神経活動からレバー引き運動をどれだけ説明できるかを調べたものである.データ解析の視点からは,多次元の神経活動から1次元の時系列を予測することにより,把握しづらい多次元の情報の次元を減らして可視化したともいえる.同じように,多次元の情報を縮約して表現する方法には主成分分析や因子分析といった手法がある.これらの手法に対して,デコーディングは観測した行動量に即して縮約するので意味づけが明快だという利点がある.具体的には,回帰とよばれる統計的な手法により,データから神経活動とレバー軌跡とを結ぶ数式をもとめる.回帰にもさまざまあるが,今回は,ガウシアンカーネルを用いたサポートベクター回帰を用いた.この手法の特徴としては,神経活動とレバー軌跡とを結ぶ数式が非線形なものになる点,ノイズによる影響をうけにくい点,最適値が性質よく定まる点があげられる.モデルの非線形性により神経活動とレバー軌跡との関係を柔軟にもとめることで,今回も多くの場合によい予測精度を示した.サポートベクター回帰の弱点はおのおののニューロンの活動とレバー軌跡との関係性がわかりにくい点であるが,今回は,記録したニューロンがレバー軌跡についてもちうる情報を最大限に取り出すことを優先した.
この研究ではさらに,神経デコーディングに対し交差検定という手法を用いることにより予測精度をもとめている.予測精度は,予測されたレバー軌跡と実際に記録されたレバー軌跡とがどのくらい似ていたかということであるが,これは2つの時系列の統計的な依存性を測定することにより定量できる.この研究では,統計的な依存性の尺度として相互情報量を用いた.レバーの軌跡は非常にかたよっているため,相関係数では類似性をつかみそこねる可能性があったためである.
熟練した運動の情報が5a層の神経回路にとどまりやすくなることがわかり,その意味で,5a層の神経回路に“記憶”されることがわかった.つぎの進展の方向性として,新たな疑問を3点ほどあげる.1)このような神経回路の変化はどのくらい持続的なものなのだろうか.半年後に同じ運動をさせたとき,同じように情報を保持できるだろうか.2)このような情報の保持には運動について特異性があるだろうか.ほかの運動を学習させたときには新たな神経回路がつくられるのだろうか.3)この実体としての変化は動物の行動に直接的な影響をもつのであろうか.5a層の神経回路を阻害すると熟練した運動の実行に影響があるのか,あるいは,学習中に阻害すれば運動学習は起こらなくなるのか.これらはこの研究の延長としての課題ではあるが,記憶に関する一般的な問いの変異形であり,今回,みつかった現象が大脳新皮質における自己組織化のひとつの典型として追及すべき研究対象であることを示している.
略歴:2010年 京都大学大学院医学研究科博士課程 修了,2012年より基礎生物学研究所 博士研究員
研究テーマ:大脳皮質が世界を知り,世界を変えるために用いている原理の探求.
正水 芳人(Yoshito Masamizu)
略歴:2008年 京都大学大学院医学研究科博士課程 修了,2014年より基礎生物学研究所 助教.
松崎 政紀(Masanori Matsuzaki)
基礎生物学研究所 教授.
研究室URL:http://www.nibb.ac.jp/circuits/
© 2014 田中康裕・正水芳人・松崎政紀 Licensed under CC 表示 2.1 日本
(基礎生物学研究所 光脳回路研究部門)
email:田中康裕,正水芳人,松崎政紀
DOI: 10.7875/first.author.2014.095
Two distinct layer-specific dynamics of cortical ensembles during learning of a motor task.
Yoshito Masamizu, Yasuhiro R. Tanaka, Yasuyo H. Tanaka, Riichiro Hira, Fuki Ohkubo, Kazuo Kitamura, Yoshikazu Isomura, Takashi Okada, Masanori Matsuzaki
Nature Neuroscience, 17, 987-994 (2014)
要 約
1次運動野は運動出力を5b層から出すが,2つの中間層として2/3層および5a層をもつ.どちらの層もさまざまな脳部位から入力をうけ大脳皮質に投射を出し,それにくわえて,5a層は皮質下への投射をもつ.それゆえ,2/3層および5a層においては運動学習による神経回路の再編成は異なっていると考えられる.筆者らは,レバー引きの運動学習を行っているマウスにおいて1次運動野の2光子Ca2+イメージングを行い,ニューロンの集団および単一のニューロンからのレバー軌跡の予測精度を評価した.2/3層ではニューロンの集団からのレバー軌跡の予測精度は全体としてあまり変わらなかったものの,一部のニューロンは学習期間をとおして高い予測精度を示した.一方,5a層ではニューロンの集団からのレバー軌跡の予測精度は学習をつうじて改善し,皮質下に投射するニューロンを含む33%のニューロンが学習後期における集団による予測に貢献するようになった.2/3層の神経回路は学習をつうじてほかの皮質領野からの情報をうまく統合しながら変化していき,一方で,5a層ではよく学習された運動を表現する学習回路に加担するようになったものと考えられた.
はじめに
大脳の表面をつつむように広がる大脳皮質はさまざまな脳部位との結合をもちながら,それ自体が精緻に組織化された6層の構造をもつ.おのおのの層はおおまかではあるが入力および出力に特異性をもち,この6層の構造,および,その入出力の特異性は哺乳類のあいだで種をこえて,また,機能の異なる皮質領野のあいだでも,ある程度は保存されている.運動を直接に支配していると考えられる1次運動野では,深層にあたる5層において2つの下位構造が知られており,そのひとつである5b層を中心に大脳皮質の主たる運動出力といえる皮質脊髄投射ニューロンが局在する.一方,5a層には皮質脊髄投射ニューロンにくわえ,反対側の線条体へ投射する交差性皮質線条体投射ニューロンが多く局在している1,2).線条体を含む大脳基底核はさまざまな神経変性疾患とかかわり,運動の制御および実行における重要な役割が知られている.一方,皮質の浅層である2/3層は大脳皮質のほかの領野から入力をうけ,それらの領野へ演算の結果を送り返すと考えられている.
近年,2光子顕微鏡技術の進展にともない急速に発展した生体内Ca2+イメージング法により,大脳皮質の2/3層を中心に神経活動の時空間的な特性が調べられるようになった3).この手法の利点は,おおよそ500μm四方に含まれる数百個のニューロンを同時に観察できることである.一方で,レーザーの光学的な特性によるため脳の深い位置での計測には制限があり,これまでの知見は2/3層のものにかたよっていた4-6).今回,筆者らは,2週間にわたる運動学習中に5a層から神経活動を継続的に記録し,学習による特異的な活動変化を検出した.
1.実験および解析の方法
マウスは右前肢によるレバー引きを2週間にわたって学習した(図1).レバーを引くと口の前にあるノズルから水が出るため,これを報酬としてマウスは自らのリズムでレバーを引くようになる5,7).毎日1時間のセッションを行い,成功回数,成功率,成功間隔時間により学習の進行を定量した.成績は1日目から4日目までは急激に上がり,10日程度で高止まりした.以下,1日目から4日目を学習初期,11日目から14日目を学習後期とする.
レバー引きの実行中に左脳の前肢領域においてCa2+イメージングを行った.Ca2+指示体としては蛍光タンパク質GFP,Ca2+応答タンパク質であるカルモジュリン,ミオシン軽鎖キナーゼのC末端側からつくられたGCaMP3 8) を用い,アデノ随伴ウイルスを用いて導入した.Ca2+イメージングは2光子顕微鏡を用いて3~4.5 Hzの周波数で行い,ゆれなどのアーティファクトを取り除いたのち,1視野あたり20~100個のニューロンの輝度の時系列を抽出した.これらの神経活動の時系列と同時に,レバー引きの時系列も記録しておき,神経活動からレバーの軌跡をどのくらい予測できるかにより両者の関連性を定量した.この際,予測精度を調べるため交差検定という手法を用いた9).時系列データを10分割した9/10の期間において神経活動とレバー軌跡との関係を統計的な方法により数式で定め,この定められた数式にしたがい残り1/10の期間の神経活動からレバー軌跡を予測し,予測されたレバー軌跡と実際に記録されたレバー軌跡との類似度を相互情報量という尺度で定量した.この作業を10分割したそれぞれの区間について行い,ニューロンの集団あるいは単一のニューロンからのレバー軌跡の予測精度を決定した.予測精度が高いほど神経活動とレバー軌跡との関係が保たれていたことになり,ニューロンの集団あるいは単一のニューロンがレバー引き運動の情報を保持していたと考えられる.
2.ニューロンの集団における神経活動の変化
学習初期および学習後期において,ニューロンの集団の神経活動からどのくらいレバー軌跡を予測できるか調べた(図2).2/3層ではレバー軌跡の予測精度が上がる視野もあれば下がる視野もありさまざまであったが,5a層では調べた6匹のマウスの7つの視野のすべてで予測精度が上がった.これは,5a層ではレバー引き運動の情報が少なくともセッション内では保持されるようになったことを示唆した.このように,5a層にレバー引き運動の情報が保持されるようになったことは運動学習と関係があるのだろうか.これをみるため,セッションごとのレバー引きの成功回数や成功率と予測精度との関係を調べた.すると,5a層のニューロンの集団によるレバー引き運動の情報の保持がよくなるほど,運動学習を上手にこなしていた.一方,2/3層では5a層のように明瞭な関係は観測されなかった.
3.単一のニューロンにおける神経活動の変化
2/3層および5a層それぞれ7つの視野からのべ約8千個のニューロンの活動を記録したが,そのなかで,2/3層において126個,5a層において187個のニューロンを継続的に記録した(図2).継続的な記録とは,学習初期と学習後期,おのおの最低1日を含む8日間以上にわたり同一のニューロンを記録できたことをさす.これら継続的に記録した単一のニューロンの神経活動からレバー軌跡を予測して予測精度の変化を調べたところ,2/3層では19%のニューロンがレバー引き運動の情報の保持がよくなる一方で,17%のニューロンは情報の保持が悪くなっていた.5a層では33%のニューロンがレバー引き運動の情報の保持がよくなり,情報の保持が悪くなったニューロンは3%のみであった.5a層においてレバー引き運動の情報の保持がよくなったニューロンの活動を調べると,レバー引きのときの神経活動が学習初期に比べ学習後期において高くなっていることがわかった.
単一のニューロンの神経活動の変化は,ニューロンの集団からのレバー軌跡の予測精度にどのような影響をあたえているのだろうか.この問題を調べるため,以下の仮想実験を行った.まず,視野ごとに10個のニューロンを無作為に選び,このニューロンの集団からのレバー軌跡の予測精度をもとめる.つぎに,レバー引き運動の情報の保持がよくなったニューロンがその集団に含まれていた場合,同じ視野から記録されたニューロンに無作為に置き換え,レバー軌跡の予測精度を計算しなおした.すると,運動学習の効果とニューロンの集団からのレバー軌跡の予測精度との関係性がくずれることがわかった.このことから,単一のニューロンとしてレバー引き運動の情報を保持するようになるニューロンが,ニューロンの集団からのレバー軌跡の予測精度と運動学習の効果とを結びつけるのに必須であることがわかった.
ここまで,単一のニューロンからのレバー軌跡の予測精度が絶対値としてどのように変化するかに焦点をあててきたが,つぎに,これらの単一のニューロンが集団のなかでの位置づけをどのように変化させたかを調べた.この問題に取り組むため,継続的に記録された単一のニューロンをレバー軌跡の予測精度によりおのおのの視野においてランクづけし,そのランクの変化を調べた.2/3層では学習初期から学習後期にかけてランクの高いニューロンが比較的保たれていたのに対し,5a層ではランクの高いニューロンの多くが学習初期から学習後期にかけて入れ替わっていることがわかった.したがって,5a層ではレバー引き運動の情報を保持するようになったニューロンが集団のなかでもぬきんでることにより,ニューロンの集団からのレバー軌跡の予測精度を上げるという構図がみえてきた.一方,2/3層ではランクが高く保たれているニューロンが存在し,それらの神経活動はレバー軌跡の予測精度との関連性が学習の全期間をつうじ高かった.これらの神経活動はレバーを動かすときに必須の要素的な運動や付随する感覚と関連しているのかもしれない.
4.皮質下に投射するニューロンにおける神経活動の変化
5a層は皮質下に投射するニューロンを多くもつ.なかでも,反対側の線条体に投射する交差性皮質線条体投射ニューロンおよび皮質脊髄投射ニューロンに着目し,14日間の学習期間においてどのような神経活動の変化を示すか調べた.交差性皮質線条体投射ニューロンおよび皮質脊髄投射ニューロンのいずれも30%程度が予測精度を向上させ,5%程度が予測精度を低下させた.ところが,交差性皮質線条体投射ニューロンでは学習初期から学習後期にかけて高いランクを維持しつづけるニューロンが20%程度だったのに対し,皮質脊髄投射ニューロンでは学習初期にランクの高いニューロンのすべてが学習後期にも高いランクを示した.これらの結果から,交差性皮質線条体投射ニューロンと皮質脊髄投射ニューロンのどちらも予測精度を変化させるニューロンの割合は5a層の全体と同じ程度であることがわかった.それにくわえ,新たにレバー引きを表現するようになった神経回路の一部として交差性皮質線条体投射ニューロンが組み込まれる可能性が示唆された.皮質脊髄投射ニューロンのなかで学習初期から学習後期までレバー引き運動の情報を保持できるニューロンは,レバー引き運動に必須の動きをつかさどるなど,レバー引き運動ととくに関連の高いニューロンであると考えられた.
5.用いた解析法のねらい
この研究においては,ニューロンの集団あるいは単一のニューロンがレバー引き運動を表現しているかどうかに関して,神経デコーディング10) という手法を用いてアプローチした.これは,多数のニューロンの神経活動からレバー引き運動をどれだけ説明できるかを調べたものである.データ解析の視点からは,多次元の神経活動から1次元の時系列を予測することにより,把握しづらい多次元の情報の次元を減らして可視化したともいえる.同じように,多次元の情報を縮約して表現する方法には主成分分析や因子分析といった手法がある.これらの手法に対して,デコーディングは観測した行動量に即して縮約するので意味づけが明快だという利点がある.具体的には,回帰とよばれる統計的な手法により,データから神経活動とレバー軌跡とを結ぶ数式をもとめる.回帰にもさまざまあるが,今回は,ガウシアンカーネルを用いたサポートベクター回帰を用いた.この手法の特徴としては,神経活動とレバー軌跡とを結ぶ数式が非線形なものになる点,ノイズによる影響をうけにくい点,最適値が性質よく定まる点があげられる.モデルの非線形性により神経活動とレバー軌跡との関係を柔軟にもとめることで,今回も多くの場合によい予測精度を示した.サポートベクター回帰の弱点はおのおののニューロンの活動とレバー軌跡との関係性がわかりにくい点であるが,今回は,記録したニューロンがレバー軌跡についてもちうる情報を最大限に取り出すことを優先した.
この研究ではさらに,神経デコーディングに対し交差検定という手法を用いることにより予測精度をもとめている.予測精度は,予測されたレバー軌跡と実際に記録されたレバー軌跡とがどのくらい似ていたかということであるが,これは2つの時系列の統計的な依存性を測定することにより定量できる.この研究では,統計的な依存性の尺度として相互情報量を用いた.レバーの軌跡は非常にかたよっているため,相関係数では類似性をつかみそこねる可能性があったためである.
おわりに
熟練した運動の情報が5a層の神経回路にとどまりやすくなることがわかり,その意味で,5a層の神経回路に“記憶”されることがわかった.つぎの進展の方向性として,新たな疑問を3点ほどあげる.1)このような神経回路の変化はどのくらい持続的なものなのだろうか.半年後に同じ運動をさせたとき,同じように情報を保持できるだろうか.2)このような情報の保持には運動について特異性があるだろうか.ほかの運動を学習させたときには新たな神経回路がつくられるのだろうか.3)この実体としての変化は動物の行動に直接的な影響をもつのであろうか.5a層の神経回路を阻害すると熟練した運動の実行に影響があるのか,あるいは,学習中に阻害すれば運動学習は起こらなくなるのか.これらはこの研究の延長としての課題ではあるが,記憶に関する一般的な問いの変異形であり,今回,みつかった現象が大脳新皮質における自己組織化のひとつの典型として追及すべき研究対象であることを示している.
文 献
- Morishima, M., & Kawaguchi, Y.: Recurrent connection patterns of corticostriatal pyramidal cells in frontal cortex. J. Neurosci., 26, 4394-4405 (2006)[PubMed]
- Kiritani, T., Wickersham, I. R., Seung, H. S. et al.: Hierarchical connectivity and connection-specific dynamics in the corticospinal-corticostriatal microcircuit in mouse motor cortex. J. Neurosci., 32, 4992-5001 (2012)[PubMed]
- Ohki, K., Chung, S., Ch’ng, Y. H. et al.: Functional imaging with cellular resolution reveals precise micro-architecture in visual cortex. Nature, 433, 597-603 (2005)[PubMed]
- Huber, D., Gutnisky, D. A., Peron, S. et al.: Multiple dynamic representations in the motor cortex during sensorimotor learning. Nature, 484, 473-478 (2012)[PubMed]
- Hira, R., Ohkubo, F., Ozawa, K. et al.: Spatiotemporal dynamics of functional clusters of neurons in the mouse motor cortex during a voluntary movement. J. Neurosci., 33, 1377-1390 (2013)[PubMed]
- Peters, A. J., Chen, S. X. & Komiyama, T.: Emergence of reproducible spatiotemporal activity during motor learning. Nature, 510, 263-267 (2014)[PubMed]
- Isomura, Y., Harukuni, R., Takekawa, T. et al.: Microcircuitry coordination of cortical motor information in self-initiation of voluntary movements. Nat. Neurosci., 12, 1586-1593 (2009)[PubMed]
- Tian, L., Hires, S. A., Mao, T. et al.: Imaging neural activity in worms, flies and mice with improved GCaMP calcium indicators. Nat. Methods, 6, 875-881 (2009)[PubMed]
- Bishop, C. M.: Pattern Recognition and Machine Learning. Springer, New York (2006)
- Wessberg, J., Stambaugh, C. R., Kralik, J. D. et al.: Real-time prediction of hand trajectory by ensembles of cortical neurons in primates. Nature, 408, 361-365 (2000)[PubMed]
著者プロフィール
略歴:2010年 京都大学大学院医学研究科博士課程 修了,2012年より基礎生物学研究所 博士研究員
研究テーマ:大脳皮質が世界を知り,世界を変えるために用いている原理の探求.
正水 芳人(Yoshito Masamizu)
略歴:2008年 京都大学大学院医学研究科博士課程 修了,2014年より基礎生物学研究所 助教.
松崎 政紀(Masanori Matsuzaki)
基礎生物学研究所 教授.
研究室URL:http://www.nibb.ac.jp/circuits/
© 2014 田中康裕・正水芳人・松崎政紀 Licensed under CC 表示 2.1 日本
