連合学習における嗅内皮質と海馬との協調的な神経活動の増強
五十嵐 啓
(ノルウェーNorwegian University of Science and Technology,Kavli Institute for Systems Neuroscience)
email:五十嵐 啓
DOI: 10.7875/first.author.2014.075
Coordination of entorhinal-hippocampal ensemble activity during associative learning.
Kei M. Igarashi, Li Lu, Laura L. Colgin, May-Britt Moser, Edvard I. Moser
Nature, 510, 143-147 (2014)
記憶,とくに陳述的な記憶の機能は,脳皮質と海馬とのあいだの情報を橋渡しする嗅内皮質により担われている.最近,嗅内皮質と海馬の神経回路がガンマ波長帯の神経振動活動により相互作用していることが示唆されたが,記憶が形成される過程との関係は不明であった.この研究では,ラットが匂い-場所連合学習を行う際に嗅内皮質および海馬から同時記録することにより,学習中に嗅内皮質と海馬の神経振動活動の同期が増強され,この同期の増強が個々の細胞のスパイク活動の集合と相関していることを明らかにした.この結果は,神経振動活動が匂い-場所連合学習中に発達する海馬と嗅内皮質の神経回路の情報表現を統合させる機能をもつことを示唆した.
脳のさまざまな領域において脳波を記録すると,シータ波長帯(6~12 Hz)あるいはガンマ波長帯(30~100 Hz)などの神経振動活動(神経オシレーション活動)の観察されることが知られている1-3).これまでの研究から,これらの神経振動活動の同期が特化した機能をもつ脳領域を統合させる役割をもつことが示唆されてきた.このような脳領域の統合を必要とすると考えられる脳機能のひとつに陳述的な記憶がある4).陳述的な記憶の機能は脳皮質と海馬とのあいだの情報を橋渡しする嗅内皮質により担われているが,陳述記憶の記銘および想起の過程において,嗅内皮質と海馬の神経回路はガンマ波長帯の神経振動活動により相互作用していると考えられている.実際に,覚醒中のげっ歯類ではこの波長帯の活動が多く観察されており,以前に,筆者らの研究室は,学習後のラットの嗅内皮質および海馬から同時記録を行うと同期したシータ波およびガンマ波のみられることを報告した5).しかしながら,これらの実験は動物が学習をすませたあとに行われたものであり,領域のあいだの神経振動活動の同期と記憶の形成との関係は不明であった.
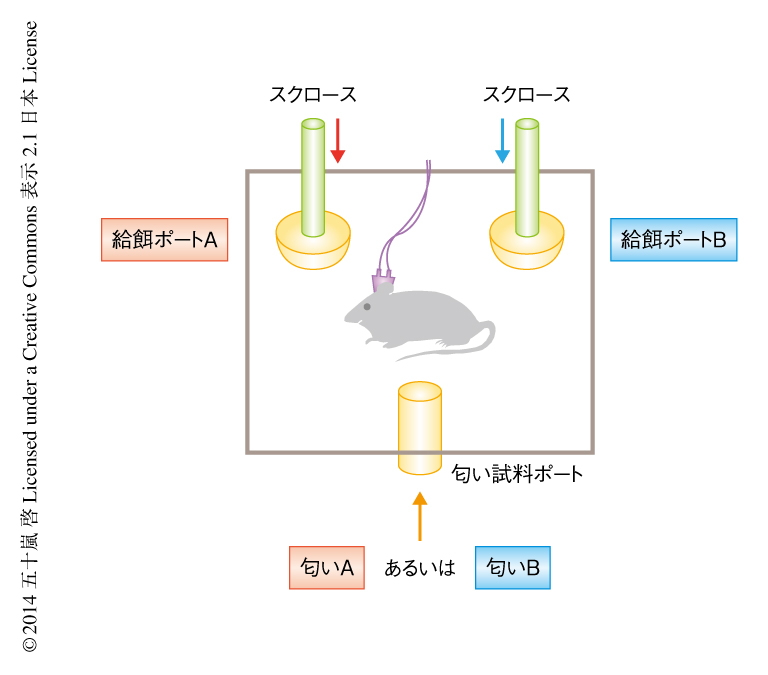
複数の記録電極をラットの海馬CA1領域および嗅内皮質(図1)に留置し,スパイク活動および局所電場電位(local field potential:LFP)の記録を行った.海馬に依存的な学習である匂い-場所連合課題6)(図2)を3週間で85%以上の正答率が得られるまでトレーニングし,その期間の神経活動を記録した.匂い-場所連合課題は,1)匂い試料ポートに鼻先を入れ,匂いAあるいは匂いBを嗅ぐ,2)匂いを嗅ぎおわり,ポートから鼻先を出して2つある給餌ポートAあるいは給餌ポートBのうちいずれかに移動する,3)給餌ポートに鼻先を入れ,それが正解であればスクロースの餌が得られる,という3段階の行動タスクである.匂いAと匂いBはランダムに出現するが,匂いAが呈示されたときは給餌ポートAを,匂いBが呈示されたにときは給餌ポートBを選択しなければ餌は得られない.
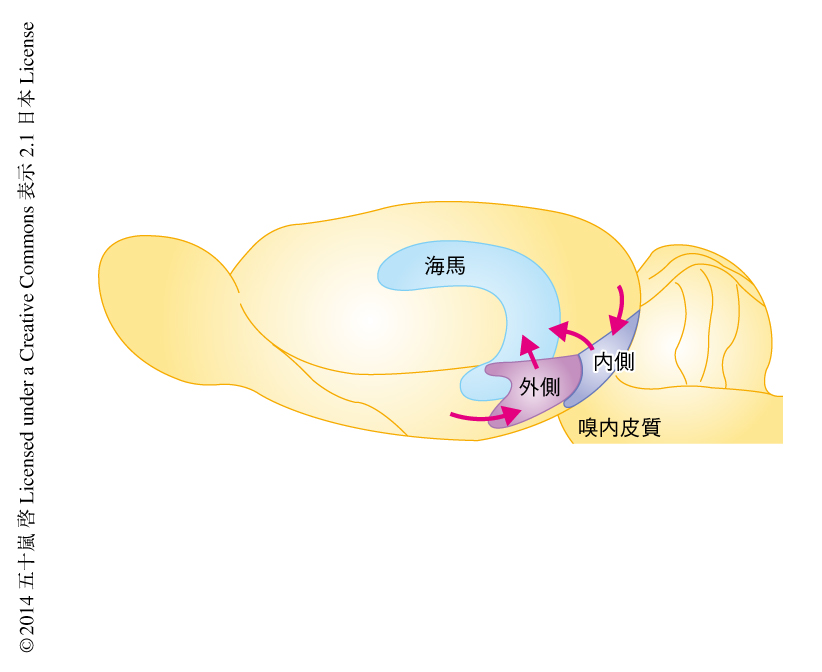
連合学習をおえたラットの海馬から記録を行った.その結果,海馬CA1領域では匂いを嗅いでいるあいだ20~40 Hz波長帯の強い神経振動活動の観察されることが明らかになった.この神経振動活動は海馬CA1遠位部にて強く海馬CA1近位部ではあまり観察されなかった.解剖学的には,海馬CA1遠位部は嗅内皮質の外側部から,海馬CA1近位部は嗅内皮質の内側部から,直接の投射をうけることが知られている7)(図1).そこで,嗅内皮質の外側部とおよび内側部からそれぞれ記録を行った.その結果,匂いを嗅いでいるあいだ,嗅内皮質の外側部にのみ20~40 Hz波長帯の神経振動活動が観察された.さらに,海馬CA1遠位部および嗅内皮質の外側部から同時記録を行うと,これらの領域で観察される20~40 Hz波長帯の神経振動活動が同期していることが明らかになった.
この同期は海馬CA1遠位部と嗅内皮質の外側部とのあいだで特異的なのだろうか? これを明らかにするため,海馬CA1遠位部あるいは海馬CA1近位部,および,嗅内皮質の外側部あるいは内側部の組合せで同時記録を行ったところ,同期は海馬CA1遠位部と嗅内皮質の外側部とのあいだのみで観察された.よって,匂い-場所連合学習においては,20~40 Hz波長帯の神経振動活動を介した機能的な結合が海馬CA1遠位部と嗅内皮質の外側部とのあいだでみられることが示された.
この研究の主目的である,神経振動活動の同期が学習中に変化するかどうかという命題を検証した.学習をしていないラットの海馬CA1遠位部および嗅内皮質の外側部に記録電極を留置し,課題を学習する際の神経活動の変化を記録した.その結果,学習の開始直後より20~40 Hz波長帯の神経振動活動それ自体は海馬CA1遠位部および嗅内皮質の外側部それぞれにおいて観察されたものの,それらは同期していなかった.しかし,学習の進行にともない20~40 Hz波長帯の神経振動活動の同期は増強され,学習をおえた段階で同期はもっとも増強されていた.
20~40 Hz波長帯の神経振動活動の同期はラットの正答行動に必要なのだろうか? これを明らかにするため,エラー試行における神経活動について検証した.ラットは85%の正答率で行動するが,残りの15%ではまちがった給餌ポートを選択してしまう.このようなエラー試行においては20~40 Hz波長帯の神経振動活動の同期は減弱していることが明らかになった.すなわち,学習中に海馬と嗅内皮質とのあいだの機能的な結合が20~40 Hz波長帯の神経振動活動の同期により増強され,この同期がラットの正答行動に必要であることが示された.
ここまでの結果は,脳波として観察される複数の細胞の集合活動の結果であった.細胞の集合活動としてみられる脳波の振動と類似した時間的な構造が,個々の細胞においてもみられるだろうか? また,個々の細胞も20~40 Hz波長帯の神経振動活動の同期のような学習中の変化を示すのだろうか? 匂いを嗅いでいるあいだ1 Hz以上のスパイク発火の頻度を示す細胞を海馬CA1遠位部および嗅内皮質の外側部それぞれから60~80個を記録し,この細胞におけるスパイク活動の学習中の変化をおった.
これらのスパイク列の時間的な構造を検討した.スパイク列が神経振動活動と同様の時間的な構造をもつかどうかを検証するため,20~40 Hz波長帯の神経振動活動の位相とスパイク発火のタイミングとの相互関係を解析したところ,海馬CA1遠位部の細胞および嗅内皮質の外側部の細胞のスパイク列は,学習にともない20~40 Hz波長帯の神経振動活動の特定の位相において発火する傾向の強まる(位相がロックする)ことが明らかになった.このことは,海馬CA1遠位部および嗅内皮質の外側部のいずれにおいても,20~40 Hz波長帯の神経振動活動が個々の細胞のスパイク活動の集合として形成されていることを示唆した.
細胞のスパイク活動が表現する情報について検討した.海馬CA1遠位部および嗅内皮質の外側部の細胞の一部は,学習後に匂いAと匂いBとのあいだで異なるスパイク発火の頻度を示す.このような匂いに選択的な細胞の割合を学習の前後で比較すると,学習前と比べて学習後は有意に割合が上昇していた.このような変化は海馬CA1遠位部よりも嗅内皮質の外側部においてより顕著に観察された.さらに,匂いに選択的な細胞における選択的なスパイク発火は,エラー試行において減弱していた.この結果から,神経振動活動の同期と同様に,海馬CA1遠位部および嗅内皮質の外側部の細胞における匂いに選択的なスパイク発火がラットの正答行動に必要であることが示された.
ここまで,ラットの行動の正答率の上昇にともない,20~40 Hz波長帯の神経振動活動の同期,および,海馬CA1遠位部および嗅内皮質の外側部の細胞における匂いに選択的なスパイク発火が,それぞれ増強されることが明らかになった.では,これらの現象のあいだの相互関係はどのようになっているのだろうか? いずれかの部位の神経活動がさきに変化して,ほかの部位の活動を促進しているのだろうか? これを明らかにするため,動物の学習期間を未習熟から習熟まで5つの段階に分け,行動の正答率,20~40 Hz波長帯の神経振動活動の同期,海馬CA1遠位部における匂いに選択的なスパイク発火,嗅内皮質の外側部における匂いに選択的なスパイク発火,の増強のパターンを比較した.その結果,いずれのあいだにも相関がみられた.しかし,行動の正答率,20~40 Hz波長帯の神経振動活動の同期,嗅内皮質の外側部における匂いに選択的なスパイク発火は,類似した上昇曲線を示したが,海馬CA1遠位部における匂いに選択的なスパイク発火は,それよりも立ち上がりの遅れた上昇曲線を示した.この結果は,嗅内皮質におけるスパイク発火が海馬CA1領域の変化を誘導している可能性を示唆した.
以上の結果から,20~40 Hz波長帯の神経振動活動の同期が,匂い-場所連合学習中に発達する海馬と嗅内皮質の神経回路の情報表現を統合させる機能をもつことが示唆された.これまで,神経振動活動が脳の別個の機能モジュールを統合させる可能性は示唆されてきたが3),この統合機能が学習に関与しているかどうかは不明だった.この研究は,神経振動活動が脳の学習,および,海馬と嗅内皮質との機能的な結合の増強に関与していることを示唆したはじめての結果であった.20~40 Hz波長帯の神経振動活動の同期により,嗅内皮質の外側部の細胞のシナプス前部と海馬CA1遠位部の細胞のシナプス後部のスパイク発火が時間的に近接したパターンを示すことで,スパイク発火のタイミングに依存したシナプスの可塑性などの機構により,海馬と嗅内皮質とのあいだ機能的な結合の増強が実現されるものと考えられる.20~40 Hz波長帯の神経振動活動は脳において広くみられる現象である8).とくに,嗅球や梨状皮質などの嗅覚野では強い20~40 Hz波長帯の神経振動活動が観察され,これらの嗅覚野の機能的な結合を促進すると予想されている9).嗅内皮質の外側部はこれらの嗅覚野からの直接の投射をうけて海馬へと嗅覚シグナルを送り込む部位であり10),20~40 Hz波長帯の神経振動活動が嗅覚野から海馬の広い範囲にわたる機能的な結合を促進している可能性もある.われわれは日常生活において,ある匂いを嗅いだ瞬間に過去の場面を鮮やかに思い出すことがあるが,このような匂いによる記憶の想起には,われわれの脳で生じる20~40 Hz波長帯の神経振動活動が関与しているのかもしれない.
略歴:2007年 東京大学大学院医学系研究科 修了,同年 同 研究員,2009年 ノルウェーNorwegian University of Science and Technology博士研究員を経て,2013年より同 リサーチアソシエイト.
研究テーマ:脳の機能統合を実現する神経機構.
関心事:記憶や対象物の認識といった脳の高次機能は,脳の個々の領域の機能により担われているわけではなく,複数の領域の機能的な結合により担われるものと考えられる.このような機能的な結合を実現する神経機構を,システム生物学的な発想で明らかにしていきたい.
© 2014 五十嵐 啓 Licensed under CC 表示 2.1 日本
(ノルウェーNorwegian University of Science and Technology,Kavli Institute for Systems Neuroscience)
email:五十嵐 啓
DOI: 10.7875/first.author.2014.075
Coordination of entorhinal-hippocampal ensemble activity during associative learning.
Kei M. Igarashi, Li Lu, Laura L. Colgin, May-Britt Moser, Edvard I. Moser
Nature, 510, 143-147 (2014)
要 約
記憶,とくに陳述的な記憶の機能は,脳皮質と海馬とのあいだの情報を橋渡しする嗅内皮質により担われている.最近,嗅内皮質と海馬の神経回路がガンマ波長帯の神経振動活動により相互作用していることが示唆されたが,記憶が形成される過程との関係は不明であった.この研究では,ラットが匂い-場所連合学習を行う際に嗅内皮質および海馬から同時記録することにより,学習中に嗅内皮質と海馬の神経振動活動の同期が増強され,この同期の増強が個々の細胞のスパイク活動の集合と相関していることを明らかにした.この結果は,神経振動活動が匂い-場所連合学習中に発達する海馬と嗅内皮質の神経回路の情報表現を統合させる機能をもつことを示唆した.
はじめに
脳のさまざまな領域において脳波を記録すると,シータ波長帯(6~12 Hz)あるいはガンマ波長帯(30~100 Hz)などの神経振動活動(神経オシレーション活動)の観察されることが知られている1-3).これまでの研究から,これらの神経振動活動の同期が特化した機能をもつ脳領域を統合させる役割をもつことが示唆されてきた.このような脳領域の統合を必要とすると考えられる脳機能のひとつに陳述的な記憶がある4).陳述的な記憶の機能は脳皮質と海馬とのあいだの情報を橋渡しする嗅内皮質により担われているが,陳述記憶の記銘および想起の過程において,嗅内皮質と海馬の神経回路はガンマ波長帯の神経振動活動により相互作用していると考えられている.実際に,覚醒中のげっ歯類ではこの波長帯の活動が多く観察されており,以前に,筆者らの研究室は,学習後のラットの嗅内皮質および海馬から同時記録を行うと同期したシータ波およびガンマ波のみられることを報告した5).しかしながら,これらの実験は動物が学習をすませたあとに行われたものであり,領域のあいだの神経振動活動の同期と記憶の形成との関係は不明であった.
1.匂いを嗅いでいるあいだの20~40 Hz波長帯の神経振動活動の同期
複数の記録電極をラットの海馬CA1領域および嗅内皮質(図1)に留置し,スパイク活動および局所電場電位(local field potential:LFP)の記録を行った.海馬に依存的な学習である匂い-場所連合課題6)(図2)を3週間で85%以上の正答率が得られるまでトレーニングし,その期間の神経活動を記録した.匂い-場所連合課題は,1)匂い試料ポートに鼻先を入れ,匂いAあるいは匂いBを嗅ぐ,2)匂いを嗅ぎおわり,ポートから鼻先を出して2つある給餌ポートAあるいは給餌ポートBのうちいずれかに移動する,3)給餌ポートに鼻先を入れ,それが正解であればスクロースの餌が得られる,という3段階の行動タスクである.匂いAと匂いBはランダムに出現するが,匂いAが呈示されたときは給餌ポートAを,匂いBが呈示されたにときは給餌ポートBを選択しなければ餌は得られない.
連合学習をおえたラットの海馬から記録を行った.その結果,海馬CA1領域では匂いを嗅いでいるあいだ20~40 Hz波長帯の強い神経振動活動の観察されることが明らかになった.この神経振動活動は海馬CA1遠位部にて強く海馬CA1近位部ではあまり観察されなかった.解剖学的には,海馬CA1遠位部は嗅内皮質の外側部から,海馬CA1近位部は嗅内皮質の内側部から,直接の投射をうけることが知られている7)(図1).そこで,嗅内皮質の外側部とおよび内側部からそれぞれ記録を行った.その結果,匂いを嗅いでいるあいだ,嗅内皮質の外側部にのみ20~40 Hz波長帯の神経振動活動が観察された.さらに,海馬CA1遠位部および嗅内皮質の外側部から同時記録を行うと,これらの領域で観察される20~40 Hz波長帯の神経振動活動が同期していることが明らかになった.
この同期は海馬CA1遠位部と嗅内皮質の外側部とのあいだで特異的なのだろうか? これを明らかにするため,海馬CA1遠位部あるいは海馬CA1近位部,および,嗅内皮質の外側部あるいは内側部の組合せで同時記録を行ったところ,同期は海馬CA1遠位部と嗅内皮質の外側部とのあいだのみで観察された.よって,匂い-場所連合学習においては,20~40 Hz波長帯の神経振動活動を介した機能的な結合が海馬CA1遠位部と嗅内皮質の外側部とのあいだでみられることが示された.
2.匂い-場所連合学習中に20~40 Hz波長帯の神経振動活動の同期は増強される
この研究の主目的である,神経振動活動の同期が学習中に変化するかどうかという命題を検証した.学習をしていないラットの海馬CA1遠位部および嗅内皮質の外側部に記録電極を留置し,課題を学習する際の神経活動の変化を記録した.その結果,学習の開始直後より20~40 Hz波長帯の神経振動活動それ自体は海馬CA1遠位部および嗅内皮質の外側部それぞれにおいて観察されたものの,それらは同期していなかった.しかし,学習の進行にともない20~40 Hz波長帯の神経振動活動の同期は増強され,学習をおえた段階で同期はもっとも増強されていた.
20~40 Hz波長帯の神経振動活動の同期はラットの正答行動に必要なのだろうか? これを明らかにするため,エラー試行における神経活動について検証した.ラットは85%の正答率で行動するが,残りの15%ではまちがった給餌ポートを選択してしまう.このようなエラー試行においては20~40 Hz波長帯の神経振動活動の同期は減弱していることが明らかになった.すなわち,学習中に海馬と嗅内皮質とのあいだの機能的な結合が20~40 Hz波長帯の神経振動活動の同期により増強され,この同期がラットの正答行動に必要であることが示された.
3.学習中の匂いに特異的なスパイク発火の変化
ここまでの結果は,脳波として観察される複数の細胞の集合活動の結果であった.細胞の集合活動としてみられる脳波の振動と類似した時間的な構造が,個々の細胞においてもみられるだろうか? また,個々の細胞も20~40 Hz波長帯の神経振動活動の同期のような学習中の変化を示すのだろうか? 匂いを嗅いでいるあいだ1 Hz以上のスパイク発火の頻度を示す細胞を海馬CA1遠位部および嗅内皮質の外側部それぞれから60~80個を記録し,この細胞におけるスパイク活動の学習中の変化をおった.
これらのスパイク列の時間的な構造を検討した.スパイク列が神経振動活動と同様の時間的な構造をもつかどうかを検証するため,20~40 Hz波長帯の神経振動活動の位相とスパイク発火のタイミングとの相互関係を解析したところ,海馬CA1遠位部の細胞および嗅内皮質の外側部の細胞のスパイク列は,学習にともない20~40 Hz波長帯の神経振動活動の特定の位相において発火する傾向の強まる(位相がロックする)ことが明らかになった.このことは,海馬CA1遠位部および嗅内皮質の外側部のいずれにおいても,20~40 Hz波長帯の神経振動活動が個々の細胞のスパイク活動の集合として形成されていることを示唆した.
細胞のスパイク活動が表現する情報について検討した.海馬CA1遠位部および嗅内皮質の外側部の細胞の一部は,学習後に匂いAと匂いBとのあいだで異なるスパイク発火の頻度を示す.このような匂いに選択的な細胞の割合を学習の前後で比較すると,学習前と比べて学習後は有意に割合が上昇していた.このような変化は海馬CA1遠位部よりも嗅内皮質の外側部においてより顕著に観察された.さらに,匂いに選択的な細胞における選択的なスパイク発火は,エラー試行において減弱していた.この結果から,神経振動活動の同期と同様に,海馬CA1遠位部および嗅内皮質の外側部の細胞における匂いに選択的なスパイク発火がラットの正答行動に必要であることが示された.
4.神経活動と行動との相互関係
ここまで,ラットの行動の正答率の上昇にともない,20~40 Hz波長帯の神経振動活動の同期,および,海馬CA1遠位部および嗅内皮質の外側部の細胞における匂いに選択的なスパイク発火が,それぞれ増強されることが明らかになった.では,これらの現象のあいだの相互関係はどのようになっているのだろうか? いずれかの部位の神経活動がさきに変化して,ほかの部位の活動を促進しているのだろうか? これを明らかにするため,動物の学習期間を未習熟から習熟まで5つの段階に分け,行動の正答率,20~40 Hz波長帯の神経振動活動の同期,海馬CA1遠位部における匂いに選択的なスパイク発火,嗅内皮質の外側部における匂いに選択的なスパイク発火,の増強のパターンを比較した.その結果,いずれのあいだにも相関がみられた.しかし,行動の正答率,20~40 Hz波長帯の神経振動活動の同期,嗅内皮質の外側部における匂いに選択的なスパイク発火は,類似した上昇曲線を示したが,海馬CA1遠位部における匂いに選択的なスパイク発火は,それよりも立ち上がりの遅れた上昇曲線を示した.この結果は,嗅内皮質におけるスパイク発火が海馬CA1領域の変化を誘導している可能性を示唆した.
おわりに
以上の結果から,20~40 Hz波長帯の神経振動活動の同期が,匂い-場所連合学習中に発達する海馬と嗅内皮質の神経回路の情報表現を統合させる機能をもつことが示唆された.これまで,神経振動活動が脳の別個の機能モジュールを統合させる可能性は示唆されてきたが3),この統合機能が学習に関与しているかどうかは不明だった.この研究は,神経振動活動が脳の学習,および,海馬と嗅内皮質との機能的な結合の増強に関与していることを示唆したはじめての結果であった.20~40 Hz波長帯の神経振動活動の同期により,嗅内皮質の外側部の細胞のシナプス前部と海馬CA1遠位部の細胞のシナプス後部のスパイク発火が時間的に近接したパターンを示すことで,スパイク発火のタイミングに依存したシナプスの可塑性などの機構により,海馬と嗅内皮質とのあいだ機能的な結合の増強が実現されるものと考えられる.20~40 Hz波長帯の神経振動活動は脳において広くみられる現象である8).とくに,嗅球や梨状皮質などの嗅覚野では強い20~40 Hz波長帯の神経振動活動が観察され,これらの嗅覚野の機能的な結合を促進すると予想されている9).嗅内皮質の外側部はこれらの嗅覚野からの直接の投射をうけて海馬へと嗅覚シグナルを送り込む部位であり10),20~40 Hz波長帯の神経振動活動が嗅覚野から海馬の広い範囲にわたる機能的な結合を促進している可能性もある.われわれは日常生活において,ある匂いを嗅いだ瞬間に過去の場面を鮮やかに思い出すことがあるが,このような匂いによる記憶の想起には,われわれの脳で生じる20~40 Hz波長帯の神経振動活動が関与しているのかもしれない.
文 献
- Freeman, W. J.: Spatial properties of an EEG event in the olfactory bulb and cortex. Electroencephalogr. Clin. Neurophysiol., 44, 586-605 (1978)[PubMed]
- Gray, C. M., Konig, P., Engel, A. K. et al.: Oscillatory responses in cat visual cortex exhibit inter-columnar synchronization which reflects global stimulus properties. Nature, 338, 334-337 (1989)[PubMed]
- Singer, W.: Synchronization of cortical activity and its putative role in information processing and learning. Annu. Rev. Physiol., 55, 349-374 (1993)[PubMed]
- Buzsaki, G. & Moser, E. I.: Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat. Neurosci., 16, 130-138 (2013)[PubMed]
- Colgin, L. L., Denninger, T., Fyhn, M. et al.: Frequency of gamma oscillations routes flow of information in the hippocampus. Nature, 462, 353-357 (2009)[PubMed]
- Day, M., Langston, R. & Morris, R. G.: Glutamate-receptor-mediated encoding and retrieval of paired-associate learning. Nature, 424, 205-209 (2003)[PubMed]
- Tamamaki, N. & Nojyo, Y.: Preservation of topography in the connections between the subiculum, field CA1, and the entorhinal cortex in rats. J. Comp. Neurol., 353, 379-390 (1995)[PubMed]
- Engel, A. K. & Fries, P.: Beta-band oscillations: signalling the status quo? Curr. Opin. Neurobiol., 20, 156-165 (2010)[PubMed]
- Ravel, N., Chabaud, P., Martin, C. et al.: Olfactory learning modifies the expression of odour-induced oscillatory responses in the gamma (60-90 Hz) and beta (15-40 Hz) bands in the rat olfactory bulb. Eur. J. Neurosci., 17, 350-358 (2003)[PubMed]
- Igarashi, K. M., Ieki, N., An, M. et al.: Parallel mitral and tufted cell pathways route distinct odor information to different targets in the olfactory cortex. J. Neurosci., 32, 7970-7985 (2012)[PubMed]
著者プロフィール
略歴:2007年 東京大学大学院医学系研究科 修了,同年 同 研究員,2009年 ノルウェーNorwegian University of Science and Technology博士研究員を経て,2013年より同 リサーチアソシエイト.
研究テーマ:脳の機能統合を実現する神経機構.
関心事:記憶や対象物の認識といった脳の高次機能は,脳の個々の領域の機能により担われているわけではなく,複数の領域の機能的な結合により担われるものと考えられる.このような機能的な結合を実現する神経機構を,システム生物学的な発想で明らかにしていきたい.
© 2014 五十嵐 啓 Licensed under CC 表示 2.1 日本
