ゼブラフィッシュにおいて視覚運動刺激の情報処理をつかさどる神経機構
久保 郁
(ドイツMax Planck Institute of Neurobiology,Department Genes-Circuits-Behavior)
email:久保 郁
DOI: 10.7875/first.author.2014.051
Functional architecture of an optic flow-responsive area that drives horizontal eye movements in zebrafish.
Fumi Kubo, Bastian Hablitzel, Marco Dal Maschio, Wolfgang Driever, Herwig Baier, Aristides B. Arrenberg
Neuron, 81, 1344-1359 (2014)
多くの動物は外界の視野全体が大きく動くような視覚刺激を経験すると,それを打ち消すようにして補完的に眼や身体を動かす反応を示す.このような視覚刺激は,視野が回転する特性をもつ場合と,視野が平行移動する特性をもつ場合の2つに大別される.これら2つの視覚特性はそれぞれ異なる運動反応をひき起こすことから,脳において互いに明確に区別される必要がある.この論文では,光遺伝学的な手法とCa2+イメージング法とを組み合わせることにより,このような視覚情報の処理にかかわる神経回路を体系的に調べた.そして,視蓋前野に存在する何千ものニューロンの活動を記録することにより,視蓋前野のニューロンは高い方向選択性をもち,単眼性あるいは両眼性の入力をうけていて,回転する視覚刺激と平行移動する視覚刺激とを区別しうるような構成要素を含んでいることが明らかにされた.筆者らのデータから,視覚刺激によりひき起こされる補完的な運動反応を駆動するのに適した神経回路の仮説が提唱された.
われわれが道を歩いているとき,あるいは,電車に乗っているとき,外界の風景は進行方向とは逆の方向へと動いているように知覚される.このような視覚情報は“光学的流動”(optic flow)とよばれ,自分の身体が外界に対し相対的に移動することにより像が網膜のうえを流れるようにして移動することを意味する.われわれは光学的流動を感知すると,光学的流動と同じ方向に眼あるいは身体を動かすことにより,生じた視覚のエラーを最小限にとどめることができる.この反応は移動における視覚情報の処理を助けるのにきわめて効果的であり,多くの脊椎動物において広く保存されている1).
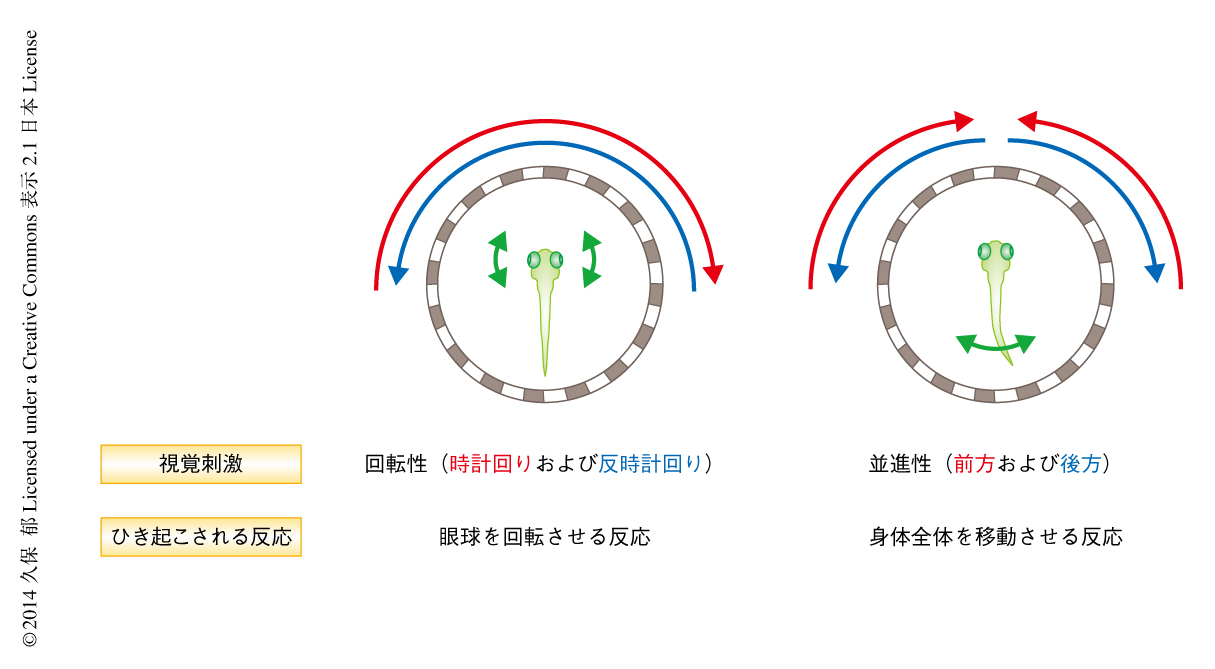
とりわけ魚類では,光学的流動が回転する特性をもつ場合には眼球を回転させる反応が誘導され,一方,光学的流動が平行移動(並進運動)する特性をもつ場合には前方あるいは後方に動くことにより身体全体を移動させる反応が誘導されることが知られている2).これらの適切な運動パターンを生み出すためには,回転性の視覚特性と並進性の視覚特性とが脳において明確に区別される必要がある(図1).個体が回転性あるいは並進性の視覚情報を区別しうるための戦略のひとつとして,両方の眼からの入力をなんらかの方法で比較し統合するという機構が考えられるが,このような情報処理あるいは演算処理が神経回路のどこでどのように行われているかについてのコンセンサスは得られていなかった.この研究では,光学的な手法による神経活動の観察および操作のしやすいゼブラフィッシュをモデルとして3,4),網膜の出力ニューロンからの直接の投射をうける視蓋前野(pretectum)に注目した.光遺伝学的な手法を用いてゼブラフィッシュにおける視蓋前野を機能的に同定したうえで,この領域における神経活動を2光子Ca2+イメージング法により単一のニューロンのレベルで記録し,視蓋前野における両眼からの視覚情報の処理の機構について解析した.
ゼブラフィッシュにおいて視蓋前野が眼球を回転させる反応に関与するかどうかを調べるため,光遺伝学的な手法を用いて脳の限局した領域を光刺激した.光に依存して神経活動を活性化する効果をもつチャネルロドプシン2を神経系の全体で発現するトランスジェニックゼブラフィッシュの稚魚を用いて,光ファイバーにより片側の視蓋前野を光刺激すると両方の眼球が回転する反応が観察された.この眼球の反応は視覚刺激により誘導される眼球を回転させる反応とよく似た特性をもっていた.逆に,光に依存して神経活動を抑制する効果をもつハロロドプシンを発現させたトランスジェニックゼブラフィッシュの稚魚において視蓋前野を光刺激すると,視覚刺激によりひき起こされる眼球を回転させる反応がいちじるしく阻害された.以上から,視蓋前野は眼球を回転させる反応をひき起こすのに必要かつ十分であることが示された.
視蓋前野のニューロンは視覚情報に対してどのように反応するのだろうか? ゼブラフィッシュの稚魚において神経活動をモニターする画期的な手法が,数年前に発表された“全脳機能イメージング法”である5,6).この技術はゼブラフィッシュの稚魚が透明であること,および,脳の大きさが小さいことを利用して,脳全体の活動をCa2+イメージング法により記録するものである.この手法を応用して,視覚刺激を提示すると同時に視蓋前野の機能イメージングを試みた.こうすることにより,特定の視覚刺激に対しどのニューロンが反応し,それらのニューロンが脳のどこに存在するかを網羅的に調べることができる.神経活動の指標として用いられるCa2+インジケーターGCaMP5を神経系の全体で発現するトランスジェニックゼブラフィッシュを用いて,視蓋前野の何千ものニューロンの活動を同時に可視化することに成功した.ゼブラフィッシュの稚魚に時計回りあるいは反時計回りに回転する光学的流動を提示すると同時に,視蓋前野の個々のニューロンの反応を測定したところ,視蓋前野の多くのニューロンは,高い方向選択性をもち時計回りあるいは反時計回りのどちらかの光学的流動だけに反応すること,特定の回転速度の光学的流動に対し高い反応性を示すこと,視蓋前野において4対の空間的なクラスターを形成すること,がわかった.
回転性の光学的流動と並進性の光学的流動とを区別するようなニューロンの反応パターンが存在するかどうかを検証するため,水平に運動する光学的流動のあらゆるパターンを含むような視覚刺激プロトコールをデザインした(図2a).最初に,片方の眼のみに光学的流動(前方あるいは後方)を提示し,つぎに,両眼に対して回転性の光学的流動(時計回りあるいは反時計回り),さらに,光学的流動(前方あるいは後方)を提示した.このような視覚刺激をあたえると同時に,さきと同様の方法を用いて視蓋前野のニューロンを大規模に観察すると,多くのニューロンはある特定の視覚刺激の組合せに対し選択的に反応した.たとえば,あるニューロンは,左眼のみで前方に動く光学的流動,両眼で時計回りに動く回転性の光学的流動,両眼で前方に動く光学的流動,の3つに対し強く反応し,そのほかの光学的流動に対してはほとんど反応しなかった(図2b).このようにニューロンの反応がほぼ全か無かの反応パターンを示すことに着想を得て,それぞれのニューロンが示す反応パターンを“バーコード”のように表わすことを考えた.視覚刺激プロトコールは8つの独立した要素から構成されることから,ニューロンは理論的には2の8乗= 256通りの反応パターンをとることが可能である.そこで,Ca2+イメージング法により光学的流動に対し反応するとして同定した約3000のニューロンについて,仮想的な256の反応パターンとの相関を調べることにより7),同定したすべてのニューロンを256の異なるタイプに分類した.
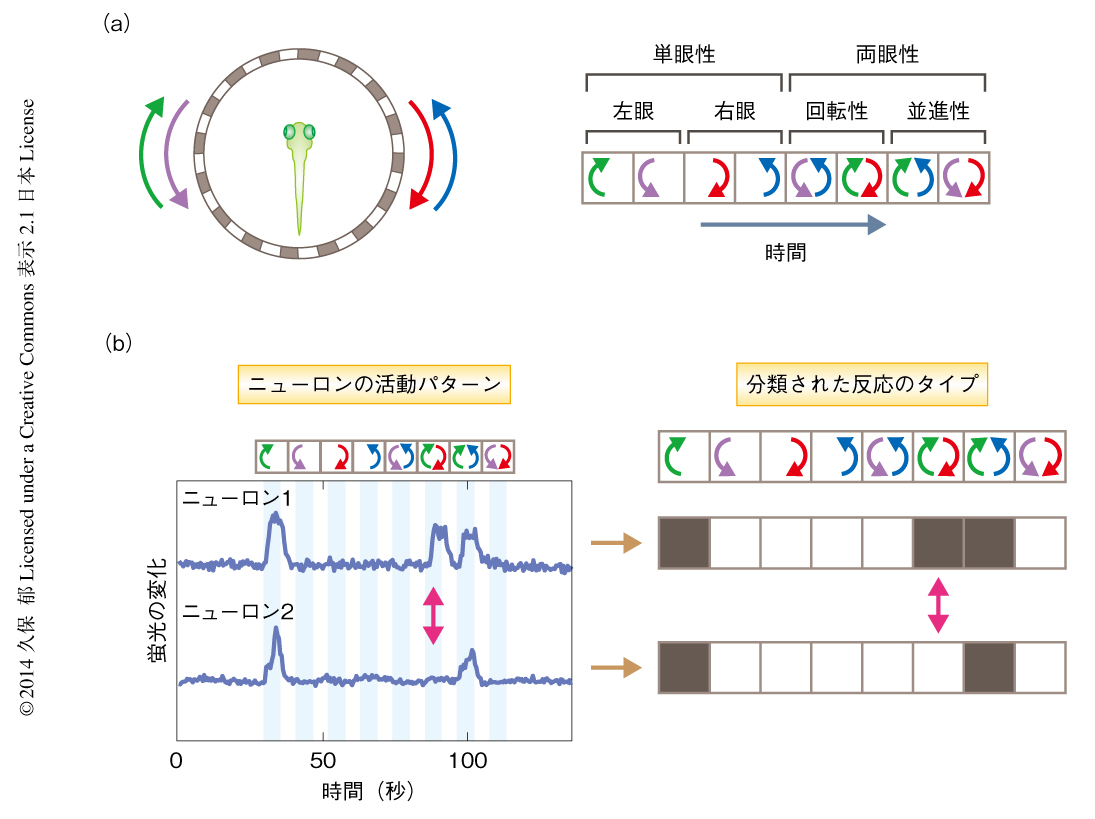
この分類を個体全体,そして,複数の個体からのデータに適用したところ,視蓋前野のニューロンには11の主要なタイプが存在することがわかった.そのうちの4つのタイプは,片方の眼のみで前方あるいは後方に動く光学的流動に対しつねに反応するニューロンであり(図2b),これまでほかの魚類において報告されていた単眼性の方向選択性をもつニューロンに対応するものと考えられた1).それに対し,そのほかの7つのタイプはより複雑な反応パターンを示した.つまり,単なる単眼性の方向選択性だけでは説明されない,より高次な演算を必要とする反応パターンを示した.たとえば,あるニューロンは,左眼のみで前方に動く光学的流動,および,両眼で前方に動く光学的流動には反応するが,両眼で時計回りに動く回転性の光学的流動には反応しなかった(図2b).これは,本来,このニューロンは左眼のみで前方に動く光学的流動に対し反応する性質をもつが,もう片方の眼(右眼)が後方に動く光学的流動をうけとった場合,この性質が“打ち消される”ことを示唆した.このようなニューロンは並進性の光学的流動に対しては強く反応するのに対し,回転性の光学的流動に対してはほとんど反応しないことから,この2つの視覚情報の特性の違いを区別することができ,また,それぞれの光学的流動のもとで異なる運動反応をひき起こすための構成要素としてはたらきうる可能性を示した.また,これらのニューロンは両方の眼からの情報を統合していることから両眼性であり,この点においてこれまで知られていたニューロンのタイプとは大きく異なっていた.興味深いことに,このような両眼性のニューロンは,回転性の光学的流動に選択的に反応するものはごく少なかったのに対し,並進性の光学的流動に選択的に反応するものは多く存在していた.以上の結果から,視蓋前野には両眼からの光学的流動に関する情報を統合するニューロンが存在しており,それらは単眼性の方向選択性をもつニューロンによる“足し算および引き算”,つまり,興奮性および抑制性の階層的な相互作用により実現されている可能性が示唆された.
この研究では,体系的な視覚刺激プロトコール,大規模なサンプル数,統計的な解析による分類,が大きな鍵となり,観察するニューロンにバイアスをかけることなく,くまなく調べることが可能になり,いままでに報告されていなかったニューロンのタイプを同定することができた.さらに,今回,新たに同定されたニューロンのタイプがどのように相互に連結し両眼性の光学的流動に関する情報を処理しているかについての仮説を導きだすことができた.興味深いことに,同時に報告された論文では,筆者らと同様の視覚刺激プロトコールおよび解析法を用いてCa2+イメージングの対象を脳全体に展開している8).結果として,互いのデータを補完し合うような結論が導きだされた.今後は,個々のニューロンの解剖学的な神経接続の形式や神経伝達物質の性質などを同定していくことにより神経接続に関する仮説を実証し神経ネットワークのより深い理解をめざしたい.
略歴:2007年 京都大学大学院生命科学研究科 修了,同年 理化学研究所 研究員,2010年 米国California大学San Francisco校 研究員を経て,2012年よりドイツMax Planck Institute of Neurobiology研究員.
研究テーマ:視覚情報にもとづき適切な運動パターンの生み出される神経機構.
関心事:光学的なアプローチを駆使したゼブラフィッシュにおける神経科学的な研究に強い魅力と可能性を感じています.
© 2014 久保 郁 Licensed under CC 表示 2.1 日本
(ドイツMax Planck Institute of Neurobiology,Department Genes-Circuits-Behavior)
email:久保 郁
DOI: 10.7875/first.author.2014.051
Functional architecture of an optic flow-responsive area that drives horizontal eye movements in zebrafish.
Fumi Kubo, Bastian Hablitzel, Marco Dal Maschio, Wolfgang Driever, Herwig Baier, Aristides B. Arrenberg
Neuron, 81, 1344-1359 (2014)
要 約
多くの動物は外界の視野全体が大きく動くような視覚刺激を経験すると,それを打ち消すようにして補完的に眼や身体を動かす反応を示す.このような視覚刺激は,視野が回転する特性をもつ場合と,視野が平行移動する特性をもつ場合の2つに大別される.これら2つの視覚特性はそれぞれ異なる運動反応をひき起こすことから,脳において互いに明確に区別される必要がある.この論文では,光遺伝学的な手法とCa2+イメージング法とを組み合わせることにより,このような視覚情報の処理にかかわる神経回路を体系的に調べた.そして,視蓋前野に存在する何千ものニューロンの活動を記録することにより,視蓋前野のニューロンは高い方向選択性をもち,単眼性あるいは両眼性の入力をうけていて,回転する視覚刺激と平行移動する視覚刺激とを区別しうるような構成要素を含んでいることが明らかにされた.筆者らのデータから,視覚刺激によりひき起こされる補完的な運動反応を駆動するのに適した神経回路の仮説が提唱された.
はじめに
われわれが道を歩いているとき,あるいは,電車に乗っているとき,外界の風景は進行方向とは逆の方向へと動いているように知覚される.このような視覚情報は“光学的流動”(optic flow)とよばれ,自分の身体が外界に対し相対的に移動することにより像が網膜のうえを流れるようにして移動することを意味する.われわれは光学的流動を感知すると,光学的流動と同じ方向に眼あるいは身体を動かすことにより,生じた視覚のエラーを最小限にとどめることができる.この反応は移動における視覚情報の処理を助けるのにきわめて効果的であり,多くの脊椎動物において広く保存されている1).
とりわけ魚類では,光学的流動が回転する特性をもつ場合には眼球を回転させる反応が誘導され,一方,光学的流動が平行移動(並進運動)する特性をもつ場合には前方あるいは後方に動くことにより身体全体を移動させる反応が誘導されることが知られている2).これらの適切な運動パターンを生み出すためには,回転性の視覚特性と並進性の視覚特性とが脳において明確に区別される必要がある(図1).個体が回転性あるいは並進性の視覚情報を区別しうるための戦略のひとつとして,両方の眼からの入力をなんらかの方法で比較し統合するという機構が考えられるが,このような情報処理あるいは演算処理が神経回路のどこでどのように行われているかについてのコンセンサスは得られていなかった.この研究では,光学的な手法による神経活動の観察および操作のしやすいゼブラフィッシュをモデルとして3,4),網膜の出力ニューロンからの直接の投射をうける視蓋前野(pretectum)に注目した.光遺伝学的な手法を用いてゼブラフィッシュにおける視蓋前野を機能的に同定したうえで,この領域における神経活動を2光子Ca2+イメージング法により単一のニューロンのレベルで記録し,視蓋前野における両眼からの視覚情報の処理の機構について解析した.
1.光遺伝学的な手法による眼球を回転させる反応の制御
ゼブラフィッシュにおいて視蓋前野が眼球を回転させる反応に関与するかどうかを調べるため,光遺伝学的な手法を用いて脳の限局した領域を光刺激した.光に依存して神経活動を活性化する効果をもつチャネルロドプシン2を神経系の全体で発現するトランスジェニックゼブラフィッシュの稚魚を用いて,光ファイバーにより片側の視蓋前野を光刺激すると両方の眼球が回転する反応が観察された.この眼球の反応は視覚刺激により誘導される眼球を回転させる反応とよく似た特性をもっていた.逆に,光に依存して神経活動を抑制する効果をもつハロロドプシンを発現させたトランスジェニックゼブラフィッシュの稚魚において視蓋前野を光刺激すると,視覚刺激によりひき起こされる眼球を回転させる反応がいちじるしく阻害された.以上から,視蓋前野は眼球を回転させる反応をひき起こすのに必要かつ十分であることが示された.
2.視覚刺激に反応する視蓋前野ニューロンのCa2+イメージング
視蓋前野のニューロンは視覚情報に対してどのように反応するのだろうか? ゼブラフィッシュの稚魚において神経活動をモニターする画期的な手法が,数年前に発表された“全脳機能イメージング法”である5,6).この技術はゼブラフィッシュの稚魚が透明であること,および,脳の大きさが小さいことを利用して,脳全体の活動をCa2+イメージング法により記録するものである.この手法を応用して,視覚刺激を提示すると同時に視蓋前野の機能イメージングを試みた.こうすることにより,特定の視覚刺激に対しどのニューロンが反応し,それらのニューロンが脳のどこに存在するかを網羅的に調べることができる.神経活動の指標として用いられるCa2+インジケーターGCaMP5を神経系の全体で発現するトランスジェニックゼブラフィッシュを用いて,視蓋前野の何千ものニューロンの活動を同時に可視化することに成功した.ゼブラフィッシュの稚魚に時計回りあるいは反時計回りに回転する光学的流動を提示すると同時に,視蓋前野の個々のニューロンの反応を測定したところ,視蓋前野の多くのニューロンは,高い方向選択性をもち時計回りあるいは反時計回りのどちらかの光学的流動だけに反応すること,特定の回転速度の光学的流動に対し高い反応性を示すこと,視蓋前野において4対の空間的なクラスターを形成すること,がわかった.
3.回転性あるいは並進性の光学的流動に選択的に反応するニューロンの同定
回転性の光学的流動と並進性の光学的流動とを区別するようなニューロンの反応パターンが存在するかどうかを検証するため,水平に運動する光学的流動のあらゆるパターンを含むような視覚刺激プロトコールをデザインした(図2a).最初に,片方の眼のみに光学的流動(前方あるいは後方)を提示し,つぎに,両眼に対して回転性の光学的流動(時計回りあるいは反時計回り),さらに,光学的流動(前方あるいは後方)を提示した.このような視覚刺激をあたえると同時に,さきと同様の方法を用いて視蓋前野のニューロンを大規模に観察すると,多くのニューロンはある特定の視覚刺激の組合せに対し選択的に反応した.たとえば,あるニューロンは,左眼のみで前方に動く光学的流動,両眼で時計回りに動く回転性の光学的流動,両眼で前方に動く光学的流動,の3つに対し強く反応し,そのほかの光学的流動に対してはほとんど反応しなかった(図2b).このようにニューロンの反応がほぼ全か無かの反応パターンを示すことに着想を得て,それぞれのニューロンが示す反応パターンを“バーコード”のように表わすことを考えた.視覚刺激プロトコールは8つの独立した要素から構成されることから,ニューロンは理論的には2の8乗= 256通りの反応パターンをとることが可能である.そこで,Ca2+イメージング法により光学的流動に対し反応するとして同定した約3000のニューロンについて,仮想的な256の反応パターンとの相関を調べることにより7),同定したすべてのニューロンを256の異なるタイプに分類した.
4.体系的および定量的な解析を用いたニューロンの分類
この分類を個体全体,そして,複数の個体からのデータに適用したところ,視蓋前野のニューロンには11の主要なタイプが存在することがわかった.そのうちの4つのタイプは,片方の眼のみで前方あるいは後方に動く光学的流動に対しつねに反応するニューロンであり(図2b),これまでほかの魚類において報告されていた単眼性の方向選択性をもつニューロンに対応するものと考えられた1).それに対し,そのほかの7つのタイプはより複雑な反応パターンを示した.つまり,単なる単眼性の方向選択性だけでは説明されない,より高次な演算を必要とする反応パターンを示した.たとえば,あるニューロンは,左眼のみで前方に動く光学的流動,および,両眼で前方に動く光学的流動には反応するが,両眼で時計回りに動く回転性の光学的流動には反応しなかった(図2b).これは,本来,このニューロンは左眼のみで前方に動く光学的流動に対し反応する性質をもつが,もう片方の眼(右眼)が後方に動く光学的流動をうけとった場合,この性質が“打ち消される”ことを示唆した.このようなニューロンは並進性の光学的流動に対しては強く反応するのに対し,回転性の光学的流動に対してはほとんど反応しないことから,この2つの視覚情報の特性の違いを区別することができ,また,それぞれの光学的流動のもとで異なる運動反応をひき起こすための構成要素としてはたらきうる可能性を示した.また,これらのニューロンは両方の眼からの情報を統合していることから両眼性であり,この点においてこれまで知られていたニューロンのタイプとは大きく異なっていた.興味深いことに,このような両眼性のニューロンは,回転性の光学的流動に選択的に反応するものはごく少なかったのに対し,並進性の光学的流動に選択的に反応するものは多く存在していた.以上の結果から,視蓋前野には両眼からの光学的流動に関する情報を統合するニューロンが存在しており,それらは単眼性の方向選択性をもつニューロンによる“足し算および引き算”,つまり,興奮性および抑制性の階層的な相互作用により実現されている可能性が示唆された.
おわりに
この研究では,体系的な視覚刺激プロトコール,大規模なサンプル数,統計的な解析による分類,が大きな鍵となり,観察するニューロンにバイアスをかけることなく,くまなく調べることが可能になり,いままでに報告されていなかったニューロンのタイプを同定することができた.さらに,今回,新たに同定されたニューロンのタイプがどのように相互に連結し両眼性の光学的流動に関する情報を処理しているかについての仮説を導きだすことができた.興味深いことに,同時に報告された論文では,筆者らと同様の視覚刺激プロトコールおよび解析法を用いてCa2+イメージングの対象を脳全体に展開している8).結果として,互いのデータを補完し合うような結論が導きだされた.今後は,個々のニューロンの解剖学的な神経接続の形式や神経伝達物質の性質などを同定していくことにより神経接続に関する仮説を実証し神経ネットワークのより深い理解をめざしたい.
文 献
- Masseck, O. A. & Hoffmann, K. -P.: Comparative neurobiology of the optokinetic reflex. Ann. NY Acad. Sci., 1164, 430-439 (2009)[PubMed]
- Neuhauss, S. C. F.: Behavioral genetic approaches to visual system development and function in zebrafish. J. Neurobiol., 54, 148-160 (2003)[PubMed]
- Portugues, R., Severi, K. E., Wyart, C. et al.: Optogenetics in a transparent animal: circuit function in the larval zebrafish. Curr. Opin. Neurobiol., 23, 119-126 (2013)[PubMed]
- McLean, D. L. & Fetcho, J. R.: Movement, technology and discovery in the zebrafish. Curr. Opin. Neurobiol., 21, 110-115 (2011)[PubMed]
- Ahrens, M. B., Li, J. M., Orger, M. B. et al.: Brain-wide neuronal dynamics during motor adaptation in zebrafish. Nature, 485, 471-477 (2012)[PubMed]
- Ahrens, M. B., Orger, M. B., Robson, D. N. et al.: Whole-brain functional imaging at cellular resolution using light-sheet microscopy. Nat. Methods, 10, 413-420 (2013)[PubMed]
- Miri, A., Daie, K., Burdine, R. D. et al.: Regression-based identification of behavior-encoding neurons during large-scale optical imaging of neural activity at cellular resolution. J. Neurophysiol. 105, 964-980 (2011)[PubMed]
- Portugues, R., Feierstein, C. E., Engert, F. et al.: Whole-brain activity maps reveal stereotyped, distributed networks for visuomotor behavior. Neuron, 81, 1328-1343 (2014)[PubMed]
著者プロフィール
略歴:2007年 京都大学大学院生命科学研究科 修了,同年 理化学研究所 研究員,2010年 米国California大学San Francisco校 研究員を経て,2012年よりドイツMax Planck Institute of Neurobiology研究員.
研究テーマ:視覚情報にもとづき適切な運動パターンの生み出される神経機構.
関心事:光学的なアプローチを駆使したゼブラフィッシュにおける神経科学的な研究に強い魅力と可能性を感じています.
© 2014 久保 郁 Licensed under CC 表示 2.1 日本
