平面内細胞極性にかかわるタンパク質が共通の細胞骨格を制御して細胞集団の極性制御と繊毛形成とを制御する
進藤 麻子
(米国Texas大学Austin校Section of Molecular Cell and Developmental Biology)
email:進藤麻子
DOI: 10.7875/first.author.2010.014
Planar cell polarity acts through septins to control collective cell movement and ciliogenesis.
Su Kyoung Kim, Asako Shindo, Tae Joo Park, Edwin C. Oh, Srimoyee Ghosh, Ryan S. Gray, Richard A. Lewis, Colin A. Johnson, Tania Attie-Bittach, Nicholas Katsanis, John B. Wallingford
Science, 329, 1337-1340 (2010)
発生過程において細胞極性を制御する平面内細胞極性(PCP)経路の新たなターゲットがみつかった.セプチンは細胞分裂や細胞移動に必要な細胞骨格タンパク質として知られているが,筆者らは,アフリカツメガエル胚をモデルとしてPCP経路が関与する細胞運動と繊毛形成とに着目し,セプチンがPCP経路の関連タンパク質であるFritzによってその局在を制御されていることを発見した.Fritzとセプチンの機能は正常な細胞極性の形成にもとづいた細胞運動と繊毛形成の双方に必要であること,さらには,繊毛形成不全が原因とされるヒト遺伝病ではFritzに変異の生じていることも明らかにした.この研究は,PCP経路の新たなターゲットを示すとともに,PCP経路が制御する細胞運動と繊毛病との関連性を検討する必要性を提唱するものである.
生物の発生過程において,細胞がある規則にしたがって変形し整列することは,適切な組織を形成するために必須の現象である.細胞がどのように変形・整列し,最終的にどのような形態をとるかは組織によって異なるが,“揃える”という現象(細胞極性の形成)を制御する共通のシグナル経路は,組織や種をこえて存在している.それが,平面内細胞極性(planar cell polarity:PCP)経路である1).PCP経路ではコアPCPとよばれるいくつかのタンパク質が主要な役割を担っていて,これらが欠けると正常な細胞極性は確立されず,異常な形態の胚になることが知られている.しかし一方で,これらタンパク質の重要性は明らかであるものの,どのような分子機構によりどんなタンパク質を介して細胞にはたらきかけているのかについては多くが不明である.唯一,細胞内で細胞骨格を構成するアクチンの挙動がPCP経路により決められていることが知られているが,それ以外はほとんどわかっていない.この点を明らかにするためには,細胞骨格といった細胞内動態の変化を直接に制御しているタンパク質により近く,コアPCPよりも下流で機能しているタンパク質に着目する必要がある.そこで筆者らは,アフリカツメガエル胚の原腸形成の際にみられる,PCP経路によって制御されている“収斂と伸長”(convergent extension)とよばれる細胞運動をモデルとし2),コアPCPより下流で機能すると報告されているタンパク質Fritzの機能を解析することで3),この問題に取り組んできた.収斂と伸長は,のちに脊索となる領域(背側の中胚葉)で起こり,アフリカツメガエル胚の原腸陥入に必須である.この細胞運動ではそれぞれの細胞が頭尾軸に垂直の方向に伸長し(細胞極性の確立),さらに,同じ方向に互いに入り込むようにして配列することで脊索の頭尾方向への伸長を可能にしている(図1).この過程でコアPCPの機能が阻害されると細胞の伸長が抑制されて配列も乱れ,短い脊索が形成されたり原腸陥入運動が阻害されたりする.
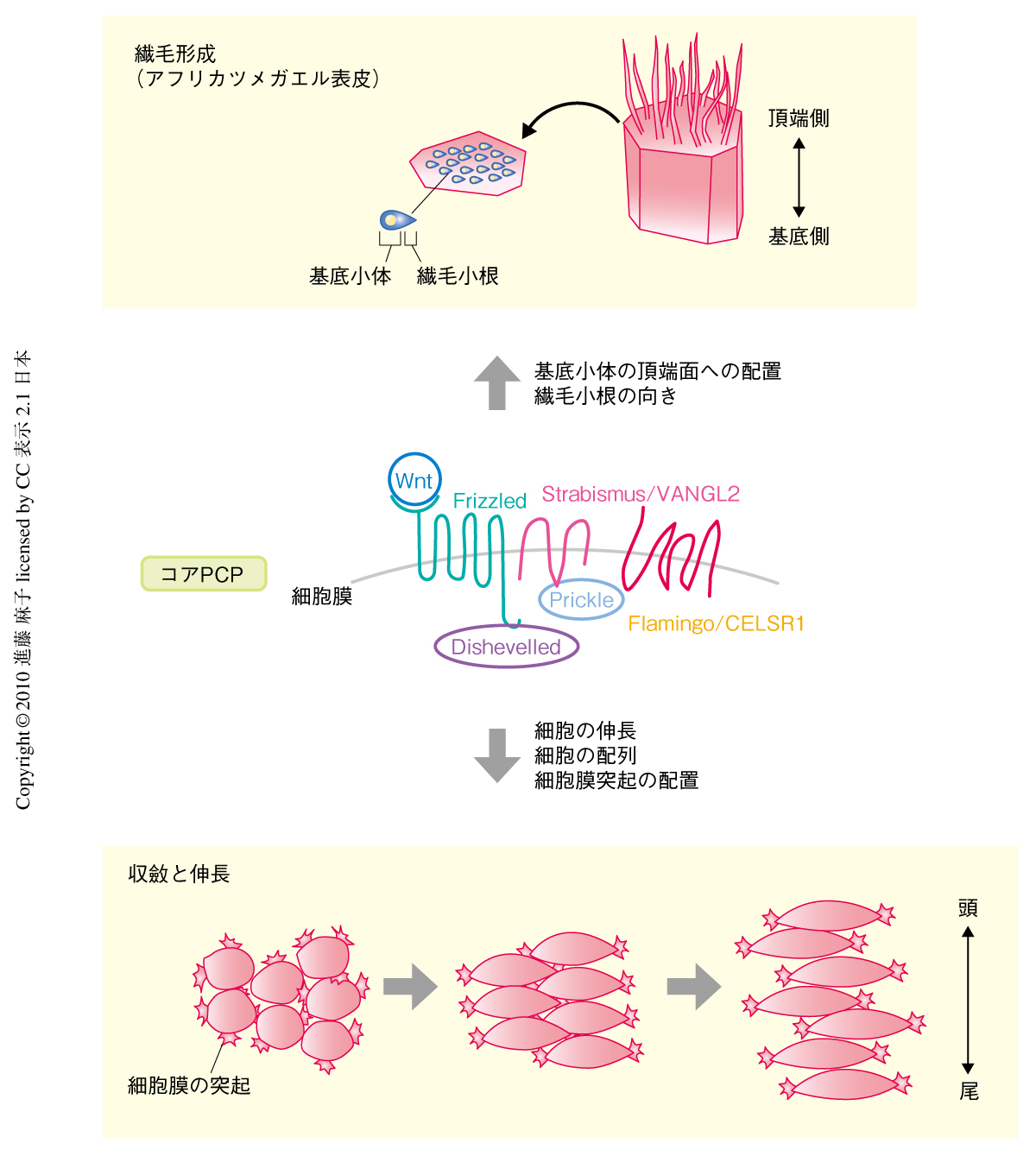
これにくわえて,PCP経路関連タンパク質がかかわっていると報告されている発生現象に,繊毛形成があげられる4)(図1).ほとんどの細胞にアンテナのように存在する繊毛(cilia)は,生物の左右軸の形成や5),初期胚の形態形成に重要なShhシグナル伝達に必須であることが示されており6),形態形成において欠かせない研究対象となっている.これらシグナルに関与する繊毛は一次繊毛(primary cilia)とよばれ,このほかに可動性をもつ運動性繊毛(motile cilia)とよばれる繊毛も存在する.一次繊毛と運動性繊毛の形成は共通のタンパク質により制御されていることが知られている.筆者らは,これまでにアフリカツメガエル胚の表皮の繊毛細胞をモデルとして,コアPCPのひとつが繊毛の根元にある基底小体を細胞の頂端側に配置させ,かつ,その向きの制御に必要であることを示してきた4)(図1).また,発生過程における繊毛形成の不全はヒトでは遺伝病として現れ,Bardet-Biedl症候群やMeckel-Gruber症候群などが知られている.
PCP経路の制御機構を明らかにすることにくわえ,これまで検討されてこなかった,最終形態の異なるさまざまな組織の形態形成がどのように制御されているのかという疑問を解決するため,今回の研究では,収斂と伸長および繊毛形成におけるFritzの役割を解析した.
収斂と伸長におけるFritzの機能をモルフォリノアンチセンスオリゴの胚への注入により阻害したところ,胚の形態においてはコアPCPを阻害したような原腸陥入の異常がみられたが,細胞レベルの異常についてはコアPCPの阻害とはやや異なる形質が得られた.コアPCPの阻害では細胞の伸長は起こらず,同時に,その配列も乱れているが,Fritzの阻害では細胞の伸長は弱まるものの,配列は維持されていた.細胞の変形がさまたげられている原因として,細胞膜の安定性の損なわれていることが考えられた.収斂と伸長を起こしている細胞は運動性が高く,となりどうしの細胞の接着は弱まっているが,細胞膜は適切な強度が保たれてピンと張った状態を維持している.Fritzを阻害すると,この細胞膜の強度が損なわれるためか,たるんだ状態のようにみえ,細胞運動の動画を撮影するとその違いは明らかであった.重要な点は,コアPCPを阻害した場合にも同様の細胞膜の異常がみられたことである.これらのことは,FritzがコアPCPの制御する現象の一部を担っており,その下流で機能していることを支持するものであった.さらに,Fritzは細胞膜および細胞膜に近い細胞質に局在していることがわかった.しかしながら,Fritzがどのように細胞膜の安定性を制御しているかについては,これだけでは明らかではない.
そこでつぎに,Fritzのターゲットとしてセプチン(Septin2とSeptin7)に着目した7).セプチンは,in vitroの研究で,細胞膜にそって局在し膜の剛性を維持する機能をもっていることが報告されている8).そこで,生化学的な手法によりFritzがセプチンと直接もしくは間接的に結合しうることを明らかにし,そこから,上述した収斂と伸長を起こしている細胞の膜動態をセプチンが最終的に制御しているという可能性を考えた.そこで,セプチンの機能をモルフォリノアンチセンスオリゴで阻害してその影響を観察した.すると,Fritzを阻害したときと同様に,細胞膜の安定性が失われて細胞の伸長が阻害され,正常な原腸陥入運動が起こらなかった.さらに,Septin2の局在を観察したところ,Fritzの存在下では細胞膜および膜近辺に限局していたが,Fritzの非存在下では細胞膜からより離れた細胞質まで広がって分布していた.これらのことから,収斂と伸長のときのPCP経路による制御のひとつとして,Fritzがセプチンを正しく局在させ,細胞膜の安定化を介して細胞伸長を促す,という経路の存在が明らかとなった(図2,左).
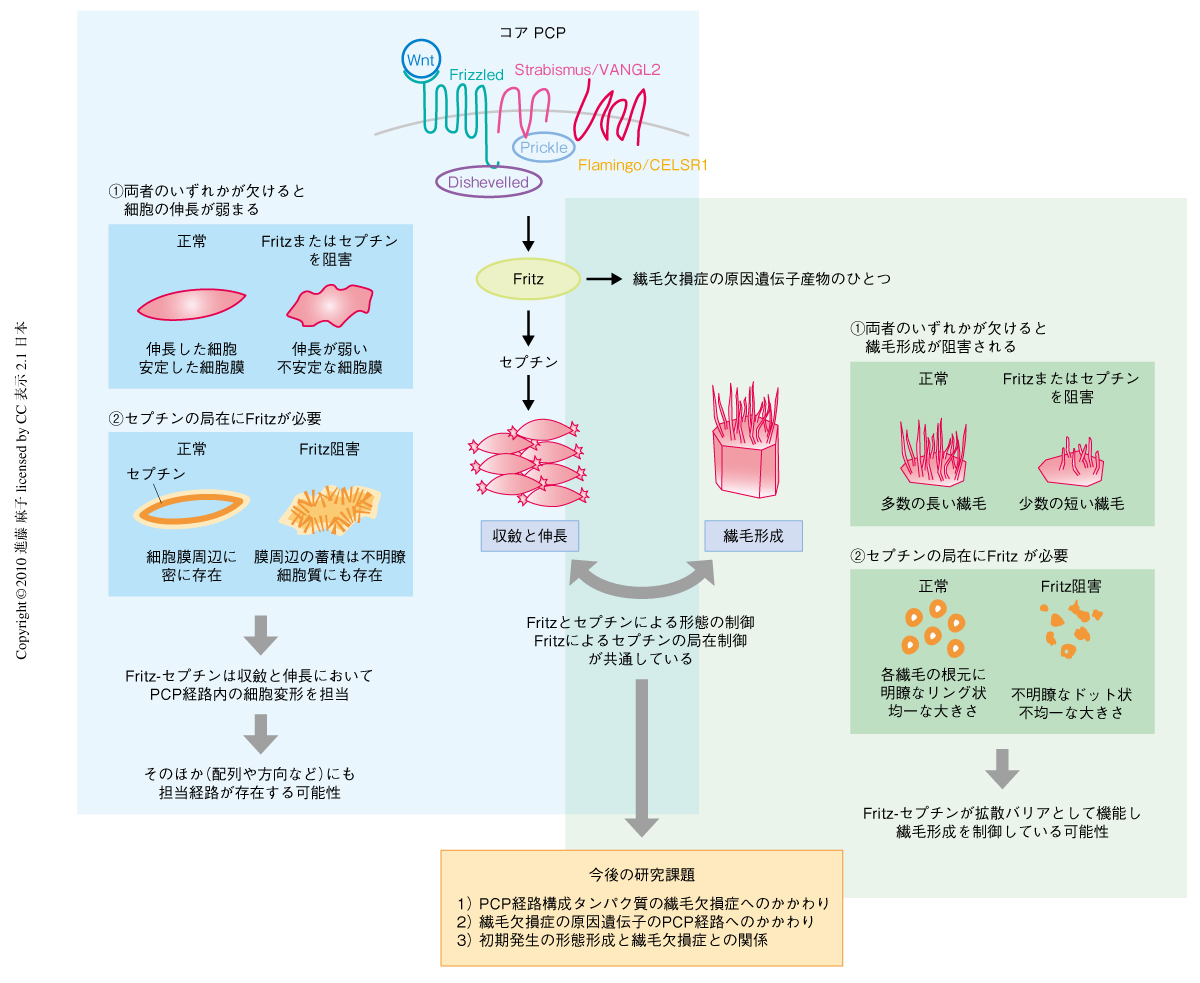
収斂と伸長においてPCP経路にFritzとセプチンとが関与していたことをふまえ,アフリカツメガエルの体表に存在する繊毛細胞を用いて,同じくPCP経路関連タンパク質が関与するとされる繊毛形成におけるそれらの役割を調べてみた.すると,Fritzの阻害により繊毛形成は正常に行なわれず,セプチンも同様に繊毛形成に必要であることがわかった.すなわち,Fritzまたはセプチンのモルフォリノアンチセンスオリゴを胚に注入すると,通常は何本もの長い繊毛が形成されるべきところ,短い繊毛がわずかに形成されるにとどまっていた.興味深いことに,正常な状態ではセプチンは繊毛の根元にリング状に局在しているが,Fritzの機能を阻害するとこの局在がくずれることがわかった.このリング状の構造は,つい最近,培養細胞を用いた研究により,繊毛の拡散バリア(繊毛を構成する膜タンパク質と細胞本体の膜構造とが混ざるのを防ぐ構造)としての機能をもつ可能性が報告された9).繊毛は細胞本体とは異なる特有の膜タンパク質をもち,この拡散バリアがなければ特有の構造が形成・維持できず,シグナル伝達も正常に行なわれない.おそらく,Fritzとセプチンはこの拡散バリアを形成するために必須であるものと予想される.これに関連し,Fritzとセプチンの存在が一次繊毛によるシグナル伝達に重要であるかどうかを検討するため,Fritzとセプチンのいずれかが機能しないとき,Shhシグナル伝達が阻害されるかどうかを検討した.原腸形成の終了したのち,神経管領域ではShhに誘導されるタンパク質が発現するが,Fritzまたはセプチンを阻害したところ,これらShhの下流タンパク質の発現誘導は抑えられていることがわかった.このことから,Fritzとセプチンによる繊毛形成の制御は,運動性繊毛においても一次繊毛においても,その機能維持のために必須であることが明らかとなった(図2,右).
繊毛形成の不全によるヒトの遺伝病がいくつか知られている.Bardet-Biedl症候群およびMeckel-Gruber症候群は10),繊毛の形成不全が原因とされる典型的な症状を示す.おもな症状として,多指症,知能発育不全,腎不全,網膜異常,肥満,などがあげられる.これらの症状がどのようにひき起こされるのかについては不明な点が多いが,繊毛を必須とするShhシグナルの欠損が原因のひとつと考えられている.しかしながら,それだけでは説明できない症状も多い.今回の研究で,Bardet-Biedl症候群あるいはMeckel-Gruber症候群と診断された患者の疫学的な調査から,5人の患者にFritzの変異が起こっていることを発見した.この5人のFritz変異部位に共通性はなかったが,興味深いことに,Fritzは立体構造予測によるとβプロペラ構造をとるが,その立体構造の同一面に変異の起こっていることが2人の患者において共通していた.その面に変異が存在することにより,セプチンなどなんらかのタンパク質とFritzとの相互作用に影響が及び,繊毛病がひき起こされている可能性が予想できる.また,このFritzの変異がBardet-Biedl症候群を起こすに十分であることを患者のある家族の系譜より明らかにした.このことは,Fritzが種をこえてその機能の重要性を保持していることを示している.
今回の結果から,Fritzが異なる発生段階の異なる組織で,セプチンという共通したターゲットを制御していることがわかった.さらに,Fritzの変異がヒトの遺伝病の原因となっていたことから,筆者らは,初期発生を制御するPCP経路,シグナル伝達に必須な繊毛形成,ヒトの繊毛病の関連性を総合的に検討する必要性を提唱している.
アフリカツメガエル胚の脊索形成の過程で生じる収斂と伸長は,PCP経路により制御される現象としてここ20年ほど注目されてきた.しかしこれまで,その分子機構は,コアPCPの機能阻害実験によりPCP経路を遮断すると細胞の適切な形態変化が起こらず配列も乱れる,といった,“すべて壊れる”状態でしか検討できていなかった.今回の結果は,Fritzという,これまで脊椎動物で調べられていなかったコアPCPの下流のタンパク質に着目したことで,細胞のかたちを特異的に制御している新たなルートが明らかにされた点で重要である.このことは,コアPCPの下流にそのほかの現象,たとえば,細胞の配列やオルガネラの配置をそれぞれ特異的に制御するルートの存在する可能性を示唆している.これらを明らかにするためには,当然ながらFritzと同様にコアPCPの下流で機能するとされるFuzzyやInturnedも興味深いが11),筆者らは,ヒトの繊毛病の原因遺伝子として報告されているBBS遺伝子群の機能に着目することも重要であると考えている.Fritz以外にも,ヒトの繊毛病の原因遺伝子産物とされているものがPCP経路にかかわっている可能性は高い.さらには,収斂と伸長のようなPCP経路が制御している細胞運動の異常が,ヒト繊毛病の症状の原因として含まれているかどうかを検討する必要もある.
今回の研究は,発生過程におけるPCP経路の制御機構の解明と繊毛病の原因の解明,この双方の研究に今後の道筋を示し,ひいては,ヒト遺伝病の原因解明のためには発生の根幹をなす細胞極性や運動の解析もまた重要であることをあらためて示したものといえる.
略歴:2008年 総合研究大学院大学生命科学研究科博士課程 修了,同年 基礎生物学研究所 研究員(上野直人研究室)を経て,2009年より米国Texas大学Austin校 研究員(John Wallingford研究室).
研究テーマ:アフリカツメガエル初期胚における細胞極性の形成機構の解明.
抱負:医学はもっと発生学的な思考と知識を取り込む必要があるのではないかという(漠然とした)意識をもち,獣医学部を卒業したのち,発生学を専門として研究をつづけている.カエルのたまごひとつをとっても解明困難な発生現象がたたみかけるように現れ,目がくらみつつも,ひとつでも何かを発見し解明していくことが人類の未来を開くものと信じて,日々,カエルのたまご相手に奮闘している.
© 2010 進藤 麻子 Licensed under CC 表示 2.1 日本
(米国Texas大学Austin校Section of Molecular Cell and Developmental Biology)
email:進藤麻子
DOI: 10.7875/first.author.2010.014
Planar cell polarity acts through septins to control collective cell movement and ciliogenesis.
Su Kyoung Kim, Asako Shindo, Tae Joo Park, Edwin C. Oh, Srimoyee Ghosh, Ryan S. Gray, Richard A. Lewis, Colin A. Johnson, Tania Attie-Bittach, Nicholas Katsanis, John B. Wallingford
Science, 329, 1337-1340 (2010)
要 約
発生過程において細胞極性を制御する平面内細胞極性(PCP)経路の新たなターゲットがみつかった.セプチンは細胞分裂や細胞移動に必要な細胞骨格タンパク質として知られているが,筆者らは,アフリカツメガエル胚をモデルとしてPCP経路が関与する細胞運動と繊毛形成とに着目し,セプチンがPCP経路の関連タンパク質であるFritzによってその局在を制御されていることを発見した.Fritzとセプチンの機能は正常な細胞極性の形成にもとづいた細胞運動と繊毛形成の双方に必要であること,さらには,繊毛形成不全が原因とされるヒト遺伝病ではFritzに変異の生じていることも明らかにした.この研究は,PCP経路の新たなターゲットを示すとともに,PCP経路が制御する細胞運動と繊毛病との関連性を検討する必要性を提唱するものである.
はじめに
生物の発生過程において,細胞がある規則にしたがって変形し整列することは,適切な組織を形成するために必須の現象である.細胞がどのように変形・整列し,最終的にどのような形態をとるかは組織によって異なるが,“揃える”という現象(細胞極性の形成)を制御する共通のシグナル経路は,組織や種をこえて存在している.それが,平面内細胞極性(planar cell polarity:PCP)経路である1).PCP経路ではコアPCPとよばれるいくつかのタンパク質が主要な役割を担っていて,これらが欠けると正常な細胞極性は確立されず,異常な形態の胚になることが知られている.しかし一方で,これらタンパク質の重要性は明らかであるものの,どのような分子機構によりどんなタンパク質を介して細胞にはたらきかけているのかについては多くが不明である.唯一,細胞内で細胞骨格を構成するアクチンの挙動がPCP経路により決められていることが知られているが,それ以外はほとんどわかっていない.この点を明らかにするためには,細胞骨格といった細胞内動態の変化を直接に制御しているタンパク質により近く,コアPCPよりも下流で機能しているタンパク質に着目する必要がある.そこで筆者らは,アフリカツメガエル胚の原腸形成の際にみられる,PCP経路によって制御されている“収斂と伸長”(convergent extension)とよばれる細胞運動をモデルとし2),コアPCPより下流で機能すると報告されているタンパク質Fritzの機能を解析することで3),この問題に取り組んできた.収斂と伸長は,のちに脊索となる領域(背側の中胚葉)で起こり,アフリカツメガエル胚の原腸陥入に必須である.この細胞運動ではそれぞれの細胞が頭尾軸に垂直の方向に伸長し(細胞極性の確立),さらに,同じ方向に互いに入り込むようにして配列することで脊索の頭尾方向への伸長を可能にしている(図1).この過程でコアPCPの機能が阻害されると細胞の伸長が抑制されて配列も乱れ,短い脊索が形成されたり原腸陥入運動が阻害されたりする.
これにくわえて,PCP経路関連タンパク質がかかわっていると報告されている発生現象に,繊毛形成があげられる4)(図1).ほとんどの細胞にアンテナのように存在する繊毛(cilia)は,生物の左右軸の形成や5),初期胚の形態形成に重要なShhシグナル伝達に必須であることが示されており6),形態形成において欠かせない研究対象となっている.これらシグナルに関与する繊毛は一次繊毛(primary cilia)とよばれ,このほかに可動性をもつ運動性繊毛(motile cilia)とよばれる繊毛も存在する.一次繊毛と運動性繊毛の形成は共通のタンパク質により制御されていることが知られている.筆者らは,これまでにアフリカツメガエル胚の表皮の繊毛細胞をモデルとして,コアPCPのひとつが繊毛の根元にある基底小体を細胞の頂端側に配置させ,かつ,その向きの制御に必要であることを示してきた4)(図1).また,発生過程における繊毛形成の不全はヒトでは遺伝病として現れ,Bardet-Biedl症候群やMeckel-Gruber症候群などが知られている.
PCP経路の制御機構を明らかにすることにくわえ,これまで検討されてこなかった,最終形態の異なるさまざまな組織の形態形成がどのように制御されているのかという疑問を解決するため,今回の研究では,収斂と伸長および繊毛形成におけるFritzの役割を解析した.
1.PCP経路の制御下にFritzにより制御されるセプチンがくわわった
収斂と伸長におけるFritzの機能をモルフォリノアンチセンスオリゴの胚への注入により阻害したところ,胚の形態においてはコアPCPを阻害したような原腸陥入の異常がみられたが,細胞レベルの異常についてはコアPCPの阻害とはやや異なる形質が得られた.コアPCPの阻害では細胞の伸長は起こらず,同時に,その配列も乱れているが,Fritzの阻害では細胞の伸長は弱まるものの,配列は維持されていた.細胞の変形がさまたげられている原因として,細胞膜の安定性の損なわれていることが考えられた.収斂と伸長を起こしている細胞は運動性が高く,となりどうしの細胞の接着は弱まっているが,細胞膜は適切な強度が保たれてピンと張った状態を維持している.Fritzを阻害すると,この細胞膜の強度が損なわれるためか,たるんだ状態のようにみえ,細胞運動の動画を撮影するとその違いは明らかであった.重要な点は,コアPCPを阻害した場合にも同様の細胞膜の異常がみられたことである.これらのことは,FritzがコアPCPの制御する現象の一部を担っており,その下流で機能していることを支持するものであった.さらに,Fritzは細胞膜および細胞膜に近い細胞質に局在していることがわかった.しかしながら,Fritzがどのように細胞膜の安定性を制御しているかについては,これだけでは明らかではない.
そこでつぎに,Fritzのターゲットとしてセプチン(Septin2とSeptin7)に着目した7).セプチンは,in vitroの研究で,細胞膜にそって局在し膜の剛性を維持する機能をもっていることが報告されている8).そこで,生化学的な手法によりFritzがセプチンと直接もしくは間接的に結合しうることを明らかにし,そこから,上述した収斂と伸長を起こしている細胞の膜動態をセプチンが最終的に制御しているという可能性を考えた.そこで,セプチンの機能をモルフォリノアンチセンスオリゴで阻害してその影響を観察した.すると,Fritzを阻害したときと同様に,細胞膜の安定性が失われて細胞の伸長が阻害され,正常な原腸陥入運動が起こらなかった.さらに,Septin2の局在を観察したところ,Fritzの存在下では細胞膜および膜近辺に限局していたが,Fritzの非存在下では細胞膜からより離れた細胞質まで広がって分布していた.これらのことから,収斂と伸長のときのPCP経路による制御のひとつとして,Fritzがセプチンを正しく局在させ,細胞膜の安定化を介して細胞伸長を促す,という経路の存在が明らかとなった(図2,左).
2.Fritzとセプチンは繊毛形成も制御する
収斂と伸長においてPCP経路にFritzとセプチンとが関与していたことをふまえ,アフリカツメガエルの体表に存在する繊毛細胞を用いて,同じくPCP経路関連タンパク質が関与するとされる繊毛形成におけるそれらの役割を調べてみた.すると,Fritzの阻害により繊毛形成は正常に行なわれず,セプチンも同様に繊毛形成に必要であることがわかった.すなわち,Fritzまたはセプチンのモルフォリノアンチセンスオリゴを胚に注入すると,通常は何本もの長い繊毛が形成されるべきところ,短い繊毛がわずかに形成されるにとどまっていた.興味深いことに,正常な状態ではセプチンは繊毛の根元にリング状に局在しているが,Fritzの機能を阻害するとこの局在がくずれることがわかった.このリング状の構造は,つい最近,培養細胞を用いた研究により,繊毛の拡散バリア(繊毛を構成する膜タンパク質と細胞本体の膜構造とが混ざるのを防ぐ構造)としての機能をもつ可能性が報告された9).繊毛は細胞本体とは異なる特有の膜タンパク質をもち,この拡散バリアがなければ特有の構造が形成・維持できず,シグナル伝達も正常に行なわれない.おそらく,Fritzとセプチンはこの拡散バリアを形成するために必須であるものと予想される.これに関連し,Fritzとセプチンの存在が一次繊毛によるシグナル伝達に重要であるかどうかを検討するため,Fritzとセプチンのいずれかが機能しないとき,Shhシグナル伝達が阻害されるかどうかを検討した.原腸形成の終了したのち,神経管領域ではShhに誘導されるタンパク質が発現するが,Fritzまたはセプチンを阻害したところ,これらShhの下流タンパク質の発現誘導は抑えられていることがわかった.このことから,Fritzとセプチンによる繊毛形成の制御は,運動性繊毛においても一次繊毛においても,その機能維持のために必須であることが明らかとなった(図2,右).
3.Fritzの変異はヒトの遺伝病の原因となっていた
繊毛形成の不全によるヒトの遺伝病がいくつか知られている.Bardet-Biedl症候群およびMeckel-Gruber症候群は10),繊毛の形成不全が原因とされる典型的な症状を示す.おもな症状として,多指症,知能発育不全,腎不全,網膜異常,肥満,などがあげられる.これらの症状がどのようにひき起こされるのかについては不明な点が多いが,繊毛を必須とするShhシグナルの欠損が原因のひとつと考えられている.しかしながら,それだけでは説明できない症状も多い.今回の研究で,Bardet-Biedl症候群あるいはMeckel-Gruber症候群と診断された患者の疫学的な調査から,5人の患者にFritzの変異が起こっていることを発見した.この5人のFritz変異部位に共通性はなかったが,興味深いことに,Fritzは立体構造予測によるとβプロペラ構造をとるが,その立体構造の同一面に変異の起こっていることが2人の患者において共通していた.その面に変異が存在することにより,セプチンなどなんらかのタンパク質とFritzとの相互作用に影響が及び,繊毛病がひき起こされている可能性が予想できる.また,このFritzの変異がBardet-Biedl症候群を起こすに十分であることを患者のある家族の系譜より明らかにした.このことは,Fritzが種をこえてその機能の重要性を保持していることを示している.
4.初期発生の細胞集団のふるまい,繊毛形成,ヒトの繊毛病をつなぐFritz
今回の結果から,Fritzが異なる発生段階の異なる組織で,セプチンという共通したターゲットを制御していることがわかった.さらに,Fritzの変異がヒトの遺伝病の原因となっていたことから,筆者らは,初期発生を制御するPCP経路,シグナル伝達に必須な繊毛形成,ヒトの繊毛病の関連性を総合的に検討する必要性を提唱している.
おわりに
アフリカツメガエル胚の脊索形成の過程で生じる収斂と伸長は,PCP経路により制御される現象としてここ20年ほど注目されてきた.しかしこれまで,その分子機構は,コアPCPの機能阻害実験によりPCP経路を遮断すると細胞の適切な形態変化が起こらず配列も乱れる,といった,“すべて壊れる”状態でしか検討できていなかった.今回の結果は,Fritzという,これまで脊椎動物で調べられていなかったコアPCPの下流のタンパク質に着目したことで,細胞のかたちを特異的に制御している新たなルートが明らかにされた点で重要である.このことは,コアPCPの下流にそのほかの現象,たとえば,細胞の配列やオルガネラの配置をそれぞれ特異的に制御するルートの存在する可能性を示唆している.これらを明らかにするためには,当然ながらFritzと同様にコアPCPの下流で機能するとされるFuzzyやInturnedも興味深いが11),筆者らは,ヒトの繊毛病の原因遺伝子として報告されているBBS遺伝子群の機能に着目することも重要であると考えている.Fritz以外にも,ヒトの繊毛病の原因遺伝子産物とされているものがPCP経路にかかわっている可能性は高い.さらには,収斂と伸長のようなPCP経路が制御している細胞運動の異常が,ヒト繊毛病の症状の原因として含まれているかどうかを検討する必要もある.
今回の研究は,発生過程におけるPCP経路の制御機構の解明と繊毛病の原因の解明,この双方の研究に今後の道筋を示し,ひいては,ヒト遺伝病の原因解明のためには発生の根幹をなす細胞極性や運動の解析もまた重要であることをあらためて示したものといえる.
文 献
- Seifert, J. R. & Mlodzik, M.: Frizzled/PCP signalling: a conserved mechanism regulating cell polarity and directed motility. Nat. Rev. Genet., 8, 126-138 (2007)[PubMed]
- Davidson, L. A., Joshi, S. D., Kim, H. Y. et al.: Emergent morphogenesis: elastic mechanics of a self-deforming tissue. J. Biomech., 43, 63-70 (2010)[PubMed]
- Collier, S., Lee, H., Burgess, R. et al.: The WD40 repeat protein fritz links cytoskeletal planar polarity to frizzled subcellular localization in the Drosophila epidermis. Genetics, 169, 2035-2045 (2005)[PubMed]
- Park, T. J., Mitchell, B. J., Abitua, P. B. et al.: Dishevelled controls apical docking and planar polarization of basal bodies in ciliated epithelial cells. Nat. Genet., 40, 871-879 (2008)[PubMed]
- Hirokawa, N., Tanaka, Y, Okada, Y. et al.: Nodal flow and the generation of left-right asymmetry. Cell, 125, 33-45 (2006)[PubMed]
- Huangfu, D. & Anderson, K. V.: Cilia and Hedgehog responsiveness in the mouse. Proc. Natl. Acad. Sci. USA, 102, 11325-11330 (2005)[PubMed]
- Weirich, C. S., Erzberger, J. P., Barral, Y.: The septin family of GTPases: architecture and dynamics. Nat. Rev. Mol. Cell Biol., 9, 478-489 (2008)[PubMed]
- Tooley, A. J., Gilden, J., Jacobelli, J. et al.: Amoeboid T lymphocytes require the septin cytoskeleton for cortical integrity and persistent motility. Nat. Cell Biol., 11, 17-26 (2009)[PubMed]
- Hu, Q., Milenkovic, L., Jin, H. et al.: A septin diffusion barrier at the base of the primary cilium maintains ciliary membrane protein distribution. Science, 329, 436-439 (2010)[PubMed]
- Tobin, J. L. & Beales, P. L.: Bardet-Biedl syndrome: beyond the cilium. Pediatr. Nephrol., 22, 926-936 (2007)[PubMed]
- Park, T. J., Haigo, S. L., Wallingford, J. B.: Ciliogenesis defects in embryos lacking inturned or fuzzy function are associated with failure of planar cell polarity and Hedgehog signaling. Nat. Genet., 38, 303-311 (2006)[PubMed]
著者プロフィール
略歴:2008年 総合研究大学院大学生命科学研究科博士課程 修了,同年 基礎生物学研究所 研究員(上野直人研究室)を経て,2009年より米国Texas大学Austin校 研究員(John Wallingford研究室).
研究テーマ:アフリカツメガエル初期胚における細胞極性の形成機構の解明.
抱負:医学はもっと発生学的な思考と知識を取り込む必要があるのではないかという(漠然とした)意識をもち,獣医学部を卒業したのち,発生学を専門として研究をつづけている.カエルのたまごひとつをとっても解明困難な発生現象がたたみかけるように現れ,目がくらみつつも,ひとつでも何かを発見し解明していくことが人類の未来を開くものと信じて,日々,カエルのたまご相手に奮闘している.
© 2010 進藤 麻子 Licensed under CC 表示 2.1 日本
