局所的なCa2+シグナルは不要な神経突起の選択的な除去を制御する
金森崇浩・榎本和生
(大阪バイオサイエンス研究所 神経細胞生物学部門)
email:金森崇浩,榎本和生
DOI: 10.7875/first.author.2013.080
Compartmentalized calcium transients trigger dendrite pruning in Drosophila sensory neurons.
Takahiro Kanamori, Makoto I. Kanai, Yusuke Dairyo, Kei-ichiro Yasunaga, Rei K. Morikawa, Kazuo Emoto
Science, 340, 1475-1478 (2013)
幼弱期の神経回路は不要な神経接続が数多く存在する“混線”状態にある.神経回路が機能的に成熟するためには,この混線した回路から不要な神経突起が選択的に除去されることが重要である.しかし,ニューロンがいかにして不要な神経突起のみを選択的に除去するのかという問題は,長年にわたり解かれていない謎であった.今回,筆者らは,ショウジョウバエの感覚ニューロンをモデル系として解析することにより,局所的に起こるCa2+濃度の変動が不要な神経突起を時間的および空間的に規定するシグナルとして機能していることを明らかにした.
ニューロンは神経突起を精妙に配線することにより機能的な神経回路を構築する.神経突起が配線される過程は大きく2つの段階に分けられる1).まず,神経突起の伸長およびガイダンスが起こり,過剰な神経接続を含む未熟な神経回路がつくられる.そののち,未熟な神経回路から不要な神経突起のみが除去され,神経回路は機能的に成熟する.たとえば,哺乳類の視覚系において胎生期に生み出された網膜神経節細胞は,発生の初期には内網状層の全体にわたり樹状突起を分岐させるが,そののち,不要な樹状突起を削ぎ落とし,最終的には,内網状層のオン層(もしくは,オフ層)に選択的に樹状突起を分布させる1).このような成熟の過程においてニューロンは神経突起の“要”“不要”を明確に区別し,不要な神経突起のみを選択的に除去していると考えられる.しかし,ネコやフェレットなどを用いたこれまでの解析系においては,不要な神経突起が除去される一部始終をリアルタイムに追跡することは困難であり,神経突起の“要”“不要”を規定するシグナルの実体はとらえられていなかった.
近年,ショウジョウバエが幼虫から蛹へと形態変化する変態期において不要な神経突起が選択的に除去される現象が報告されており2,3),不要な神経突起の除去に必要な遺伝子も複数が同定されている4,5).しかし,それらの作用機構に関する情報はいまだ断片的で,とくに,ニューロンがいかにして不要な神経突起のみを選択的に除去するのかという問題は単純な変異体スクリーニングからは解決されてこなかった.そこで筆者らは,生細胞イメージング技術を駆使して不要な神経突起において活性化されている現象をとらえるという独自のアプローチにより,不要な神経突起の除去を制御する分子機構の解明を試みた.
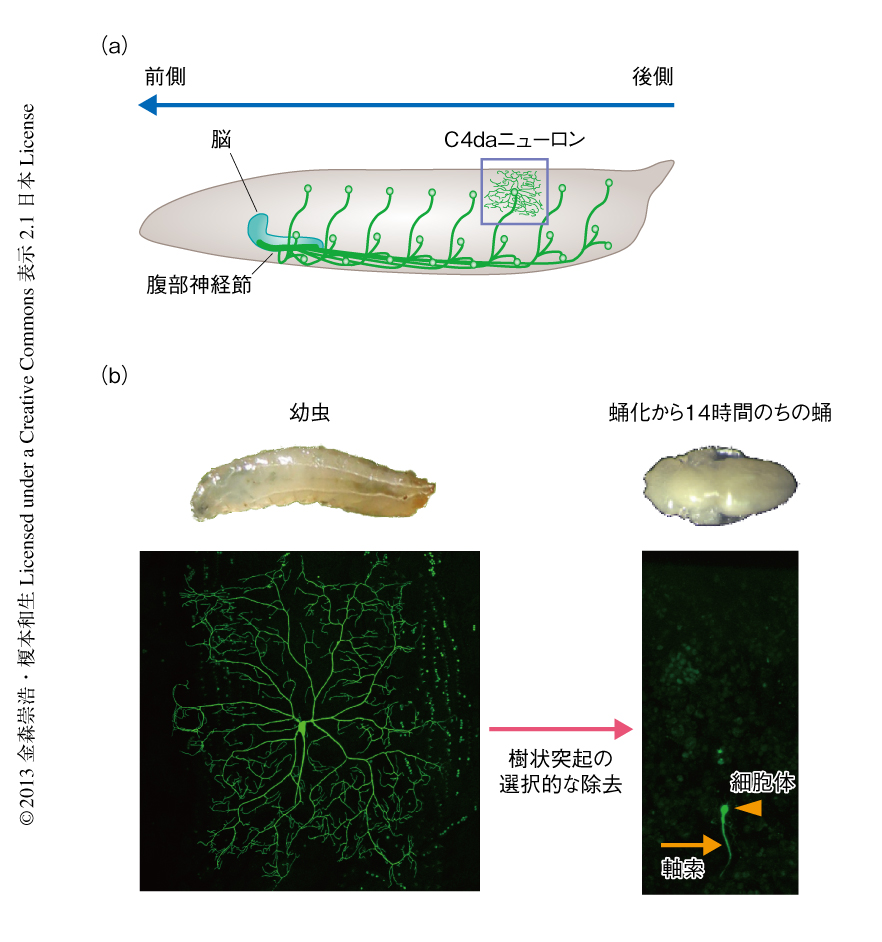
ショウジョウバエの末梢感覚ニューロンのひとつであるクラスIV感覚ニューロン(C4daニューロン)は,幼虫期に発達させた樹状突起を蛹期の前半に除去し,そののち,成虫用の樹状突起を蛹期の後半に再構築する3-5)(図1).樹状突起の除去は蛹化の開始から約14時間で完了するが,そのあいだに,もうひとつの神経突起である軸索は除去されない.したがって,C4daニューロンは神経突起の“要”“不要”を明確に区別し,樹状突起のみを選択的に除去していると考えられる.
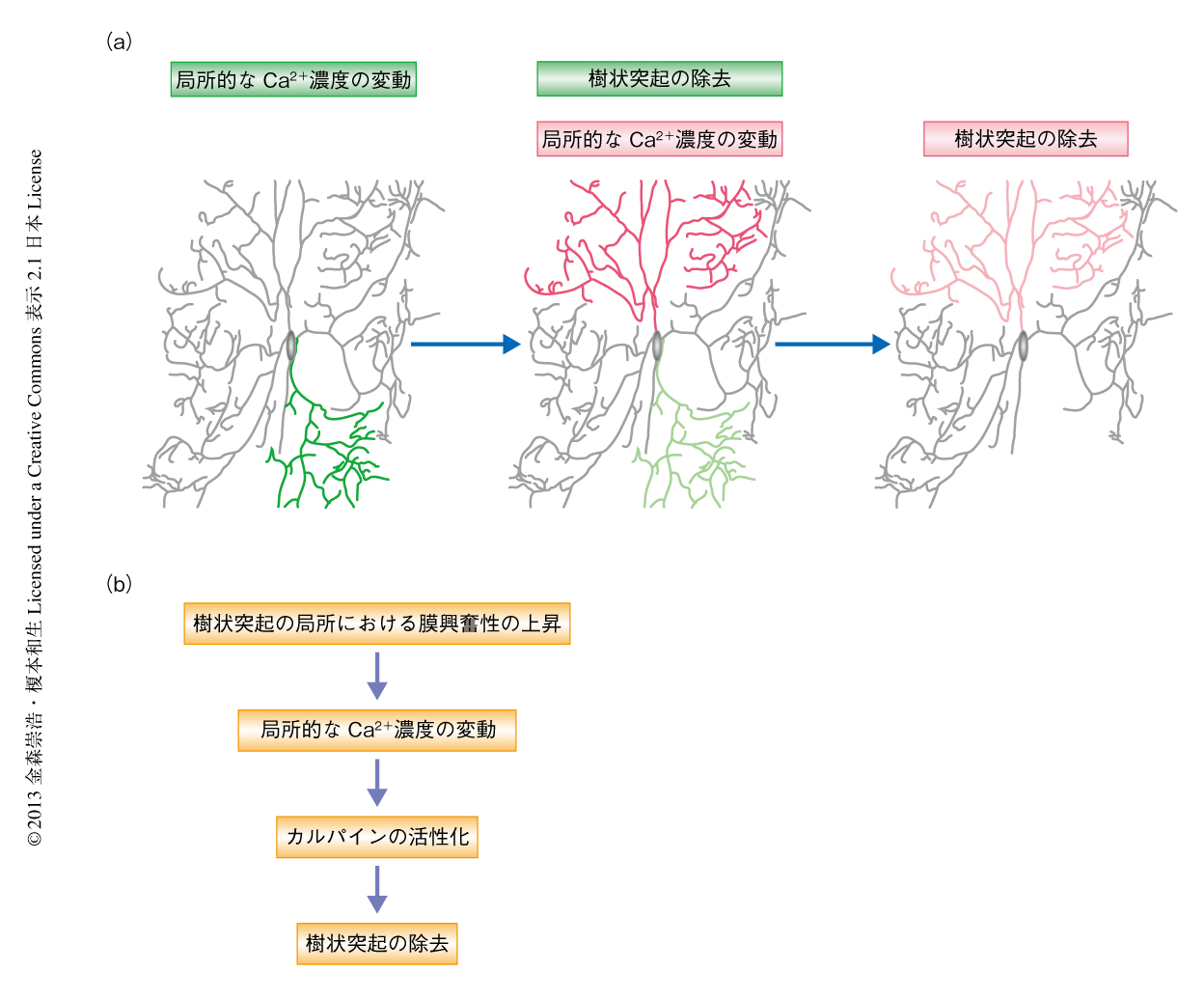
そこで,神経突起の形態制御において多彩な役割をはたすことが知られているCa2+に着目し1),緑色蛍光性Ca2+センサータンパク質GCaMP3を用いて,蛹化の直後から8時間にわたりCa2+イメージングを行った.その結果,一部の樹状突起の集団において,Ca2+濃度の一過性の上昇が1分間に約1回の低頻度で発生しつづけることを見い出した.同様のCa2+濃度の変動は軸索あるいは細胞体においてはまったく観察されなかったことから,樹状突起の選択的な除去となんらかの関連をもつ可能性が考えられた.そこで,樹状突起の形態変化に注意して解析したところ,局所的なCa2+濃度の変動を発生する樹状突起の集団は,発生の開始から2~3時間のちに一斉に除去されることがわかった.さらに,同様の過程は別の樹状突起の集団においてもくり返され,数時間にわたる局所的なCa2+濃度の変動を経験した樹状突起から順番に除去は進行した(図2a).つまり,Ca2+濃度の変動の発生と樹状突起の除去とのあいだには時間的および空間的な相関関係がみられたのである.これらの観察結果から,Ca2+濃度の変動が不要な樹状突起を時間的および空間的に規定するシグナルとしてはたらいている可能性が示唆された.
ニューロンにおけるCa2+の動態は,細胞外からのCa2+流入と細胞内ストアからのCa2+流出という2種類の経路を介し制御されている.そこで,局所的なCa2+濃度の変動を制御するCa2+流入経路を同定するため,C4daニューロンにおいて特定のCa2+チャネルの遺伝子発現を抑制することにより,局所的なCa2+濃度の変動の発生を阻害するようなCa2+チャネルを探索した.その結果,樹状突起の局所においてCa2+濃度の変動が発生するためには,電位依存性Ca2+チャネルを介した細胞外からのCa2+流入が必要であることが明らかになった.
つづいて,樹状突起の選択的な除去が電位依存性Ca2+チャネルの変異体において正常に進行するかどうかを調べることにより,局所的なCa2+濃度の変動が樹状突起の除去に必要であるかどうかを検証した.さきに述べたように,野生型のC4daニューロンの樹状突起は蛹化の開始から14時間で完全に除去される.しかし,Ca2+濃度の変動の発生しない電位依存性Ca2+チャネル変異体のニューロンにおいては樹状突起の除去は起こらず,蛹化ののち14時間の時点において多くの樹状突起が残存していた.また,静止膜電位の過分極作用をもつ内向き整流性K+チャネルであるKir2.1を用いて,電位依存性Ca2+チャネルを介したCa2+流入を蛹期において特異的に阻害した場合にも,同様の樹状突起の除去の異常が観察された.以上の結果から,電位依存性Ca2+チャネルを介し発生する局所的なCa2+濃度の変動は,樹状突起の選択的な除去に必要であることが明らかになった.
細胞内におけるCa2+の機能の発現は,Ca2+に依存して活性制御をうける細胞内のタンパク質を介することが多い.そこで,Ca2+依存性のタンパク質をコードする遺伝子の機能を抑制して樹状突起の除去にあたえる影響を調べた.その結果,Ca2+依存性のタンパク質分解酵素であるカルパインが樹状突起の選択的な除去に必要であることを見い出した.また,カルパインの変異体は電位依存性Ca2+チャネルの変異体との明確な遺伝学的な相互作用を示した.これらの結果からも,樹状突起の除去におけるCa2+濃度の変動の役割が支持された.
それでは,C4daニューロンはいかにして局所的なCa2+濃度の変動をひき起こしているのだろうか? 不要な神経突起が規定されるしくみをより本質的に理解するためには,この点を明らかにすることが重要である.C4daニューロンは外部からの多様な刺激を受容する感覚ニューロンであることから6),毎度のCa2+濃度の変動において外部からのなんらかの刺激が局所的にくわえられているのかもしれない.しかし,感覚ニューロンが外部からの刺激を感知するために重要なイオンチャネルをコードする遺伝子の機能を抑制しても,局所的なCa2+濃度の変動の発生が阻害されることはなかった.したがって,少なくとも現段階では,このような外因的な要因によりCa2+濃度の変動の局所性を説明することはできない.
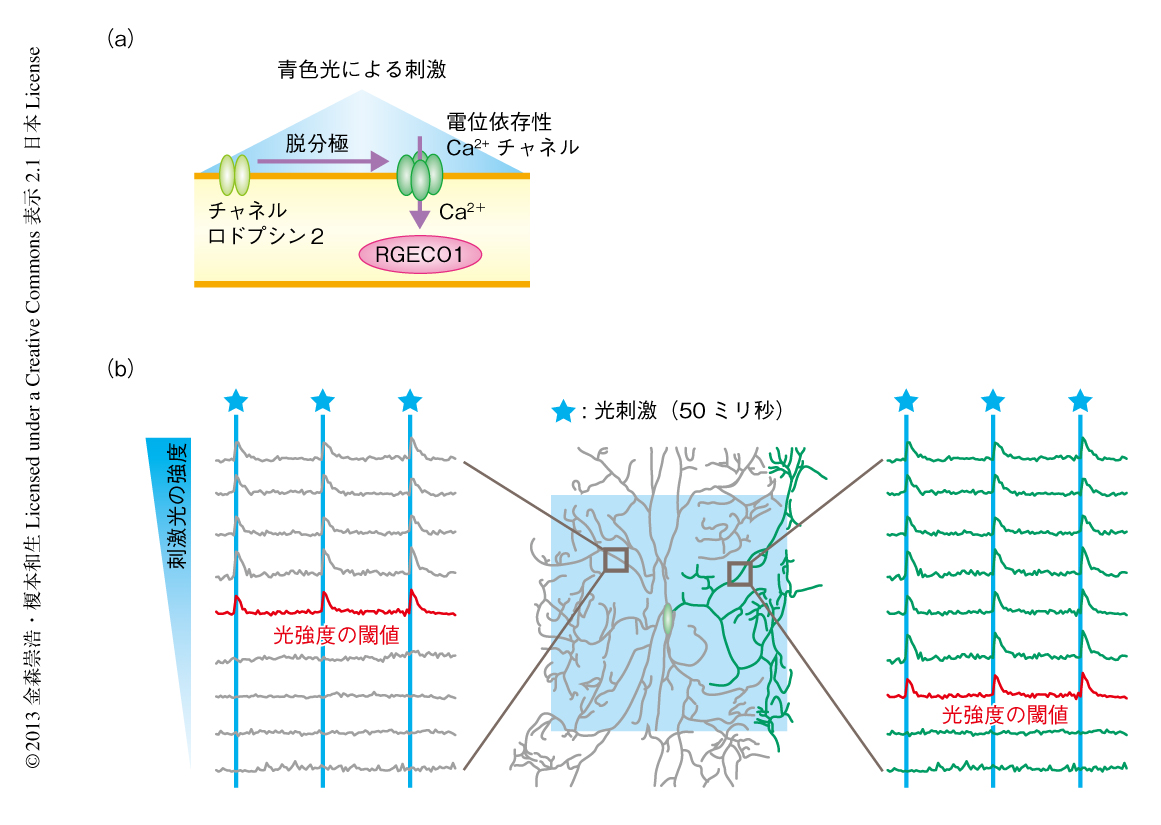
別の可能性として,樹状突起の局所において内因的な膜興奮性が上昇し,電位依存性Ca2+チャネルを介したCa2+流入が起こりやすくなっていることが考えられた.この可能性を検証するため,樹状突起の膜興奮性を定量的に評価できる実験系を構築した(図3a).具体的には,光駆動性カチオンチャネルであるチャネルロドプシン2を発現するC4daニューロンに対し,9段階に光強度の異なる青色光を照射することにより段階的な脱分極刺激をあたえた.これと並行して,赤色蛍光性Ca2+センサータンパク質RGECO1を用いたCa2+イメージングを行い,脱分極刺激により開口した電位依存性Ca2+チャネルを介して流入するCa2+を検出した.この実験系においては,光刺激に呼応したCa2+流入を起こすために必要な光強度の閾値を樹状突起の部位ごとに決定することにより,樹状突起の膜興奮性を定量的および局所的に評価することが可能である.そこで,同一のニューロンにおける樹状突起の膜興奮性を蛹化の直後から30分おきにくり返し調べた.その結果,蛹化の開始から数時間のちに,一部の樹状突起の集団において内因的な膜興奮性が上昇していることが明らかになった(図3b).そののち,膜興奮性を上昇させた樹状突起の集団は局所的なCa2+濃度の変動を開始し,最終的には選択的に除去された.したがって,ニューロンが局所的なCa2+濃度の変動をひき起こすしくみの少なくとも一部は,樹状突起の局所における内因的な膜興奮性の上昇によるものであると考えられた.
筆者らは,今回の研究により,樹状突起の局所において発生するCa2+濃度の変動が不要な樹状突起を時間的および空間的に規定するシグナルとしてはたらき,その選択的な除去を制御することを明らかにした(図2b).この過程において,ニューロンは樹状突起の局所において内因的な膜興奮性を変化させることにより,Ca2+流入を不要な樹状突起に限定しているようであった.カルパインの活性化も,このような膜興奮性の局所的な制御を介して不要な樹状突起に限局されていることが予想された.興味深いことに,樹状突起の局所における膜興奮性の変化は哺乳動物のニューロンが生理機能を発現する際にも利用されている7).したがって,高等動物ですでに報告されている樹状突起の除去において,ショウジョウバエの感覚ニューロンと同様の分子機構が用いられている可能性も大いに考えられる.
ショウジョウバエを用いた解析系の確立された現在,不要な神経突起の除去の研究はようやく緒に就いた段階にあり,筆者らの研究はその一端を明らかにしたにすぎない.不要な神経突起はいかにして膜興奮性を上昇させているのか,除去されない“必要な”突起を規定する分子機構は存在するのかなど,挑戦すべきポイントは豊富に存在する.近年,続々と開発されているイメージング技術を積極的に取り入れることにより,今後も,不要な神経突起の除去についてその分子機構の解明に貢献したい.
略歴:2008年 東京大学大学院薬学系研究科 修了,同年 国立遺伝学研究所 研究員を経て,2010年より大阪バイオサイエンス研究所 研究員.
研究テーマ:ニューロンの可塑的な変化とリモデリング.
抱負:温故知新,無念無想,体力勝負のバランスをとりつつ,自分にしかできないと思えるオリジナルな研究を発信したい.
榎本 和生(Kazuo Emoto)
東京大学大学院理学系研究科 教授.大阪バイオサイエンス研究所 研究部長 兼任.
研究室URL:http://www.biol.s.u-tokyo.ac.jp/users/seiri/index.html
© 2013 金森崇浩・榎本和生 Licensed under CC 表示 2.1 日本
(大阪バイオサイエンス研究所 神経細胞生物学部門)
email:金森崇浩,榎本和生
DOI: 10.7875/first.author.2013.080
Compartmentalized calcium transients trigger dendrite pruning in Drosophila sensory neurons.
Takahiro Kanamori, Makoto I. Kanai, Yusuke Dairyo, Kei-ichiro Yasunaga, Rei K. Morikawa, Kazuo Emoto
Science, 340, 1475-1478 (2013)
要 約
幼弱期の神経回路は不要な神経接続が数多く存在する“混線”状態にある.神経回路が機能的に成熟するためには,この混線した回路から不要な神経突起が選択的に除去されることが重要である.しかし,ニューロンがいかにして不要な神経突起のみを選択的に除去するのかという問題は,長年にわたり解かれていない謎であった.今回,筆者らは,ショウジョウバエの感覚ニューロンをモデル系として解析することにより,局所的に起こるCa2+濃度の変動が不要な神経突起を時間的および空間的に規定するシグナルとして機能していることを明らかにした.
はじめに
ニューロンは神経突起を精妙に配線することにより機能的な神経回路を構築する.神経突起が配線される過程は大きく2つの段階に分けられる1).まず,神経突起の伸長およびガイダンスが起こり,過剰な神経接続を含む未熟な神経回路がつくられる.そののち,未熟な神経回路から不要な神経突起のみが除去され,神経回路は機能的に成熟する.たとえば,哺乳類の視覚系において胎生期に生み出された網膜神経節細胞は,発生の初期には内網状層の全体にわたり樹状突起を分岐させるが,そののち,不要な樹状突起を削ぎ落とし,最終的には,内網状層のオン層(もしくは,オフ層)に選択的に樹状突起を分布させる1).このような成熟の過程においてニューロンは神経突起の“要”“不要”を明確に区別し,不要な神経突起のみを選択的に除去していると考えられる.しかし,ネコやフェレットなどを用いたこれまでの解析系においては,不要な神経突起が除去される一部始終をリアルタイムに追跡することは困難であり,神経突起の“要”“不要”を規定するシグナルの実体はとらえられていなかった.
近年,ショウジョウバエが幼虫から蛹へと形態変化する変態期において不要な神経突起が選択的に除去される現象が報告されており2,3),不要な神経突起の除去に必要な遺伝子も複数が同定されている4,5).しかし,それらの作用機構に関する情報はいまだ断片的で,とくに,ニューロンがいかにして不要な神経突起のみを選択的に除去するのかという問題は単純な変異体スクリーニングからは解決されてこなかった.そこで筆者らは,生細胞イメージング技術を駆使して不要な神経突起において活性化されている現象をとらえるという独自のアプローチにより,不要な神経突起の除去を制御する分子機構の解明を試みた.
1.不要な樹状突起において発生するCa2+濃度の変動
ショウジョウバエの末梢感覚ニューロンのひとつであるクラスIV感覚ニューロン(C4daニューロン)は,幼虫期に発達させた樹状突起を蛹期の前半に除去し,そののち,成虫用の樹状突起を蛹期の後半に再構築する3-5)(図1).樹状突起の除去は蛹化の開始から約14時間で完了するが,そのあいだに,もうひとつの神経突起である軸索は除去されない.したがって,C4daニューロンは神経突起の“要”“不要”を明確に区別し,樹状突起のみを選択的に除去していると考えられる.
そこで,神経突起の形態制御において多彩な役割をはたすことが知られているCa2+に着目し1),緑色蛍光性Ca2+センサータンパク質GCaMP3を用いて,蛹化の直後から8時間にわたりCa2+イメージングを行った.その結果,一部の樹状突起の集団において,Ca2+濃度の一過性の上昇が1分間に約1回の低頻度で発生しつづけることを見い出した.同様のCa2+濃度の変動は軸索あるいは細胞体においてはまったく観察されなかったことから,樹状突起の選択的な除去となんらかの関連をもつ可能性が考えられた.そこで,樹状突起の形態変化に注意して解析したところ,局所的なCa2+濃度の変動を発生する樹状突起の集団は,発生の開始から2~3時間のちに一斉に除去されることがわかった.さらに,同様の過程は別の樹状突起の集団においてもくり返され,数時間にわたる局所的なCa2+濃度の変動を経験した樹状突起から順番に除去は進行した(図2a).つまり,Ca2+濃度の変動の発生と樹状突起の除去とのあいだには時間的および空間的な相関関係がみられたのである.これらの観察結果から,Ca2+濃度の変動が不要な樹状突起を時間的および空間的に規定するシグナルとしてはたらいている可能性が示唆された.
2.局所的なCa2+濃度の変動は樹状突起の選択的な除去に必要である
ニューロンにおけるCa2+の動態は,細胞外からのCa2+流入と細胞内ストアからのCa2+流出という2種類の経路を介し制御されている.そこで,局所的なCa2+濃度の変動を制御するCa2+流入経路を同定するため,C4daニューロンにおいて特定のCa2+チャネルの遺伝子発現を抑制することにより,局所的なCa2+濃度の変動の発生を阻害するようなCa2+チャネルを探索した.その結果,樹状突起の局所においてCa2+濃度の変動が発生するためには,電位依存性Ca2+チャネルを介した細胞外からのCa2+流入が必要であることが明らかになった.
つづいて,樹状突起の選択的な除去が電位依存性Ca2+チャネルの変異体において正常に進行するかどうかを調べることにより,局所的なCa2+濃度の変動が樹状突起の除去に必要であるかどうかを検証した.さきに述べたように,野生型のC4daニューロンの樹状突起は蛹化の開始から14時間で完全に除去される.しかし,Ca2+濃度の変動の発生しない電位依存性Ca2+チャネル変異体のニューロンにおいては樹状突起の除去は起こらず,蛹化ののち14時間の時点において多くの樹状突起が残存していた.また,静止膜電位の過分極作用をもつ内向き整流性K+チャネルであるKir2.1を用いて,電位依存性Ca2+チャネルを介したCa2+流入を蛹期において特異的に阻害した場合にも,同様の樹状突起の除去の異常が観察された.以上の結果から,電位依存性Ca2+チャネルを介し発生する局所的なCa2+濃度の変動は,樹状突起の選択的な除去に必要であることが明らかになった.
細胞内におけるCa2+の機能の発現は,Ca2+に依存して活性制御をうける細胞内のタンパク質を介することが多い.そこで,Ca2+依存性のタンパク質をコードする遺伝子の機能を抑制して樹状突起の除去にあたえる影響を調べた.その結果,Ca2+依存性のタンパク質分解酵素であるカルパインが樹状突起の選択的な除去に必要であることを見い出した.また,カルパインの変異体は電位依存性Ca2+チャネルの変異体との明確な遺伝学的な相互作用を示した.これらの結果からも,樹状突起の除去におけるCa2+濃度の変動の役割が支持された.
3.不要な樹状突起においては内因的な膜興奮性が上昇している
それでは,C4daニューロンはいかにして局所的なCa2+濃度の変動をひき起こしているのだろうか? 不要な神経突起が規定されるしくみをより本質的に理解するためには,この点を明らかにすることが重要である.C4daニューロンは外部からの多様な刺激を受容する感覚ニューロンであることから6),毎度のCa2+濃度の変動において外部からのなんらかの刺激が局所的にくわえられているのかもしれない.しかし,感覚ニューロンが外部からの刺激を感知するために重要なイオンチャネルをコードする遺伝子の機能を抑制しても,局所的なCa2+濃度の変動の発生が阻害されることはなかった.したがって,少なくとも現段階では,このような外因的な要因によりCa2+濃度の変動の局所性を説明することはできない.
別の可能性として,樹状突起の局所において内因的な膜興奮性が上昇し,電位依存性Ca2+チャネルを介したCa2+流入が起こりやすくなっていることが考えられた.この可能性を検証するため,樹状突起の膜興奮性を定量的に評価できる実験系を構築した(図3a).具体的には,光駆動性カチオンチャネルであるチャネルロドプシン2を発現するC4daニューロンに対し,9段階に光強度の異なる青色光を照射することにより段階的な脱分極刺激をあたえた.これと並行して,赤色蛍光性Ca2+センサータンパク質RGECO1を用いたCa2+イメージングを行い,脱分極刺激により開口した電位依存性Ca2+チャネルを介して流入するCa2+を検出した.この実験系においては,光刺激に呼応したCa2+流入を起こすために必要な光強度の閾値を樹状突起の部位ごとに決定することにより,樹状突起の膜興奮性を定量的および局所的に評価することが可能である.そこで,同一のニューロンにおける樹状突起の膜興奮性を蛹化の直後から30分おきにくり返し調べた.その結果,蛹化の開始から数時間のちに,一部の樹状突起の集団において内因的な膜興奮性が上昇していることが明らかになった(図3b).そののち,膜興奮性を上昇させた樹状突起の集団は局所的なCa2+濃度の変動を開始し,最終的には選択的に除去された.したがって,ニューロンが局所的なCa2+濃度の変動をひき起こすしくみの少なくとも一部は,樹状突起の局所における内因的な膜興奮性の上昇によるものであると考えられた.
おわりに
筆者らは,今回の研究により,樹状突起の局所において発生するCa2+濃度の変動が不要な樹状突起を時間的および空間的に規定するシグナルとしてはたらき,その選択的な除去を制御することを明らかにした(図2b).この過程において,ニューロンは樹状突起の局所において内因的な膜興奮性を変化させることにより,Ca2+流入を不要な樹状突起に限定しているようであった.カルパインの活性化も,このような膜興奮性の局所的な制御を介して不要な樹状突起に限局されていることが予想された.興味深いことに,樹状突起の局所における膜興奮性の変化は哺乳動物のニューロンが生理機能を発現する際にも利用されている7).したがって,高等動物ですでに報告されている樹状突起の除去において,ショウジョウバエの感覚ニューロンと同様の分子機構が用いられている可能性も大いに考えられる.
ショウジョウバエを用いた解析系の確立された現在,不要な神経突起の除去の研究はようやく緒に就いた段階にあり,筆者らの研究はその一端を明らかにしたにすぎない.不要な神経突起はいかにして膜興奮性を上昇させているのか,除去されない“必要な”突起を規定する分子機構は存在するのかなど,挑戦すべきポイントは豊富に存在する.近年,続々と開発されているイメージング技術を積極的に取り入れることにより,今後も,不要な神経突起の除去についてその分子機構の解明に貢献したい.
文 献
- Wong, R. O. & Ghosh, A.: Activity-dependent regulation of dendritic growth and patterning. Nat. Rev. Neurosci., 3, 803-812 (2002)[PubMed]
- Luo, L. & O’Leary, D. D.: Axon retraction and degeneration in development and disease. Annu. Rev. Neurosci., 28, 127-156 (2005)[PubMed]
- Williams, D. W. & Truman, J. W.: Remodeling dendrites during insect metamorphosis. J. Neurobiol., 64, 24-33 (2005)[PubMed]
- Kuo, C. T., Jan, L. Y. & Jan, Y. N.: Dendrite-specific remodeling of Drosophila sensory neurons requires matrix metalloproteases, ubiquitin-proteasome, and ecdysone signaling. Proc. Natl. Acad. Sci. USA, 102, 15230-15235 (2005)[PubMed]
- Kirilly, D., Gu, Y., Huang, Y. et al.: A genetic pathway composed of Sox14 and Mical governs severing of dendrites during pruning. Nat. Neurosci., 12, 1497-1505 (2009)[PubMed]
- Zhong, L., Hwang, R. Y. & Tracey, W. D.: Pickpocket is a DEG/ENaC protein required for mechanical nociception in Drosophila larvae. Curr. Biol., 429-434 (2010)[PubMed]
- Losonczy, A., Makara, J. K. & Magee, J. C.: Compartmentalized dendritic plasticity and input feature storage in neurons. Nature, 452, 436-441 (2008)[PubMed]
著者プロフィール
略歴:2008年 東京大学大学院薬学系研究科 修了,同年 国立遺伝学研究所 研究員を経て,2010年より大阪バイオサイエンス研究所 研究員.
研究テーマ:ニューロンの可塑的な変化とリモデリング.
抱負:温故知新,無念無想,体力勝負のバランスをとりつつ,自分にしかできないと思えるオリジナルな研究を発信したい.
榎本 和生(Kazuo Emoto)
東京大学大学院理学系研究科 教授.大阪バイオサイエンス研究所 研究部長 兼任.
研究室URL:http://www.biol.s.u-tokyo.ac.jp/users/seiri/index.html
© 2013 金森崇浩・榎本和生 Licensed under CC 表示 2.1 日本
