プロトカドヘリン17は大脳皮質-大脳基底核神経回路において前シナプスの構築を制御する
星名直祐・山本 雅
(沖縄科学技術大学院大学 細胞シグナルユニット)
email:星名直祐,山本 雅
DOI: 10.7875/first.author.2013.077
Protocadherin 17 regulates presynaptic assembly in topographic corticobasal ganglia circuits.
Naosuke Hoshina, Asami Tanimura, Miwako Yamasaki, Takeshi Inoue, Ryoji Fukabori, Teiko Kuroda, Kazumasa Yokoyama, Tohru Tezuka, Hiroshi Sagara, Shinji Hirano, Hiroshi Kiyonari, Masahiko Takada, Kazuto Kobayashi, Masahiko Watanabe, Masanobu Kano, Takanobu Nakazawa, Tadashi Yamamoto
Neuron, 78, 839-854 (2013)
多様な脳高次機能を担う大脳皮質-大脳基底核神経回路には,並列回路的に情報処理を行う領域に特異的な神経回路の存在が示唆されている.個々の脳機能の発現には神経回路に特異的なシナプス発達の精緻化が必要であるが,分子レベルあるいは細胞レベルにおける大脳皮質-大脳基底核神経回路の制御機構についてはこれまで明らかになっていない.今回,筆者らは,δ2プロトカドヘリンファミリーに属するプロトカドヘリン17が大脳皮質-大脳基底核神経回路にそって領域特異的な発現パターンを示しシナプス部位に局在することを見い出した.プロトカドヘリン17のノックアウトマウスでは大脳皮質-大脳基底核神経回路において前シナプス小胞の集積とシナプス伝達効率の亢進が観察された.さらに,プロトカドヘリン17ノックアウトマウスは大脳皮質-大脳基底核神経回路が関与すると考えられている抑うつ様の行動をとりにくいという表現型を示した.これらの知見から,プロトカドヘリン17は大脳皮質-大脳基底核神経回路における特異的なシナプス発達に関与しており,抑うつの行動と大脳皮質-大脳基底核神経回路におけるプロトカドヘリン17の役割の重要性が示唆された.
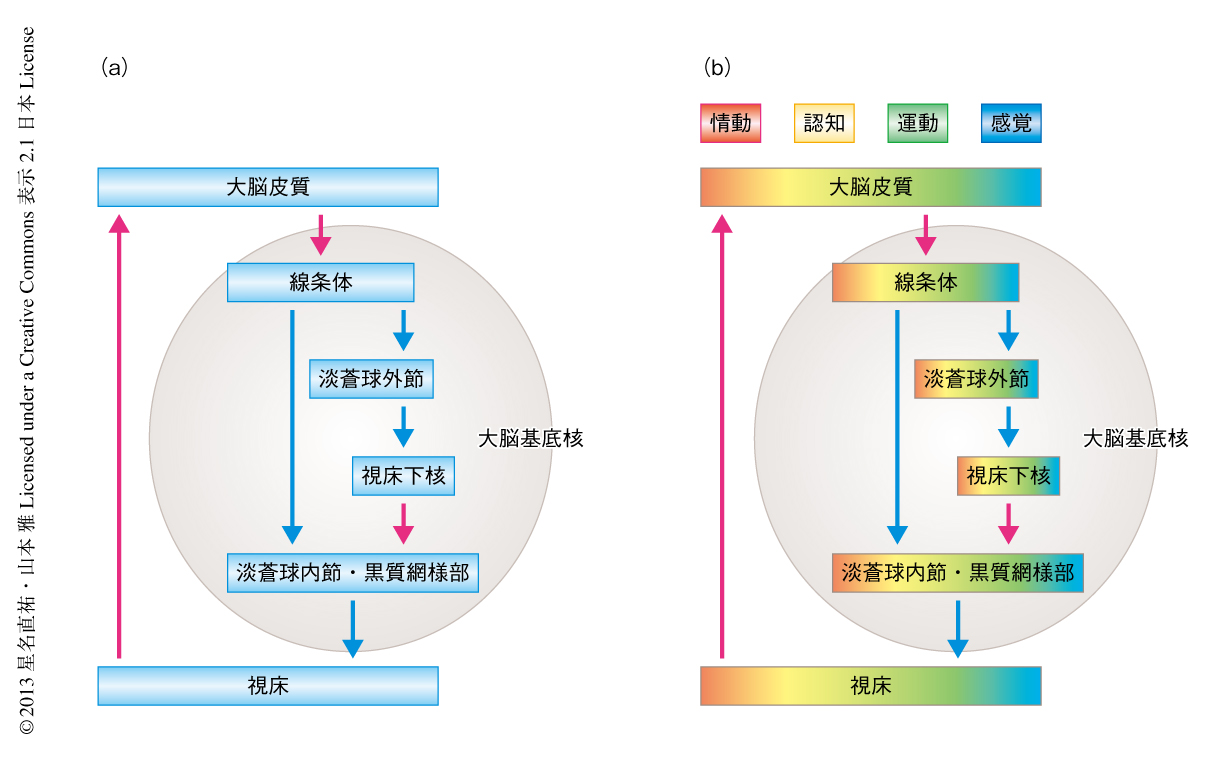
中枢神経系における神経回路は多様なニューロンが正しくシナプス結合することにより構築される.なかでも,大脳基底核を介した神経回路は,運動制御,感覚機能,認知機能,情動,動機づけ,報酬学習など,さまざまな脳高次機能をつかさどる神経回路として位置づけられており,大脳皮質→大脳基底核(線条体→淡蒼球/黒質)→視床というループを形成している(図1a).このような異なる脳機能を実行するため,基底核には並列処理回路の存在することが示唆されており,神経解剖学的に神経回路における位置情報を保持して特異的に形成された“トポグラフィックな”神経回路により具現化されていると考えられている(図1b).さらに,大脳皮質-大脳基底核神経回路の破綻は,パーキンソン病,強迫神経症,統合失調症,うつ病など,さまざまな病的な症状をひき起こすことが知られている.したがって,大脳皮質-大脳基底核神経回路における正確かつ特異的な神経回路の形成と,おのおのの神経回路におけるシナプス制御を解明することは,固有の脳機能の制御機構を理解するだけでなく,神経疾患および精神疾患への臨床応用にもつながると考えられる.
領域特異的な大脳皮質-大脳基底核神経回路の機能的な構築には,神経回路に特異的なシナプス発達とその制御が必要である.近年の研究から,個々の神経回路のシナプス発達にはさまざまなシナプス相互作用タンパク質が重要であることが示唆されている1).なかでも,プロトカドヘリンを含むカドヘリンスーパーファミリーは個々のメンバーが異なるタイプのニューロンに発現しており,シナプスの特異的な相互作用に関与する重要なタンパク質であると考えられている2).さらに,高い結合選択性をもつカドヘリンとその細胞内結合タンパク質であるカテニンとの複合体は,前シナプスおよび後シナプスの発達に必要であることが明らかになっている3).前シナプスにおける役割としては,Nカドヘリンやβカテニンを欠損したニューロンにおいてシナプス小胞の前シナプス末端への集積に異常がみられ,カドヘリン-カテニン複合体がこの過程に重要であることが示唆されている4,5).しかしながら,いくつかのカドヘリンファミリーのメンバーが大脳基底核において発現していることは報告されているものの6),神経回路に特異的なシナプス発達における役割やその生理的な意義については不明であった.そこで,筆者らは,δ2プロトカドヘリンファミリーに属するプロトカドヘリン17に注目し,大脳皮質-大脳基底核神経回路における神経回路に特異的なシナプスの制御機構につき解析した.
プロトカドヘリン17は6個の細胞外カドヘリン様ドメインとユニークな細胞質領域をもつI型膜タンパク質である.プロトカドヘリン17の発現パターンを検討したところ,マウスにおいて脳に特異的に発現しており,シナプス発達期の初期にもっとも発現が高くなることが見い出された.脳における発現パターンについて詳細に検討するためプロトカドヘリン17に対する免疫組織染色を行ったところ,線条体,淡蒼球外節および淡蒼球内節,黒質網様部を含む大脳基底核の神経回路にそって強い発現がみられた.なかでも,線条体においては前方,淡蒼球外節および淡蒼球内節においては内側,さらに,黒質網様部においては後方に発現しており,大脳基底核において領域特異的な発現パターンを示した.PCDH17遺伝子を発現するニューロンについて検討したところ,大脳皮質のニューロンにおいても強く発現していた.その発現パターンは内側前頭前皮質がもっとも高く,帯状皮質や運動野では高い発現,体性感覚野やより後方の大脳皮質では中程度から弱い発現であった.また,これらの脳領域の解剖学的な連結について神経トレーサーを用いて検討した.その結果,内側前頭前皮質は前方線条体と連結しており,前方線条体は淡蒼球外節および淡蒼球内節の内側,黒質網様部の後方と連結していた.したがって,プロトカドヘリン17は内側前頭前皮質→前方線条体→内側淡蒼球/後方黒質網様部という神経回路にそって高く発現しており,いい換えると,大脳皮質-大脳基底核神経回路にそって領域特異的な発現様式を示した.
これまで,δ2プロトカドヘリンファミリーに属するプロトカドヘリン10が線条体において高発現していることが報告されていたことから,プロトカドヘリン17とプロトカドヘリン10との大脳基底核における発現パターンを比較した.その結果,プロトカドヘリン17は前方線条体において発現しているのに対し,プロトカドヘリン10は後方線条体において発現しており,両者の発現は相補的なパターンを示した.淡蒼球外節および淡蒼球内節においては,プロトカドヘリン17は内側に分布するがプロトカドヘリン10は外側に分布し,両者は入れ子状の相補的な発現パターンを示した.さらに,黒質網様部においては,プロトカドヘリン17は後方,プロトカドヘリン10は前方に発現するという相補的なパターンを示した.また,大脳皮質や視床においてもプロトカドヘリン17とプロトカドヘリン10はゆるい相補的な発現パターンを示し,プロトカドヘリン17とプロトカドヘリン10は大脳皮質-大脳基底核-視床というループの全体において,領域特異的な神経回路を反映していることが示唆された.
大脳基底核のニューロンにおけるプロトカドヘリン17の局在について検討するため,免疫電子顕微鏡法により観察した.その結果,大脳皮質からの入力をうける前方線条体の興奮性シナプス,および,線条体の出力神経核のひとつである淡蒼球内節の抑制性シナプスにおいて,プロトカドヘリン17はシナプス周辺部位に多く局在することがわかった.カドヘリンファミリーの多くのメンバーはCa2+に依存的な同種類どうしの相互作用を示すため,プロトカドヘリン17の結合様式について検討した.その結果,プロトカドヘリン17はCa2+に依存的にプロトカドヘリン17どうしで結合するが,プロトカドヘリン10とは結合しないことが明らかになった.したがって,プロトカドヘリン17は大脳基底核における興奮性シナプスと抑制性シナプスの両方において,シナプス周辺部位にてプロトカドヘリン17どうしで結合していることが示唆された.
プロトカドヘリン17の生理学的な役割を解析するため,プロトカドヘリン17のノックアウトマウスを樹立した.組織学的な解析から,プロトカドヘリン17ノックアウトマウスの脳構造に大まかな異常はみられず,大脳基底核における軸索の投射の異常も観察されなかった.また,大脳皮質-大脳基底核神経回路における特異的な領域の形成について検討するため神経トレーサーを用いたところ,プロトカドヘリン17ノックアウトマウスにおいては特異的な神経回路の領域形成に異常はみられなかった.そこで,プロトカドヘリン17ノックアウトマウスの大脳基底核におけるシナプスの形態について検討するため,電子顕微鏡を用いたシナプスの観察を行った.プロトカドヘリン17ノックアウトマウスの前方線条体の興奮性シナプスにおいて,シナプス密度,スパイン形態,シナプス間隙距離,シナプス後肥厚の大きさは野生型マウスと同じ程度であったが,前シナプスにおけるシナプス小胞の数が有意に増加していることが見い出された.一方で,プロトカドヘリン17の発現の低い後方線条体においては前シナプスにおけるシナプス小胞の増加はみられなかった.さらに,線条体の出力神経核として淡蒼球内節における抑制性シナプスの形態の観察も行った.淡蒼球内節においてもプロトカドヘリン17の発現の高い内側の領域ではシナプス小胞の増加が観察されたが,発現の低い外側の領域では野生型マウスと同じ程度であった.以上の結果より,プロトカドヘリン17は大脳基底核のシナプスにおいて,領域特異的にシナプス小胞の集積を制御していることが示唆された.さらに,シナプスの機能についてより詳細に検討するため,大脳皮質-線条体シナプスの電気生理学的な解析を行った.プロトカドヘリン17ノックアウトマウスには,前方線条体のニューロンにおいて微小興奮性シナプス後電流や後シナプスの受容体の特性に異常はみられなかった.そこで,シナプス小胞の動的なプロセスを反映させるため,大脳皮質-線条体シナプスの反復刺激にともなうシナプスの応答を検討した.その結果,野生型マウスでは反復刺激にともないシナプスの応答は減少したが,プロトカドヘリン17ノックアウトマウスではその減少が有意に起こりにくくなっていることが明らかになった.このことより,プロトカドヘリン17ノックアウトマウスでは前シナプス小胞の集積が増加することによりシナプス伝達効率の亢進が起こっていることが示唆された.
プロトカドヘリン17ノックアウトマウスの行動の異常について検討するため,種々の脳機能を評価する行動バッテリー試験を行った.その結果,抑うつ様の行動のモデル試験として用いられる尾懸垂試験と強制水泳試験において,プロトカドヘリン17ノックアウトマウスは野生型マウスに比べ無動時間が有意に減少していることが見い出された.このことは,プロトカドヘリン17ノックアウトマウスがストレス状況においてうつ状態になりにくいことを示唆していた.一方で,プロトカドヘリン17ノックアウトマウスには自発的な行動量,不安行動,恐怖条件づけ,驚愕反応などの異常は観察されず,プロトカドヘリン17は抑うつ様の行動に特異的に関与している可能性が示唆された.
ヒトのうつ病におけるプロトカドヘリン17の機能的な役割について検討するため,霊長類におけるプロトカドヘリン17の発現パターンについて調べた.アカゲザルの脳においてプロトカドヘリン17の免疫組織染色を行ったところ,プロトカドヘリン17は前頭葉と線条体に強く発現していたが,そのパターンは領域特異的であった.前頭葉においては,内側前頭前皮質や一部の前帯状皮質,背外側前頭前皮質において発現が高かった.一方で,ほかの前頭前野の領域や運動野においては中程度に発現をしていたが,体性感覚野やほかの大脳皮質の領域における発現はきわめて低かった.線条体においては尾状核および被殻に発現していたが,前後軸にそって発現の勾配がみられた.淡蒼球や黒質においても領域特異的な発現をしており,大脳皮質-大脳基底核にそった特異的な発現パターンを示した.これらの発現パターンは大部分がマウスと同じであり,マウスと霊長類においてプロトカドヘリン17の発現部位は保存されていたことから,その機能的な役割も保存されている可能性が示唆された.
並列回路的に情報処理を行う領域特異的な神経回路は大脳皮質-大脳基底核神経回路における情報処理に必須の役割をはたしている.筆者らは,プロトカドヘリン17とプロトカドヘリン10が大脳皮質-大脳基底核神経回路において空間的に相補的な発現パターンを示し,領域特異的な神経回路を反映していることを見い出した.また,プロトカドヘリン17ノックアウトマウスでは大脳皮質-大脳基底核神経回路のシナプスにおいてシナプス小胞の集積の増加とシナプス伝達効率の増加が観察され,プロトカドヘリン17は大脳皮質-大脳基底核神経回路において特異的なシナプス発達の精緻化に重要であることが明らかになった(図2).これまでの研究において,プロトカドヘリン10のノックアウトマウスには線条体のニューロンにおける軸索伸長の異常が報告されているが7),大脳皮質-大脳基底核神経回路におけるシナプス部位でのプロトカドヘリン10の機能的な意義はいまだわかっていない.今後,プロトカドヘリン10のコンディショナルノックアウトマウスにより,大脳皮質-大脳基底核神経回路におけるプロトカドヘリン10の役割を明らかにすることが必要であろう.そして,プロトカドヘリン17とプロトカドヘリン10の機能的な役割を対比していくことが,大脳皮質-大脳基底核神経回路における領域特異的な神経回路の全体の意義を理解することに必須であると考えられる.

近年のヒトにおける脳イメージング技術による解析から,内側前頭前皮質と線条体の前方領域とが強く関連しており,これらは実行機能や情動制御に関与するループ回路を形成していると考えられている8).プロトカドヘリン17はげっ歯類および霊長類においてこの回路に対応する領域の発現が高く,ヒトにおいても脳高次機能が保存され機能している可能性が高い.また,プロトカドヘリン17ノックアウトマウスが抑うつ様の行動異常をひき起こしたことから,プロトカドヘリン17の発現が顕著である内側前頭前皮質を介した大脳皮質-大脳基底核神経回路の異常がうつ病において重要である可能性が示唆された.実際,ヒトにおけるうつ病患者の重要な徴候として,内側前頭前皮質と線条体の機能異常が報告されている9).さらに,げっ歯類において内側前頭前皮質を介した特定の神経回路を光遺伝学手法により刺激することにより,抑うつ様の行動を操作できることが示されている10).したがって,今後,プロトカドヘリン17の大脳皮質-大脳基底核における役割とうつ病との関連性を詳細に調べることにより,うつ病の発症機構を理解し,その知見を基盤とした創薬の開発が期待される.
略歴:2009年 東京大学大学院理学系研究科にて博士号取得,東京大学医科学研究所 特任研究員を経て,2011年より沖縄科学技術大学院大学 研究員.
研究テーマ:大脳基底核神経回路.
抱負:大脳皮質-大脳基底核神経回路におけるシナプス発達の機構とその機能的な意義を解明するために,多彩なアプローチを用いた研究を展開したい.また,このような基礎研究をつうじて,大脳皮質-大脳基底核神経回路の破綻により生じる病態の解明にも貢献したい.
山本 雅(Tadashi Yamamoto)
沖縄科学技術大学院大学 教授.
研究室URL:https://groups.oist.jp/ja/csu
© 2013 星名直祐・山本 雅 Licensed under CC 表示 2.1 日本
(沖縄科学技術大学院大学 細胞シグナルユニット)
email:星名直祐,山本 雅
DOI: 10.7875/first.author.2013.077
Protocadherin 17 regulates presynaptic assembly in topographic corticobasal ganglia circuits.
Naosuke Hoshina, Asami Tanimura, Miwako Yamasaki, Takeshi Inoue, Ryoji Fukabori, Teiko Kuroda, Kazumasa Yokoyama, Tohru Tezuka, Hiroshi Sagara, Shinji Hirano, Hiroshi Kiyonari, Masahiko Takada, Kazuto Kobayashi, Masahiko Watanabe, Masanobu Kano, Takanobu Nakazawa, Tadashi Yamamoto
Neuron, 78, 839-854 (2013)
要 約
多様な脳高次機能を担う大脳皮質-大脳基底核神経回路には,並列回路的に情報処理を行う領域に特異的な神経回路の存在が示唆されている.個々の脳機能の発現には神経回路に特異的なシナプス発達の精緻化が必要であるが,分子レベルあるいは細胞レベルにおける大脳皮質-大脳基底核神経回路の制御機構についてはこれまで明らかになっていない.今回,筆者らは,δ2プロトカドヘリンファミリーに属するプロトカドヘリン17が大脳皮質-大脳基底核神経回路にそって領域特異的な発現パターンを示しシナプス部位に局在することを見い出した.プロトカドヘリン17のノックアウトマウスでは大脳皮質-大脳基底核神経回路において前シナプス小胞の集積とシナプス伝達効率の亢進が観察された.さらに,プロトカドヘリン17ノックアウトマウスは大脳皮質-大脳基底核神経回路が関与すると考えられている抑うつ様の行動をとりにくいという表現型を示した.これらの知見から,プロトカドヘリン17は大脳皮質-大脳基底核神経回路における特異的なシナプス発達に関与しており,抑うつの行動と大脳皮質-大脳基底核神経回路におけるプロトカドヘリン17の役割の重要性が示唆された.
はじめに
中枢神経系における神経回路は多様なニューロンが正しくシナプス結合することにより構築される.なかでも,大脳基底核を介した神経回路は,運動制御,感覚機能,認知機能,情動,動機づけ,報酬学習など,さまざまな脳高次機能をつかさどる神経回路として位置づけられており,大脳皮質→大脳基底核(線条体→淡蒼球/黒質)→視床というループを形成している(図1a).このような異なる脳機能を実行するため,基底核には並列処理回路の存在することが示唆されており,神経解剖学的に神経回路における位置情報を保持して特異的に形成された“トポグラフィックな”神経回路により具現化されていると考えられている(図1b).さらに,大脳皮質-大脳基底核神経回路の破綻は,パーキンソン病,強迫神経症,統合失調症,うつ病など,さまざまな病的な症状をひき起こすことが知られている.したがって,大脳皮質-大脳基底核神経回路における正確かつ特異的な神経回路の形成と,おのおのの神経回路におけるシナプス制御を解明することは,固有の脳機能の制御機構を理解するだけでなく,神経疾患および精神疾患への臨床応用にもつながると考えられる.
領域特異的な大脳皮質-大脳基底核神経回路の機能的な構築には,神経回路に特異的なシナプス発達とその制御が必要である.近年の研究から,個々の神経回路のシナプス発達にはさまざまなシナプス相互作用タンパク質が重要であることが示唆されている1).なかでも,プロトカドヘリンを含むカドヘリンスーパーファミリーは個々のメンバーが異なるタイプのニューロンに発現しており,シナプスの特異的な相互作用に関与する重要なタンパク質であると考えられている2).さらに,高い結合選択性をもつカドヘリンとその細胞内結合タンパク質であるカテニンとの複合体は,前シナプスおよび後シナプスの発達に必要であることが明らかになっている3).前シナプスにおける役割としては,Nカドヘリンやβカテニンを欠損したニューロンにおいてシナプス小胞の前シナプス末端への集積に異常がみられ,カドヘリン-カテニン複合体がこの過程に重要であることが示唆されている4,5).しかしながら,いくつかのカドヘリンファミリーのメンバーが大脳基底核において発現していることは報告されているものの6),神経回路に特異的なシナプス発達における役割やその生理的な意義については不明であった.そこで,筆者らは,δ2プロトカドヘリンファミリーに属するプロトカドヘリン17に注目し,大脳皮質-大脳基底核神経回路における神経回路に特異的なシナプスの制御機構につき解析した.
1.プロトカドヘリン17は大脳皮質-大脳基底核神経回路にそって領域特異的な発現様式を示す
プロトカドヘリン17は6個の細胞外カドヘリン様ドメインとユニークな細胞質領域をもつI型膜タンパク質である.プロトカドヘリン17の発現パターンを検討したところ,マウスにおいて脳に特異的に発現しており,シナプス発達期の初期にもっとも発現が高くなることが見い出された.脳における発現パターンについて詳細に検討するためプロトカドヘリン17に対する免疫組織染色を行ったところ,線条体,淡蒼球外節および淡蒼球内節,黒質網様部を含む大脳基底核の神経回路にそって強い発現がみられた.なかでも,線条体においては前方,淡蒼球外節および淡蒼球内節においては内側,さらに,黒質網様部においては後方に発現しており,大脳基底核において領域特異的な発現パターンを示した.PCDH17遺伝子を発現するニューロンについて検討したところ,大脳皮質のニューロンにおいても強く発現していた.その発現パターンは内側前頭前皮質がもっとも高く,帯状皮質や運動野では高い発現,体性感覚野やより後方の大脳皮質では中程度から弱い発現であった.また,これらの脳領域の解剖学的な連結について神経トレーサーを用いて検討した.その結果,内側前頭前皮質は前方線条体と連結しており,前方線条体は淡蒼球外節および淡蒼球内節の内側,黒質網様部の後方と連結していた.したがって,プロトカドヘリン17は内側前頭前皮質→前方線条体→内側淡蒼球/後方黒質網様部という神経回路にそって高く発現しており,いい換えると,大脳皮質-大脳基底核神経回路にそって領域特異的な発現様式を示した.
これまで,δ2プロトカドヘリンファミリーに属するプロトカドヘリン10が線条体において高発現していることが報告されていたことから,プロトカドヘリン17とプロトカドヘリン10との大脳基底核における発現パターンを比較した.その結果,プロトカドヘリン17は前方線条体において発現しているのに対し,プロトカドヘリン10は後方線条体において発現しており,両者の発現は相補的なパターンを示した.淡蒼球外節および淡蒼球内節においては,プロトカドヘリン17は内側に分布するがプロトカドヘリン10は外側に分布し,両者は入れ子状の相補的な発現パターンを示した.さらに,黒質網様部においては,プロトカドヘリン17は後方,プロトカドヘリン10は前方に発現するという相補的なパターンを示した.また,大脳皮質や視床においてもプロトカドヘリン17とプロトカドヘリン10はゆるい相補的な発現パターンを示し,プロトカドヘリン17とプロトカドヘリン10は大脳皮質-大脳基底核-視床というループの全体において,領域特異的な神経回路を反映していることが示唆された.
2.プロトカドヘリン17は大脳皮質-大脳基底核神経回路において前シナプスの構築を制御する
大脳基底核のニューロンにおけるプロトカドヘリン17の局在について検討するため,免疫電子顕微鏡法により観察した.その結果,大脳皮質からの入力をうける前方線条体の興奮性シナプス,および,線条体の出力神経核のひとつである淡蒼球内節の抑制性シナプスにおいて,プロトカドヘリン17はシナプス周辺部位に多く局在することがわかった.カドヘリンファミリーの多くのメンバーはCa2+に依存的な同種類どうしの相互作用を示すため,プロトカドヘリン17の結合様式について検討した.その結果,プロトカドヘリン17はCa2+に依存的にプロトカドヘリン17どうしで結合するが,プロトカドヘリン10とは結合しないことが明らかになった.したがって,プロトカドヘリン17は大脳基底核における興奮性シナプスと抑制性シナプスの両方において,シナプス周辺部位にてプロトカドヘリン17どうしで結合していることが示唆された.
プロトカドヘリン17の生理学的な役割を解析するため,プロトカドヘリン17のノックアウトマウスを樹立した.組織学的な解析から,プロトカドヘリン17ノックアウトマウスの脳構造に大まかな異常はみられず,大脳基底核における軸索の投射の異常も観察されなかった.また,大脳皮質-大脳基底核神経回路における特異的な領域の形成について検討するため神経トレーサーを用いたところ,プロトカドヘリン17ノックアウトマウスにおいては特異的な神経回路の領域形成に異常はみられなかった.そこで,プロトカドヘリン17ノックアウトマウスの大脳基底核におけるシナプスの形態について検討するため,電子顕微鏡を用いたシナプスの観察を行った.プロトカドヘリン17ノックアウトマウスの前方線条体の興奮性シナプスにおいて,シナプス密度,スパイン形態,シナプス間隙距離,シナプス後肥厚の大きさは野生型マウスと同じ程度であったが,前シナプスにおけるシナプス小胞の数が有意に増加していることが見い出された.一方で,プロトカドヘリン17の発現の低い後方線条体においては前シナプスにおけるシナプス小胞の増加はみられなかった.さらに,線条体の出力神経核として淡蒼球内節における抑制性シナプスの形態の観察も行った.淡蒼球内節においてもプロトカドヘリン17の発現の高い内側の領域ではシナプス小胞の増加が観察されたが,発現の低い外側の領域では野生型マウスと同じ程度であった.以上の結果より,プロトカドヘリン17は大脳基底核のシナプスにおいて,領域特異的にシナプス小胞の集積を制御していることが示唆された.さらに,シナプスの機能についてより詳細に検討するため,大脳皮質-線条体シナプスの電気生理学的な解析を行った.プロトカドヘリン17ノックアウトマウスには,前方線条体のニューロンにおいて微小興奮性シナプス後電流や後シナプスの受容体の特性に異常はみられなかった.そこで,シナプス小胞の動的なプロセスを反映させるため,大脳皮質-線条体シナプスの反復刺激にともなうシナプスの応答を検討した.その結果,野生型マウスでは反復刺激にともないシナプスの応答は減少したが,プロトカドヘリン17ノックアウトマウスではその減少が有意に起こりにくくなっていることが明らかになった.このことより,プロトカドヘリン17ノックアウトマウスでは前シナプス小胞の集積が増加することによりシナプス伝達効率の亢進が起こっていることが示唆された.
3.プロトカドヘリン17は抑うつ様の行動に関与する
プロトカドヘリン17ノックアウトマウスの行動の異常について検討するため,種々の脳機能を評価する行動バッテリー試験を行った.その結果,抑うつ様の行動のモデル試験として用いられる尾懸垂試験と強制水泳試験において,プロトカドヘリン17ノックアウトマウスは野生型マウスに比べ無動時間が有意に減少していることが見い出された.このことは,プロトカドヘリン17ノックアウトマウスがストレス状況においてうつ状態になりにくいことを示唆していた.一方で,プロトカドヘリン17ノックアウトマウスには自発的な行動量,不安行動,恐怖条件づけ,驚愕反応などの異常は観察されず,プロトカドヘリン17は抑うつ様の行動に特異的に関与している可能性が示唆された.
ヒトのうつ病におけるプロトカドヘリン17の機能的な役割について検討するため,霊長類におけるプロトカドヘリン17の発現パターンについて調べた.アカゲザルの脳においてプロトカドヘリン17の免疫組織染色を行ったところ,プロトカドヘリン17は前頭葉と線条体に強く発現していたが,そのパターンは領域特異的であった.前頭葉においては,内側前頭前皮質や一部の前帯状皮質,背外側前頭前皮質において発現が高かった.一方で,ほかの前頭前野の領域や運動野においては中程度に発現をしていたが,体性感覚野やほかの大脳皮質の領域における発現はきわめて低かった.線条体においては尾状核および被殻に発現していたが,前後軸にそって発現の勾配がみられた.淡蒼球や黒質においても領域特異的な発現をしており,大脳皮質-大脳基底核にそった特異的な発現パターンを示した.これらの発現パターンは大部分がマウスと同じであり,マウスと霊長類においてプロトカドヘリン17の発現部位は保存されていたことから,その機能的な役割も保存されている可能性が示唆された.
おわりに
並列回路的に情報処理を行う領域特異的な神経回路は大脳皮質-大脳基底核神経回路における情報処理に必須の役割をはたしている.筆者らは,プロトカドヘリン17とプロトカドヘリン10が大脳皮質-大脳基底核神経回路において空間的に相補的な発現パターンを示し,領域特異的な神経回路を反映していることを見い出した.また,プロトカドヘリン17ノックアウトマウスでは大脳皮質-大脳基底核神経回路のシナプスにおいてシナプス小胞の集積の増加とシナプス伝達効率の増加が観察され,プロトカドヘリン17は大脳皮質-大脳基底核神経回路において特異的なシナプス発達の精緻化に重要であることが明らかになった(図2).これまでの研究において,プロトカドヘリン10のノックアウトマウスには線条体のニューロンにおける軸索伸長の異常が報告されているが7),大脳皮質-大脳基底核神経回路におけるシナプス部位でのプロトカドヘリン10の機能的な意義はいまだわかっていない.今後,プロトカドヘリン10のコンディショナルノックアウトマウスにより,大脳皮質-大脳基底核神経回路におけるプロトカドヘリン10の役割を明らかにすることが必要であろう.そして,プロトカドヘリン17とプロトカドヘリン10の機能的な役割を対比していくことが,大脳皮質-大脳基底核神経回路における領域特異的な神経回路の全体の意義を理解することに必須であると考えられる.
近年のヒトにおける脳イメージング技術による解析から,内側前頭前皮質と線条体の前方領域とが強く関連しており,これらは実行機能や情動制御に関与するループ回路を形成していると考えられている8).プロトカドヘリン17はげっ歯類および霊長類においてこの回路に対応する領域の発現が高く,ヒトにおいても脳高次機能が保存され機能している可能性が高い.また,プロトカドヘリン17ノックアウトマウスが抑うつ様の行動異常をひき起こしたことから,プロトカドヘリン17の発現が顕著である内側前頭前皮質を介した大脳皮質-大脳基底核神経回路の異常がうつ病において重要である可能性が示唆された.実際,ヒトにおけるうつ病患者の重要な徴候として,内側前頭前皮質と線条体の機能異常が報告されている9).さらに,げっ歯類において内側前頭前皮質を介した特定の神経回路を光遺伝学手法により刺激することにより,抑うつ様の行動を操作できることが示されている10).したがって,今後,プロトカドヘリン17の大脳皮質-大脳基底核における役割とうつ病との関連性を詳細に調べることにより,うつ病の発症機構を理解し,その知見を基盤とした創薬の開発が期待される.
文 献
- Williams, M. E., de Wit, J., Ghosh, A.: Molecular mechanisms of synaptic specificity in developing neural circuits. Neuron, 68, 9-18 (2010)[PubMed]
- Shapiro, L. & Colman, D. R.: The diversity of cadherins and implications for a synaptic adhesive code in the CNS. Neuron, 23, 427-430 (1999)[PubMed]
- Arikkath, J. & Reichardt, L. F.: Cadherins and catenins at synapses: roles in synaptogenesis and synaptic plasticity. Trends Neurosci., 31, 487-494 (2008)[PubMed]
- Bamji, S. X., Shimazu, K., Kimes, N. et al.: Role of β-catenin in synaptic vesicle localization and presynaptic assembly. Neuron, 40, 719-731 (2003)[PubMed]
- Stan, A., Pielarski, K. N., Brigadski, T. et al.: Essential cooperation of N-cadherin and neuroligin-1 in the transsynaptic control of vesicle accumulation. Proc. Natl. Acad. Sci. USA, 107, 11116-11121 (2010)[PubMed]
- Hertel, N., Krishna-K, Nuernberger, M. et al.: A cadherin-based code for the divisions of the mouse basal ganglia. J. Comp. Neurol., 508, 511-528 (2008)[PubMed]
- Uemura, M., Nakao, S., Suzuki, S. T. et al.: OL-Protocadherin is essential for growth of striatal axons and thalamocortical projections. Nat. Neurosci., 10, 1151-1159 (2007)[PubMed]
- Draganski, B., Kherif, F., Kloppel, S. et al.: Evidence for segregated and integrative connectivity patterns in the human Basal Ganglia. J. Neurosci., 28, 7143-7152 (2008)[PubMed]
- Price, J. L. & Drevets, W. C.: Neural circuits underlying the pathophysiology of mood disorders. Trends Cogn. Sci., 16, 61-71 (2012)[PubMed]
- Warden, M. R., Selimbeyoglu, A., Mirzabekov, J. J. et al.: A prefrontal cortex-brainstem neuronal projection that controls response to behavioural challenge. Nature, 492, 428-432 (2012)[PubMed]
著者プロフィール
略歴:2009年 東京大学大学院理学系研究科にて博士号取得,東京大学医科学研究所 特任研究員を経て,2011年より沖縄科学技術大学院大学 研究員.
研究テーマ:大脳基底核神経回路.
抱負:大脳皮質-大脳基底核神経回路におけるシナプス発達の機構とその機能的な意義を解明するために,多彩なアプローチを用いた研究を展開したい.また,このような基礎研究をつうじて,大脳皮質-大脳基底核神経回路の破綻により生じる病態の解明にも貢献したい.
山本 雅(Tadashi Yamamoto)
沖縄科学技術大学院大学 教授.
研究室URL:https://groups.oist.jp/ja/csu
© 2013 星名直祐・山本 雅 Licensed under CC 表示 2.1 日本
