インドール酢酸-グルコース加水分解酵素の欠失はイネの粒重と収量を増加させる
石丸 健1・廣津直樹2
(1農業生物資源研究所植物科学研究領域 植物生産生理機能研究ユニット,2東洋大学生命科学部 生命科学科植物ゲノム科学研究室)
email:石丸 健,廣津直樹
DOI: 10.7875/first.author.2013.060
Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield.
Ken Ishimaru, Naoki Hirotsu, Yuka Madoka, Naomi Murakami, Nao Hara, Haruko Onodera, Takayuki Kashiwagi, Kazuhiro Ujiie, Bun-ichi Shimizu, Atsuko Onishi, Hisashi Miyagawa, Etsuko Katoh
Nature Genetics, 45, 707-711 (2013)
急速な人口の増加をささえるため,世界の人口の半数以上の主食であるイネの収量を飛躍的に高めることが求められている.粒重はイネの収量性を決定する主要な要因である.筆者らは,インドのイネ在来種であるカサラスからTGW6遺伝子をクローニングしその機能を明らかにすることに成功した.この遺伝子は,インドール酢酸-グルコースの加水分解活性をもつ新規なタンパク質をコードしていた.イネの品種のひとつ日本晴において,tgw6により合成されたインドール酢酸はシンクおよびソースにおいて抑制的に作用した.一方で,機能欠失型変異をもつカサラス型のTGW6はインドール酢酸を合成できず抑制は起こらなかった.そのため,シンクでは胚乳細胞の数が増加し粒長が伸長した.それにくわえて,ソース能が向上することにより粒重ならびに収量も増加した.また,遺伝子の多様性解析の結果から,TGW6遺伝子は栽培化の過程で忘れさられた“もったいない”遺伝子であることが明らかにされた.これらのことより,TGW6遺伝子は近代あるいは現代の栽培品種における収量の改良に広く利用可能であると考えられた.
現在,栽培されているイネは,長いあいだ人の手による選抜をくり返すことによりつくられてきた.したがって,近代あるいは現代の栽培品種は多くの有用遺伝子が集積したものということができる.そのために遺伝的な多様性は失われ,ある品種から有用な遺伝子を特定したとしても,ほかの品種もすでにその遺伝子をもっている可能性が高く,遺伝子の利用範囲は限定されてしまっている.ごくかぎられた品種しかもたない有用な遺伝子をみつけることができれば,多くの品種の改良に利用することができる.栽培化の過程では,種子のサイズなどのシンクの拡大をもとに収量性は増大してきたと考えられている.実際,イネの種子サイズにかかわるいくつかの遺伝子がクローニングされたが,これらの遺伝子は栽培化の過程において選抜に用いられていた.
近代品種ではシンクへの炭水化物の供給能力(ソース能)が収量性を規定する要因となっている.しかしながら,ソース能を制御する遺伝子や分子機構についての情報はこれまでほとんど得られていない.出穂のまえに植物に蓄積された炭水化物は,出穂ののち種子に供給される.そのため,出穂前の炭水化物の蓄積量を増加させることがイネのソース能の改良にむけた主要な育種目標とされてきた.筆者らは,イネの品種のひとつ日本晴とインドのイネ在来種であるカサラスとのあいだの戻し交雑自殖系統を用いて,粒重を増加させる量的形質遺伝子座qTGW6を同定した1).カサラス型のqTGW6遺伝子座を導入した準同質遺伝子系統NIL (TGW6) の解析の結果,qTGW6遺伝子座は出穂前の炭水化物の蓄積量を増加させ,その結果,収量を約15%増加させた.そこで,イネの収量性の向上にむけた知見を得ることを目的に,TGW6遺伝子をクローニングしその機能解析を行った.
ホモ接合型の組換え個体を用いて高密度マッピングを行った結果,TGW6遺伝子の存在する領域を4.9 kbpに絞り込んだ.この領域には1個のエキソンをもつ遺伝子Os06g0623700のみが存在していた.日本晴型tgw6遺伝子と比較して,カサラス型TGW6遺伝子には6個のSNP(single nucleotide polymorphism,一塩基多型)と313番目の塩基の欠失が存在していた.この欠失は終止コドンを生じさせ正常なタンパク質の合成を阻害し機能欠失をひき起こすと考えられた.一方で,6個のSNPはコードされるタンパク質の特性に影響を及ぼさないと予想された.相補試験として,日本晴およびNIL (TGW6) にtgw6遺伝子のRNAiコンストラクトを導入した組換え体を作出して影響を解析した.日本晴のRNAi形質転換体の粒長や粒重は対照と比べ有意に増加した.RNAi形質転換体においてtgw6遺伝子の発現量と粒長とのあいだには負の相関関係がみられた.一方で,NIL (TGW6) のRNAi形質転換体では粒長に変化はみられなかった.さらに,日本晴型tgw6遺伝子をカサラス型のプロモーター配列に連結してNIL (TGW6) に導入したところ粒長と粒重が有意に減少した.これらの結果から,TGW6遺伝子がqTGW6遺伝子座の原因遺伝子であり,この313番目の1塩基欠失が対立遺伝子の多型を決定する機能的な変異であると結論した.
TGW6遺伝子はシンクとソースの両方の器官において発現していた.胚乳におけるTGW6遺伝子の発現量は,受精2日後にピークに達し,受精5日後にはピーク値の約30%まで減少した.TGW6遺伝子は細胞分裂の起こっている胚乳の周辺部で強く発現していた.日本晴と比較してNIL (TGW6) では胚乳細胞の数が約24%増加していた.胚乳細胞の形成の初期段階では細胞分裂は一時的に停止し多核体が形成され,この期間の長さが細胞の数を決定する.胚乳細胞の形成の初期段階において,NIL (TGW6) では細胞分裂に関与する遺伝子の発現レベルが対照に比べ低かったことから,カサラス型TGW6遺伝子が細胞分裂への移行を遅らせ細胞の数を増加させていると考えられた.
Conserved Domain DatabaseのRPS-BLAST解析により日本晴型のtgw6と有意な相同性のみられたストリクトシジンシンターゼおよびジイソプロピルフルオロホスファターゼとの立体構造を比較したところ,ジイソプロピルフルオロホスファターゼのCa2+結合残基はtgw6においても保存されていた.さらに,tgw6の基質ポケットにおけるHis192とTyr224の空間的な配置が加水分解酵素の活性部位と類似していたことから,tgw6は加水分解能をもつことが予測された.さらに,立体構造モデルとの結合シミュレーションによりインドール酢酸-グルコースが基質である可能性が示された.実際に,日本晴型のtgw6はインドール酢酸-グルコースを加水分解したが,カサラス型のTGW6はその活性をもっていなかった.NIL (TGW6) の受精3日後の胚乳において,インドール酢酸の含量は日本晴のわずか16.6%であり,ほとんどのオーキシン応答性遺伝子の発現レベルは低かった.また,NIL (TGW6) の受精の直後の胚珠にインドール酢酸を処理すると,受精5日後の胚乳細胞の長さが減少した.以上の結果から,tgw6によりインドール酢酸-グルコースの加水分解からつくられたインドール酢酸が細胞分裂をひき起こすことにより,胚乳細胞の数や粒長を抑制していると考えられた.そのため,機能欠失型変異をもつカサラス型のTGW6はインドール酢酸を合成できないため抑制作用がはたらかず,その結果,粒長が増加したものと考えられた.
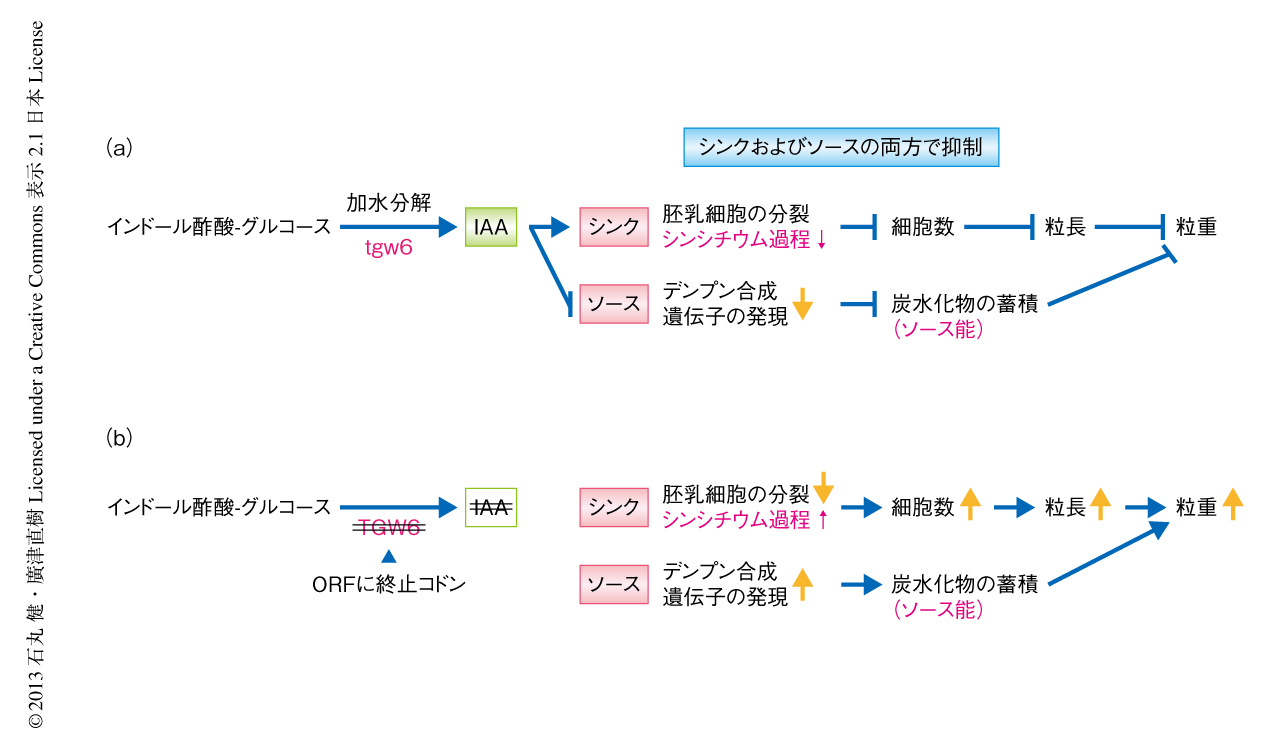
NIL (TGW6) において,出穂のまえに炭水化物の蓄積をおもに担う葉鞘での炭水化物の蓄積量は日本晴よりも多かった.また,NIL (TGW6) の出穂前の葉鞘にインドール酢酸を処理すると,デンプン合成関連遺伝子の発現レベルが低下した.つまり,日本晴型tgw6はインドール酢酸の合成を介して,シンクのみならずソースにおいても抑制的に作用するものと考えられた.また,葉を切除して炭水化物の供給量を減少させたNIL (TGW6) では,粒長は変わらないにもかかわらず粒重の有意な低下が観察された.これらの結果から,カサラス型TGW6遺伝子はソースとシンクにおいて多面的に作用し,そのことが日本晴という現代品種の収量をひとつの遺伝子により改良することのできる要因であると考えられた(図1).
TGW6遺伝子と同様に機能欠失型変異により種子のサイズの増加することは,GW2遺伝子2) やGS3遺伝子3),SW5遺伝子4) でも報告されており,イネにおいて種子のサイズを制限するなんらかの機構の存在する可能性が示唆された.ただし,TGW6遺伝子とは対照的に,これまで報告されている種子サイズを制御する遺伝子(GW2遺伝子,GS3遺伝子,SW5遺伝子,GS5遺伝子5) やGW8遺伝子6) など)は籾のサイズの制御をつうじて間接的に種子に作用する.また,ひとつの形質の改良による収量性の改良は限定的である.たとえば,シンクのサイズが収量を規定している品種では粒のサイズの改良は有効であるが,ボトルネックがほかに移り収量を規定する7).イネの収量を飛躍的に増加させるためには,シンクサイズのみならず炭水化物の生産,転流および蓄積能を並行して改良していく必要がある.
国立遺伝研究所の野生イネ(Oryza rufipogon)コアコレクション8) や農業生物資源研究所が作出した世界イネコアコレクション9) を用いて,遺伝子の多様性解析を行った(図2).その結果,TGW6遺伝子のORFには17のハプロタイプの存在することが確認された.カサラス型TGW6遺伝子に存在する機能的な変異をもつものは,野生イネ1系統(O. perennis W1690)とインドシナ地方原産のイネ4品種のみであった.カサラス型TGW6遺伝子のもつ機能的な変異のほか,自然変異には8個のSNPが存在していた.そのうち4個のSNPはアミノ酸置換を生じさせるが,置換されるアミノ酸残基は活性部位から遠くはなれていた.そのため,これらのSNPはタンパク質の構造ひいては特性にほとんど影響を及ぼさないものと考えられた.実際,イネの品種のひとつIR64のtgw6には,313番目の塩基の1塩基欠失は含まれないがカサラスのもつSNPのうち6個(うち,3個がアミノ酸置換をひき起こす)が含まれていた.IR64型のtgw6遺伝子を導入した準同質遺伝子系統では粒長に変化はみられなかった.この結果は,313番目の1塩基欠失がカサラス型TGW6遺伝子のもつ機能的な変異であることを強く裏づけた.これらの結果により,TGW6遺伝子は栽培化の過程において選抜の対象とされずに捨てさられた“もったいない”遺伝子であると考えられた.事実,コシヒカリやNERICAなどのさまざまな現代品種は,カサラス型TGW6遺伝子のもつ機能的な変異をもたない.カサラス型TGW6遺伝子をコシヒカリに導入した準同質遺伝子系統では籾の大きさは変化せずに粒長や粒重が増加し,単位面積あたりの収量は有意に増加した.これらのことから,TGW6遺伝子は広い遺伝的なバックグラウンドにおいて収量性を向上させるため有用な遺伝子であると考えられた.

GS3遺伝子,SW5遺伝子,GS5遺伝子,GIF1遺伝子は,すぐれた形質をもつ品種からクローニングされた.それとは対照的に,TGW6遺伝子は収量特性の劣るインドの在来種からクローニングされた.このことは,現代品種の改良には作物の栽培化の過程で置き去られた,表現型に現われないすぐれた遺伝子を収集する必要があるという,古くからの仮説10) を実験的に実証するものである.遺伝子あるいは対立遺伝子の価値は遺伝的にあるいは社会情勢により変化する.TGW6遺伝子は栽培化の過程において選抜の対象にされていなかったことから,栽培化の過程ではほかの栽培化遺伝子と比べ優先度は低かったと考えられる.しかし,シンクの拡大した現代品種では,ソース能とシンクのサイズの両方において多面的に作用するTGW6遺伝子の有用性は高い.さらに,現代品種の改良においては,今後の地球温暖化の影響も考慮に入れなければならない.東南アジアにおいては登熟期の高温によりシンクとソースのアンバランスが生じ,イネの収量や玄米の品質の低下がひき起こされており,わが国では2010年だけで被害が176,200トンにも及ぶと見積もられている.TGW6遺伝子はシンクとソースに多面的に作用しこのような被害を減らすことも確認された.
略歴:1991年 農業生物資源研究所 研究員,2001年 イタリアMilan大学 客員研究員を経て,2002年より農業生物資源研究所 主任研究員.
研究テーマ:イネの炭素代謝.
抱負:研究成果を田圃へ食卓へ.
廣津 直樹(Naoki Hirotsu)
東洋大学生命科学部 准教授.
© 2013 石丸 健・廣津直樹 Licensed under CC 表示 2.1 日本
(1農業生物資源研究所植物科学研究領域 植物生産生理機能研究ユニット,2東洋大学生命科学部 生命科学科植物ゲノム科学研究室)
email:石丸 健,廣津直樹
DOI: 10.7875/first.author.2013.060
Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield.
Ken Ishimaru, Naoki Hirotsu, Yuka Madoka, Naomi Murakami, Nao Hara, Haruko Onodera, Takayuki Kashiwagi, Kazuhiro Ujiie, Bun-ichi Shimizu, Atsuko Onishi, Hisashi Miyagawa, Etsuko Katoh
Nature Genetics, 45, 707-711 (2013)
要 約
急速な人口の増加をささえるため,世界の人口の半数以上の主食であるイネの収量を飛躍的に高めることが求められている.粒重はイネの収量性を決定する主要な要因である.筆者らは,インドのイネ在来種であるカサラスからTGW6遺伝子をクローニングしその機能を明らかにすることに成功した.この遺伝子は,インドール酢酸-グルコースの加水分解活性をもつ新規なタンパク質をコードしていた.イネの品種のひとつ日本晴において,tgw6により合成されたインドール酢酸はシンクおよびソースにおいて抑制的に作用した.一方で,機能欠失型変異をもつカサラス型のTGW6はインドール酢酸を合成できず抑制は起こらなかった.そのため,シンクでは胚乳細胞の数が増加し粒長が伸長した.それにくわえて,ソース能が向上することにより粒重ならびに収量も増加した.また,遺伝子の多様性解析の結果から,TGW6遺伝子は栽培化の過程で忘れさられた“もったいない”遺伝子であることが明らかにされた.これらのことより,TGW6遺伝子は近代あるいは現代の栽培品種における収量の改良に広く利用可能であると考えられた.
はじめに
現在,栽培されているイネは,長いあいだ人の手による選抜をくり返すことによりつくられてきた.したがって,近代あるいは現代の栽培品種は多くの有用遺伝子が集積したものということができる.そのために遺伝的な多様性は失われ,ある品種から有用な遺伝子を特定したとしても,ほかの品種もすでにその遺伝子をもっている可能性が高く,遺伝子の利用範囲は限定されてしまっている.ごくかぎられた品種しかもたない有用な遺伝子をみつけることができれば,多くの品種の改良に利用することができる.栽培化の過程では,種子のサイズなどのシンクの拡大をもとに収量性は増大してきたと考えられている.実際,イネの種子サイズにかかわるいくつかの遺伝子がクローニングされたが,これらの遺伝子は栽培化の過程において選抜に用いられていた.
近代品種ではシンクへの炭水化物の供給能力(ソース能)が収量性を規定する要因となっている.しかしながら,ソース能を制御する遺伝子や分子機構についての情報はこれまでほとんど得られていない.出穂のまえに植物に蓄積された炭水化物は,出穂ののち種子に供給される.そのため,出穂前の炭水化物の蓄積量を増加させることがイネのソース能の改良にむけた主要な育種目標とされてきた.筆者らは,イネの品種のひとつ日本晴とインドのイネ在来種であるカサラスとのあいだの戻し交雑自殖系統を用いて,粒重を増加させる量的形質遺伝子座qTGW6を同定した1).カサラス型のqTGW6遺伝子座を導入した準同質遺伝子系統NIL (TGW6) の解析の結果,qTGW6遺伝子座は出穂前の炭水化物の蓄積量を増加させ,その結果,収量を約15%増加させた.そこで,イネの収量性の向上にむけた知見を得ることを目的に,TGW6遺伝子をクローニングしその機能解析を行った.
1.TGW6遺伝子のポジショナルクローニング
ホモ接合型の組換え個体を用いて高密度マッピングを行った結果,TGW6遺伝子の存在する領域を4.9 kbpに絞り込んだ.この領域には1個のエキソンをもつ遺伝子Os06g0623700のみが存在していた.日本晴型tgw6遺伝子と比較して,カサラス型TGW6遺伝子には6個のSNP(single nucleotide polymorphism,一塩基多型)と313番目の塩基の欠失が存在していた.この欠失は終止コドンを生じさせ正常なタンパク質の合成を阻害し機能欠失をひき起こすと考えられた.一方で,6個のSNPはコードされるタンパク質の特性に影響を及ぼさないと予想された.相補試験として,日本晴およびNIL (TGW6) にtgw6遺伝子のRNAiコンストラクトを導入した組換え体を作出して影響を解析した.日本晴のRNAi形質転換体の粒長や粒重は対照と比べ有意に増加した.RNAi形質転換体においてtgw6遺伝子の発現量と粒長とのあいだには負の相関関係がみられた.一方で,NIL (TGW6) のRNAi形質転換体では粒長に変化はみられなかった.さらに,日本晴型tgw6遺伝子をカサラス型のプロモーター配列に連結してNIL (TGW6) に導入したところ粒長と粒重が有意に減少した.これらの結果から,TGW6遺伝子がqTGW6遺伝子座の原因遺伝子であり,この313番目の1塩基欠失が対立遺伝子の多型を決定する機能的な変異であると結論した.
2.TGW6遺伝子の発現する部位と胚乳細胞への影響
TGW6遺伝子はシンクとソースの両方の器官において発現していた.胚乳におけるTGW6遺伝子の発現量は,受精2日後にピークに達し,受精5日後にはピーク値の約30%まで減少した.TGW6遺伝子は細胞分裂の起こっている胚乳の周辺部で強く発現していた.日本晴と比較してNIL (TGW6) では胚乳細胞の数が約24%増加していた.胚乳細胞の形成の初期段階では細胞分裂は一時的に停止し多核体が形成され,この期間の長さが細胞の数を決定する.胚乳細胞の形成の初期段階において,NIL (TGW6) では細胞分裂に関与する遺伝子の発現レベルが対照に比べ低かったことから,カサラス型TGW6遺伝子が細胞分裂への移行を遅らせ細胞の数を増加させていると考えられた.
3.日本晴型のtgw6の機能解析
Conserved Domain DatabaseのRPS-BLAST解析により日本晴型のtgw6と有意な相同性のみられたストリクトシジンシンターゼおよびジイソプロピルフルオロホスファターゼとの立体構造を比較したところ,ジイソプロピルフルオロホスファターゼのCa2+結合残基はtgw6においても保存されていた.さらに,tgw6の基質ポケットにおけるHis192とTyr224の空間的な配置が加水分解酵素の活性部位と類似していたことから,tgw6は加水分解能をもつことが予測された.さらに,立体構造モデルとの結合シミュレーションによりインドール酢酸-グルコースが基質である可能性が示された.実際に,日本晴型のtgw6はインドール酢酸-グルコースを加水分解したが,カサラス型のTGW6はその活性をもっていなかった.NIL (TGW6) の受精3日後の胚乳において,インドール酢酸の含量は日本晴のわずか16.6%であり,ほとんどのオーキシン応答性遺伝子の発現レベルは低かった.また,NIL (TGW6) の受精の直後の胚珠にインドール酢酸を処理すると,受精5日後の胚乳細胞の長さが減少した.以上の結果から,tgw6によりインドール酢酸-グルコースの加水分解からつくられたインドール酢酸が細胞分裂をひき起こすことにより,胚乳細胞の数や粒長を抑制していると考えられた.そのため,機能欠失型変異をもつカサラス型のTGW6はインドール酢酸を合成できないため抑制作用がはたらかず,その結果,粒長が増加したものと考えられた.
4.TGW6遺伝子がソース能へ及ぼす影響
NIL (TGW6) において,出穂のまえに炭水化物の蓄積をおもに担う葉鞘での炭水化物の蓄積量は日本晴よりも多かった.また,NIL (TGW6) の出穂前の葉鞘にインドール酢酸を処理すると,デンプン合成関連遺伝子の発現レベルが低下した.つまり,日本晴型tgw6はインドール酢酸の合成を介して,シンクのみならずソースにおいても抑制的に作用するものと考えられた.また,葉を切除して炭水化物の供給量を減少させたNIL (TGW6) では,粒長は変わらないにもかかわらず粒重の有意な低下が観察された.これらの結果から,カサラス型TGW6遺伝子はソースとシンクにおいて多面的に作用し,そのことが日本晴という現代品種の収量をひとつの遺伝子により改良することのできる要因であると考えられた(図1).
5.TGW6遺伝子のクローニングによりみえてきたこと
TGW6遺伝子と同様に機能欠失型変異により種子のサイズの増加することは,GW2遺伝子2) やGS3遺伝子3),SW5遺伝子4) でも報告されており,イネにおいて種子のサイズを制限するなんらかの機構の存在する可能性が示唆された.ただし,TGW6遺伝子とは対照的に,これまで報告されている種子サイズを制御する遺伝子(GW2遺伝子,GS3遺伝子,SW5遺伝子,GS5遺伝子5) やGW8遺伝子6) など)は籾のサイズの制御をつうじて間接的に種子に作用する.また,ひとつの形質の改良による収量性の改良は限定的である.たとえば,シンクのサイズが収量を規定している品種では粒のサイズの改良は有効であるが,ボトルネックがほかに移り収量を規定する7).イネの収量を飛躍的に増加させるためには,シンクサイズのみならず炭水化物の生産,転流および蓄積能を並行して改良していく必要がある.
6.TGW6遺伝子の自然変異と現代品種への活用
国立遺伝研究所の野生イネ(Oryza rufipogon)コアコレクション8) や農業生物資源研究所が作出した世界イネコアコレクション9) を用いて,遺伝子の多様性解析を行った(図2).その結果,TGW6遺伝子のORFには17のハプロタイプの存在することが確認された.カサラス型TGW6遺伝子に存在する機能的な変異をもつものは,野生イネ1系統(O. perennis W1690)とインドシナ地方原産のイネ4品種のみであった.カサラス型TGW6遺伝子のもつ機能的な変異のほか,自然変異には8個のSNPが存在していた.そのうち4個のSNPはアミノ酸置換を生じさせるが,置換されるアミノ酸残基は活性部位から遠くはなれていた.そのため,これらのSNPはタンパク質の構造ひいては特性にほとんど影響を及ぼさないものと考えられた.実際,イネの品種のひとつIR64のtgw6には,313番目の塩基の1塩基欠失は含まれないがカサラスのもつSNPのうち6個(うち,3個がアミノ酸置換をひき起こす)が含まれていた.IR64型のtgw6遺伝子を導入した準同質遺伝子系統では粒長に変化はみられなかった.この結果は,313番目の1塩基欠失がカサラス型TGW6遺伝子のもつ機能的な変異であることを強く裏づけた.これらの結果により,TGW6遺伝子は栽培化の過程において選抜の対象とされずに捨てさられた“もったいない”遺伝子であると考えられた.事実,コシヒカリやNERICAなどのさまざまな現代品種は,カサラス型TGW6遺伝子のもつ機能的な変異をもたない.カサラス型TGW6遺伝子をコシヒカリに導入した準同質遺伝子系統では籾の大きさは変化せずに粒長や粒重が増加し,単位面積あたりの収量は有意に増加した.これらのことから,TGW6遺伝子は広い遺伝的なバックグラウンドにおいて収量性を向上させるため有用な遺伝子であると考えられた.
おわりに
GS3遺伝子,SW5遺伝子,GS5遺伝子,GIF1遺伝子は,すぐれた形質をもつ品種からクローニングされた.それとは対照的に,TGW6遺伝子は収量特性の劣るインドの在来種からクローニングされた.このことは,現代品種の改良には作物の栽培化の過程で置き去られた,表現型に現われないすぐれた遺伝子を収集する必要があるという,古くからの仮説10) を実験的に実証するものである.遺伝子あるいは対立遺伝子の価値は遺伝的にあるいは社会情勢により変化する.TGW6遺伝子は栽培化の過程において選抜の対象にされていなかったことから,栽培化の過程ではほかの栽培化遺伝子と比べ優先度は低かったと考えられる.しかし,シンクの拡大した現代品種では,ソース能とシンクのサイズの両方において多面的に作用するTGW6遺伝子の有用性は高い.さらに,現代品種の改良においては,今後の地球温暖化の影響も考慮に入れなければならない.東南アジアにおいては登熟期の高温によりシンクとソースのアンバランスが生じ,イネの収量や玄米の品質の低下がひき起こされており,わが国では2010年だけで被害が176,200トンにも及ぶと見積もられている.TGW6遺伝子はシンクとソースに多面的に作用しこのような被害を減らすことも確認された.
文 献
- Ishimaru, K.: Identification of a locus increasing rice yield and physiological analysis of its function. Plant Physiol., 133, 1083-1090 (2003)[PubMed]
- Song, X. -J., Huang, W., Shi, M. et al.: A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat. Genet., 39, 623-630 (2007)[PubMed]
- Fan, C., Xing, Y., Mao, H. et al.: GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet., 112, 1164-1171 (2006)[PubMed]
- Shomura, A., Izawa, T., Ebana, K. et al.: Deletion in a gene associated with grain size increased yields during rice domestication. Nat. Genet., 40, 1023-1028 (2008)[PubMed]
- Li, Y., Fan, C., Xing, Y. et al.: Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat. Genet., 43, 1266-1269 (2011)[PubMed]
- Wang, S., Wu, K., Yuan, Q. et al.: Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet., 44, 950-954 (2012)[PubMed]
- Wang, E., Wang, J., Zhu, X. et al.: Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat. Genet., 40, 1370-1374 (2008)[PubMed]
- Kurata, N. & Yamazaki, Y.: Oryzabase. An integrated biological and genome information database for rice. Plant Physiol., 140, 12-17 (2006)[PubMed]
- Kojima, Y., Ebana, K., Fukuoka, S. et al.: Development of an RFLP-based rice diversity research set of germplasm. Breed. Sci., 55, 431-440 (2005)
- Tanksley, S. D. & McCouch, S. R.: Seed banks and molecular maps: unlocking geneticpotential from the wild. Science, 277, 1063-1066 (1997)[PubMed]
著者プロフィール
略歴:1991年 農業生物資源研究所 研究員,2001年 イタリアMilan大学 客員研究員を経て,2002年より農業生物資源研究所 主任研究員.
研究テーマ:イネの炭素代謝.
抱負:研究成果を田圃へ食卓へ.
廣津 直樹(Naoki Hirotsu)
東洋大学生命科学部 准教授.
© 2013 石丸 健・廣津直樹 Licensed under CC 表示 2.1 日本
