ストア作動性Ca2+流入は制御性T細胞などアゴニスト選択性T細胞の選択ののちの成熟を制御する
大洞 将嗣
(東京医科歯科大学 歯と骨のグローバルCOEプログラム)
email:大洞将嗣
DOI: 10.7875/first.author.2013.034
Agonist-selected T cell development requires strong T cell receptor signaling and store-operated calcium entry.
Masatsugu Oh-hora, Noriko Komatsu, Mojgan Pishyareh, Stefan Feske, Shohei Hori, Masaru Taniguchi, Anjana Rao, Hiroshi Takayanagi
Immunity, 38, 881-895 (2013)
胸腺におけるT細胞の分化では,T細胞受容体とMHC-自己抗原複合体との結合の強度によりT細胞の運命が決定する.しかし,T細胞受容体の刺激の強度によりT細胞の運命決定を制御されるシグナル伝達経路については不明な点が多く残っている.この研究において,筆者らは,T細胞受容体の刺激により惹起されるストア作動性の細胞外Ca2+の流入の欠損により,通常のT細胞の分化や選択はほぼ正常であった一方,制御性T細胞などアゴニスト選択性T細胞の分化が選択的に阻害されていることを見い出した.この分化の阻害はアゴニスト選択ののち前駆細胞から成熟した細胞へと増殖し成熟していく段階で生じていることが判明した.さらに,転写因子NFATのCa2+依存性の活性化を部分的に阻害した場合,やはりアゴニスト選択性T細胞の分化は選択的に阻害されることがわかった.以上,T細胞受容体の刺激により惹起されるストア作動性の細胞外Ca2+の流入と,それにともなうNFATの高い活性化がアゴニスト選択性T細胞の選択ののちの成熟に必須であることが明らかになった.
免疫系は“自己”と“非自己”を識別し,自己である自身の組織は攻撃せず,非自己である細菌やウイルスなど病原体による感染症から身を守る重要なシステムである.免疫応答が過剰になると正常な細胞や組織を攻撃してしまい自己免疫疾患やアレルギーなどがひき起こされる.そのため,免疫応答においては活性化と抑制のバランスを保つことがとても重要になる.
免疫応答の主役を担うリンパ球であるT細胞は胸腺において発生する.これまでの研究から,胸腺においてCD4陽性CD8陽性細胞がT細胞受容体とMHC-自己抗原複合体との結合により生じるT細胞受容体シグナルの強度により選択をうけ成熟T細胞となることが明らかにされている.自己のMHC-自己抗原複合体を認識できずに非常に弱い反応を示すあるいは反応しない場合,“無の選択”によりCD4陽性CD8陽性細胞はアポトーシスに導かれ除去される.一方,自己のMHC-自己抗原複合体に反応した場合には非常に強いT細胞受容体の刺激が惹起され,“負の選択”により,やはりCD4陽性CD8陽性細胞はアポトーシスに導かれ除去される.そして,自己のMHC-自己抗原複合体と“適度”に反応した場合には適度なT細胞受容体の刺激によりCD4陽性CD8陽性細胞は“正の選択”をうけ,機能的なT細胞として成熟し末梢に供給される.
しかし最近になり,自己反応性のT細胞受容体をもつ一部のT細胞はアポトーシスにより除去されず成熟することがわかってきた1,2).このような自己反応性の成熟T細胞は,制御性T細胞,インバリアントナチュラルキラーT細胞,一部の腸管上皮細胞間リンパ球などであり,これらの細胞は抑制的な機能をもち,免疫応答の過剰な活性化や活性化した免疫細胞による自身の組織に対する攻撃を防ぐ重要な役割を担っていると考えられている.これらのT細胞は“アゴニスト選択”により選択され,このアゴニスト選択には正の選択と負の選択とのあいだの刺激が必要だと予測されているが,どのようなシグナル伝達系が必要であるかは明らかになっていない.
T細胞受容体から刺激をうけると,T細胞では細胞内のCa2+濃度が2段階で増加することが明らかにされている.第1段階として,小胞体に蓄えられていたCa2+が細胞質へと放出される.第2段階として,小胞体のCa2+がなくなったことをCa2+センサータンパク質であるSTIM1が感知すると,細胞膜にあるストア作動性Ca2+チャネルが活性化され細胞外から大量かつ長時間のCa2+の流入(ストア作動性Ca2+流入)が誘導される.その結果,Ca2+依存性の転写因子であるNFATなどが長時間にわたり活性化し,T細胞の増殖やサイトカインの産生が誘導される3,4).
T細胞受容体の刺激の強さとCa2+の流入の大きさあるいは長さとはよく相関することが知られている.負の選択では大きなCa2+の流入が誘導される一方,正の選択では比較的小さく持続的なCa2+の流入が誘導される5-7).ストア作動性Ca2+流入はこれら両方のCa2+の流入パターンに対応していることから,長いあいだT細胞の分化に必須であると予測されてきたが,その証明はされていなかった.そこで筆者らは,T細胞の分化におけるストア作動性Ca2+流入の役割を明らかにすることをめざした.
筆者らは以前に,STIM1およびそのホモログであるSTIM2のコンディショナルノックアウトマウスにおいて,T細胞の分化は正常であることを報告した8).しかし,ここで用いたコンディショナルノックアウトマウスはT細胞の分化の後期において欠損を起こすため,T細胞の分化についてその全体を評価することはできなかった.そこでこの研究では,STIM1とSTIM2のダブルノックアウトマウスに由来する造血幹細胞を含む胎仔の肝細胞を,T細胞を完全に欠損するRag2ノックアウトマウスに移植し,ストア作動性Ca2+流入のT細胞の分化における役割を解析した.CD4とCD8をマーカーとして用いたときCD4陽性CD8陽性細胞におけるストア作動性Ca2+流入は完全に欠損していたにもかかわらず,予想に反し,CD4陰性CD8陰性細胞,CD4陽性CD8陽性細胞,CD4陽性CD8陰性細胞,CD4陰性CD8陽性細胞の比率および細胞数は,野生型マウスとSTIMダブルノックアウトマウスとで同じであった.つぎに,T細胞に特異的なSTIMダブルノックアウトマウスとHY抗原に特異的なT細胞受容体トランスジェニックマウスを交配したマウスを用いて,正の選択および負の選択の解析を行った.その結果,HY抗原を発現する雄マウスにおける負の選択にいくらかの障害が認められた.さらに,黄色ブドウ球菌に由来する毒素を投与し特定の未熟胸腺細胞に誘導される負の選択を調べた結果,やはり一定の程度で障害が認められた.一方,HY抗原を発現しない雌マウスにおける正の選択は正常であり,T細胞受容体のα鎖およびβ鎖の可変領域のレパートリーにも大きな変化はなかった.以上の結果から,STIMにより制御されるストア作動性Ca2+流入はT細胞の正の選択には必須ではないことが明らかになった.
ストア作動性Ca2+流入の欠損により負の選択に部分的な障害があったという結果は,自己反応性のT細胞受容体と自己抗原による強いT細胞受容体シグナルにおいてストア作動性Ca2+流入が重要であることを示唆した.そこで,ストア作動性Ca2+流入が自己反応性をもつ成熟したT細胞“アゴニスト選択性T細胞”の分化を制御しているのではないかと考えた.末梢のリンパ組織や臓器におけるアゴニスト選択性T細胞を調べた結果,Foxp3陽性の制御性T細胞,スフィンゴ糖脂質αガラクトシルセラミドを認識するインバリアントナチュラルキラーT細胞,腸管上皮細胞のあいだに局在するT細胞受容体αβ陽性CD8αα陽性リンパ球は,末梢にほとんど存在していなかった.これらの細胞の欠損が細胞に自律的なものであるかどうかを確認するため,野生型マウスおよびSTIMダブルノックアウトマウスに由来する骨髄細胞を用いて競合的な混合骨髄キメラマウスを作製した.その結果,野生型の細胞が存在していてもSTIMを欠損した骨髄細胞からこの3種類のアゴニスト選択性T細胞への分化は認められなかった.さらに,Bcl-2の導入やブロモデオキシウリジンを用いた生存細胞の追跡などを行ったが,STIMの欠損により細胞の生存に大きな影響は認められなかった.したがって,ストア作動性Ca2+流入はアゴニスト選択性T細胞の分化を優先的に制御しているものと考えられた.
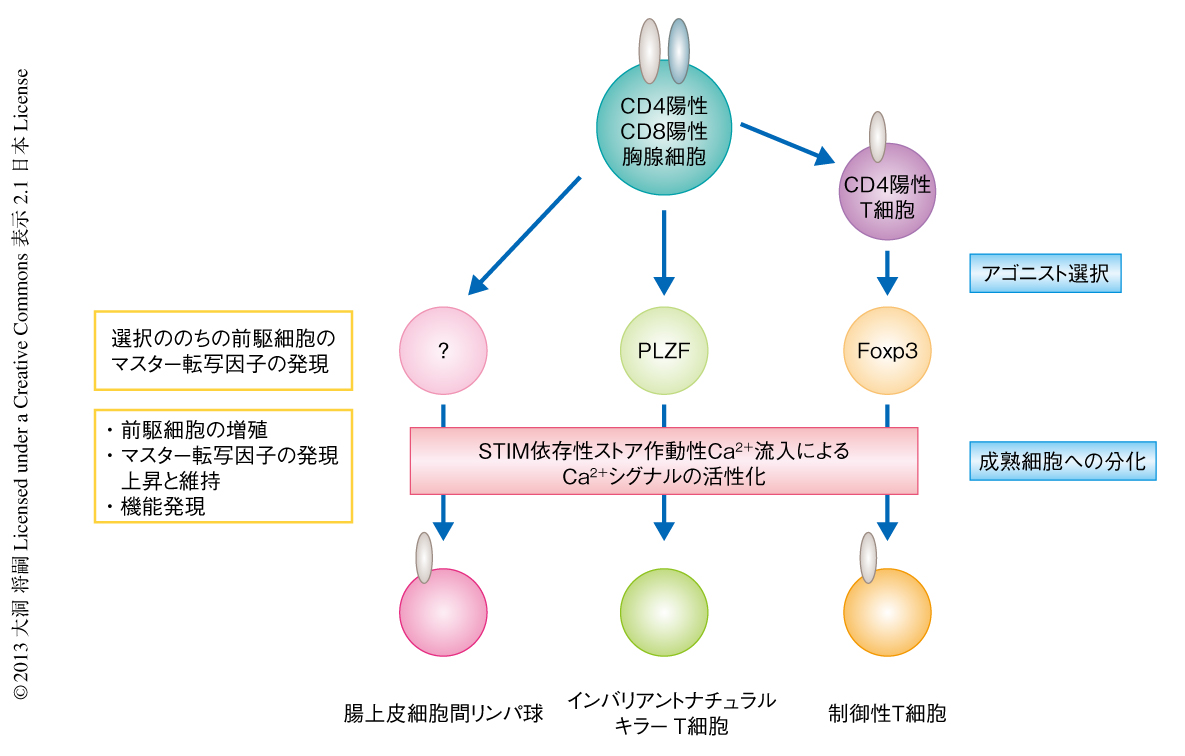
これらのアゴニスト選択性T細胞は胸腺で分化したのち末梢に分布する.そこで,ストア作動性Ca2+流入がアゴニスト選択性T細胞の分化のどの段階を制御しているのか明らかにするため,胸腺におけるアゴニスト選択性T細胞の分化を検証した.その結果,ストア作動性Ca2+流入の欠損により,もっとも未熟なインバリアントナチュラルキラーT細胞の前駆細胞は正常であった一方,分化の進んだインバリアントナチュラルキラーT細胞の前駆細胞はほぼ消失していた.腸管上皮細胞間リンパ球の前駆細胞が含まれていると考えられた細胞の集団はストア作動性Ca2+流入の欠損により減少していた.同様に,Foxp3陽性の制御性T細胞の場合も,未熟な前駆細胞の存在する集団は正常であったが,成熟した制御性T細胞のみが著減していた.これらの欠損に共通にみられる分子機構は,インターロイキン15受容体β鎖であるCD122,あるいは,インターロイキン2受容体α鎖であるCD25の発現の上昇が誘導されず,前駆細胞の増殖が阻害されていた点であった.さらに,それぞれの前駆細胞において,PLZFやFoxp3などマスター転写因子の発現は誘導されていたものの,CD122やCD25と同様に,そののちの発現の上昇が阻害されていた.以上の結果から,ストア作動性Ca2+流入はアゴニスト選択の直後の,前駆細胞から成熟した細胞にいたる成熟の過程を制御していることが明らかになった.
インターロイキン2やインターロイキン15はアゴニスト選択性T細胞の分化を促進することが知られている.そこで,STIMダブルノックアウトマウスやそれに由来する細胞にインターロイキン2あるいはインターロイキン15を投与した場合に分化が回復するかどうかを検証した.この実験では,持続的なサイトカインの刺激を生体において維持するため,インターロイキン2と抗インターロイキン2抗体の複合体,あるいは,インターロイキン15とインターロイキン15受容体α鎖の複合体を投与する方法を用いた.インターロイキン2をSTIMダブルノックアウトマウスに投与した場合,成熟した制御性T細胞の比率,数,増殖能は対照となるマウスと同じレベルにまで回復したが,Foxp3の発現の上昇は誘導されなかった.この回復した制御性T細胞では抑制機能は回復しておらず,その原因は抑制機能をもつ細胞表面タンパク質であるCTLA-4やTIGITの発現レベルが低下したままであるためだと考えられた.すなわち,ストア作動性Ca2+流入は,制御性T細胞の前駆細胞の増殖や成熟だけでなく,抑制機能の発現も制御していることが明らかになった.一方,インターロイキン15を投与した場合はアゴニスト選択性T細胞の比率にある程度の上昇が認められた程度であったが,インターロイキン15の存在のもと腸管上皮細胞間リンパ球の前駆細胞を培養したところ成熟した細胞へと分化した.この結果は,ストア作動性Ca2+流入は腸管上皮細胞間リンパ球の前駆細胞の胸腺からインターロイキン15の豊富な腸リンパ組織への移動を制御している可能性を示唆した.
ここまでの結果は,アゴニスト選択性T細胞の成熟には持続的かつ大きなCa2+の流入およびCa2+シグナルが必要であることを強く示唆した.そこで,それぞれの前駆細胞を抗CD3抗体により刺激しCa2+の流入を測定したところ,予想どおり,アゴニスト選択性T細胞の前駆細胞にはCD4陽性CD8陽性細胞に比べ大きなCa2+の流入が惹起された.Ca2+の流入に依存して活性化される転写因子NFATは,T細胞において3種類が発現している.そのうちNFAT4は,これを単独でノックアウトしたマウスにおいて通常のT細胞の分化に影響の生じることが報告されていたので,NFAT1とNFAT2のダブルノックアウトマウスを用いてアゴニスト選択性T細胞の分化を調べた.その結果,このダブルノックアウトマウスでは通常のT細胞の分化は正常であった一方,胸腺や末梢において制御性T細胞などの前駆細胞や成熟した細胞の比率や数が減少しており,すべてのアゴニスト選択性T細胞の分化が選択的に障害されていた.以上の結果から,大きなCa2+の流入によるNFATの完全な活性化がアゴニスト選択性T細胞の成熟に必須であることが明らかになった.
これまでは,Ca2+依存性の脱リン酸化酵素であるカルシニューリンがT細胞の正の選択を制御しているという報告から9),その上流であるストア作動性Ca2+流入も同様に正の選択に重要であると予想されていた.しかし,今回の研究から,STIMにより惹起されるストア作動性Ca2+流入とそれによるCa2+シグナルは,T細胞の正の選択ではなく,自己反応性のT細胞受容体をもつアゴニスト選択性T細胞の最終分化および成熟に必須であることが明らかになった(図1).今後,正の選択を制御するCa2+の流入機構あるいはCa2+シグナルがマスター転写因子の発現したあとの分化を促進する分子機構を明らかにすることにより,それぞれのT細胞の分化に必要なT細胞受容体シグナルあるいはCa2+シグナルの強度,それぞれのT細胞の最終分化の機序やその性質の維持機構の解明へとつながることが期待される.
略歴:2000年 大阪大学大学院医学系研究科博士課程 修了,2002年 関西医科大学肝臓研究所 助手,2004年 米国Harvard大学 研究員,2007年 同 インストラクター,2008年 東京医科歯科大学 特任准教授を経て,2013年より九州大学生体防御医学研究所 准教授.
研究テーマ:免疫受容体からのシグナル伝達経路.
関心事:細胞の運命決定,その性質の維持とシグナル伝達系との関係.
© 2013 大洞 将嗣 Licensed under CC 表示 2.1 日本
(東京医科歯科大学 歯と骨のグローバルCOEプログラム)
email:大洞将嗣
DOI: 10.7875/first.author.2013.034
Agonist-selected T cell development requires strong T cell receptor signaling and store-operated calcium entry.
Masatsugu Oh-hora, Noriko Komatsu, Mojgan Pishyareh, Stefan Feske, Shohei Hori, Masaru Taniguchi, Anjana Rao, Hiroshi Takayanagi
Immunity, 38, 881-895 (2013)
要 約
胸腺におけるT細胞の分化では,T細胞受容体とMHC-自己抗原複合体との結合の強度によりT細胞の運命が決定する.しかし,T細胞受容体の刺激の強度によりT細胞の運命決定を制御されるシグナル伝達経路については不明な点が多く残っている.この研究において,筆者らは,T細胞受容体の刺激により惹起されるストア作動性の細胞外Ca2+の流入の欠損により,通常のT細胞の分化や選択はほぼ正常であった一方,制御性T細胞などアゴニスト選択性T細胞の分化が選択的に阻害されていることを見い出した.この分化の阻害はアゴニスト選択ののち前駆細胞から成熟した細胞へと増殖し成熟していく段階で生じていることが判明した.さらに,転写因子NFATのCa2+依存性の活性化を部分的に阻害した場合,やはりアゴニスト選択性T細胞の分化は選択的に阻害されることがわかった.以上,T細胞受容体の刺激により惹起されるストア作動性の細胞外Ca2+の流入と,それにともなうNFATの高い活性化がアゴニスト選択性T細胞の選択ののちの成熟に必須であることが明らかになった.
はじめに
免疫系は“自己”と“非自己”を識別し,自己である自身の組織は攻撃せず,非自己である細菌やウイルスなど病原体による感染症から身を守る重要なシステムである.免疫応答が過剰になると正常な細胞や組織を攻撃してしまい自己免疫疾患やアレルギーなどがひき起こされる.そのため,免疫応答においては活性化と抑制のバランスを保つことがとても重要になる.
免疫応答の主役を担うリンパ球であるT細胞は胸腺において発生する.これまでの研究から,胸腺においてCD4陽性CD8陽性細胞がT細胞受容体とMHC-自己抗原複合体との結合により生じるT細胞受容体シグナルの強度により選択をうけ成熟T細胞となることが明らかにされている.自己のMHC-自己抗原複合体を認識できずに非常に弱い反応を示すあるいは反応しない場合,“無の選択”によりCD4陽性CD8陽性細胞はアポトーシスに導かれ除去される.一方,自己のMHC-自己抗原複合体に反応した場合には非常に強いT細胞受容体の刺激が惹起され,“負の選択”により,やはりCD4陽性CD8陽性細胞はアポトーシスに導かれ除去される.そして,自己のMHC-自己抗原複合体と“適度”に反応した場合には適度なT細胞受容体の刺激によりCD4陽性CD8陽性細胞は“正の選択”をうけ,機能的なT細胞として成熟し末梢に供給される.
しかし最近になり,自己反応性のT細胞受容体をもつ一部のT細胞はアポトーシスにより除去されず成熟することがわかってきた1,2).このような自己反応性の成熟T細胞は,制御性T細胞,インバリアントナチュラルキラーT細胞,一部の腸管上皮細胞間リンパ球などであり,これらの細胞は抑制的な機能をもち,免疫応答の過剰な活性化や活性化した免疫細胞による自身の組織に対する攻撃を防ぐ重要な役割を担っていると考えられている.これらのT細胞は“アゴニスト選択”により選択され,このアゴニスト選択には正の選択と負の選択とのあいだの刺激が必要だと予測されているが,どのようなシグナル伝達系が必要であるかは明らかになっていない.
T細胞受容体から刺激をうけると,T細胞では細胞内のCa2+濃度が2段階で増加することが明らかにされている.第1段階として,小胞体に蓄えられていたCa2+が細胞質へと放出される.第2段階として,小胞体のCa2+がなくなったことをCa2+センサータンパク質であるSTIM1が感知すると,細胞膜にあるストア作動性Ca2+チャネルが活性化され細胞外から大量かつ長時間のCa2+の流入(ストア作動性Ca2+流入)が誘導される.その結果,Ca2+依存性の転写因子であるNFATなどが長時間にわたり活性化し,T細胞の増殖やサイトカインの産生が誘導される3,4).
T細胞受容体の刺激の強さとCa2+の流入の大きさあるいは長さとはよく相関することが知られている.負の選択では大きなCa2+の流入が誘導される一方,正の選択では比較的小さく持続的なCa2+の流入が誘導される5-7).ストア作動性Ca2+流入はこれら両方のCa2+の流入パターンに対応していることから,長いあいだT細胞の分化に必須であると予測されてきたが,その証明はされていなかった.そこで筆者らは,T細胞の分化におけるストア作動性Ca2+流入の役割を明らかにすることをめざした.
1.ストア作動性Ca2+流入は正の選択に必須ではない
筆者らは以前に,STIM1およびそのホモログであるSTIM2のコンディショナルノックアウトマウスにおいて,T細胞の分化は正常であることを報告した8).しかし,ここで用いたコンディショナルノックアウトマウスはT細胞の分化の後期において欠損を起こすため,T細胞の分化についてその全体を評価することはできなかった.そこでこの研究では,STIM1とSTIM2のダブルノックアウトマウスに由来する造血幹細胞を含む胎仔の肝細胞を,T細胞を完全に欠損するRag2ノックアウトマウスに移植し,ストア作動性Ca2+流入のT細胞の分化における役割を解析した.CD4とCD8をマーカーとして用いたときCD4陽性CD8陽性細胞におけるストア作動性Ca2+流入は完全に欠損していたにもかかわらず,予想に反し,CD4陰性CD8陰性細胞,CD4陽性CD8陽性細胞,CD4陽性CD8陰性細胞,CD4陰性CD8陽性細胞の比率および細胞数は,野生型マウスとSTIMダブルノックアウトマウスとで同じであった.つぎに,T細胞に特異的なSTIMダブルノックアウトマウスとHY抗原に特異的なT細胞受容体トランスジェニックマウスを交配したマウスを用いて,正の選択および負の選択の解析を行った.その結果,HY抗原を発現する雄マウスにおける負の選択にいくらかの障害が認められた.さらに,黄色ブドウ球菌に由来する毒素を投与し特定の未熟胸腺細胞に誘導される負の選択を調べた結果,やはり一定の程度で障害が認められた.一方,HY抗原を発現しない雌マウスにおける正の選択は正常であり,T細胞受容体のα鎖およびβ鎖の可変領域のレパートリーにも大きな変化はなかった.以上の結果から,STIMにより制御されるストア作動性Ca2+流入はT細胞の正の選択には必須ではないことが明らかになった.
2.ストア作動性Ca2+流入の欠損はアゴニスト選択される抑制性T細胞の分化を特異的に阻害する
ストア作動性Ca2+流入の欠損により負の選択に部分的な障害があったという結果は,自己反応性のT細胞受容体と自己抗原による強いT細胞受容体シグナルにおいてストア作動性Ca2+流入が重要であることを示唆した.そこで,ストア作動性Ca2+流入が自己反応性をもつ成熟したT細胞“アゴニスト選択性T細胞”の分化を制御しているのではないかと考えた.末梢のリンパ組織や臓器におけるアゴニスト選択性T細胞を調べた結果,Foxp3陽性の制御性T細胞,スフィンゴ糖脂質αガラクトシルセラミドを認識するインバリアントナチュラルキラーT細胞,腸管上皮細胞のあいだに局在するT細胞受容体αβ陽性CD8αα陽性リンパ球は,末梢にほとんど存在していなかった.これらの細胞の欠損が細胞に自律的なものであるかどうかを確認するため,野生型マウスおよびSTIMダブルノックアウトマウスに由来する骨髄細胞を用いて競合的な混合骨髄キメラマウスを作製した.その結果,野生型の細胞が存在していてもSTIMを欠損した骨髄細胞からこの3種類のアゴニスト選択性T細胞への分化は認められなかった.さらに,Bcl-2の導入やブロモデオキシウリジンを用いた生存細胞の追跡などを行ったが,STIMの欠損により細胞の生存に大きな影響は認められなかった.したがって,ストア作動性Ca2+流入はアゴニスト選択性T細胞の分化を優先的に制御しているものと考えられた.
3.ストア作動性Ca2+流入はアゴニスト選択性T細胞の胸腺におけるアゴニスト選択ののちの成熟に必要である
これらのアゴニスト選択性T細胞は胸腺で分化したのち末梢に分布する.そこで,ストア作動性Ca2+流入がアゴニスト選択性T細胞の分化のどの段階を制御しているのか明らかにするため,胸腺におけるアゴニスト選択性T細胞の分化を検証した.その結果,ストア作動性Ca2+流入の欠損により,もっとも未熟なインバリアントナチュラルキラーT細胞の前駆細胞は正常であった一方,分化の進んだインバリアントナチュラルキラーT細胞の前駆細胞はほぼ消失していた.腸管上皮細胞間リンパ球の前駆細胞が含まれていると考えられた細胞の集団はストア作動性Ca2+流入の欠損により減少していた.同様に,Foxp3陽性の制御性T細胞の場合も,未熟な前駆細胞の存在する集団は正常であったが,成熟した制御性T細胞のみが著減していた.これらの欠損に共通にみられる分子機構は,インターロイキン15受容体β鎖であるCD122,あるいは,インターロイキン2受容体α鎖であるCD25の発現の上昇が誘導されず,前駆細胞の増殖が阻害されていた点であった.さらに,それぞれの前駆細胞において,PLZFやFoxp3などマスター転写因子の発現は誘導されていたものの,CD122やCD25と同様に,そののちの発現の上昇が阻害されていた.以上の結果から,ストア作動性Ca2+流入はアゴニスト選択の直後の,前駆細胞から成熟した細胞にいたる成熟の過程を制御していることが明らかになった.
4.ストア作動性Ca2+流入は制御性T細胞の抑制機能に必須である
インターロイキン2やインターロイキン15はアゴニスト選択性T細胞の分化を促進することが知られている.そこで,STIMダブルノックアウトマウスやそれに由来する細胞にインターロイキン2あるいはインターロイキン15を投与した場合に分化が回復するかどうかを検証した.この実験では,持続的なサイトカインの刺激を生体において維持するため,インターロイキン2と抗インターロイキン2抗体の複合体,あるいは,インターロイキン15とインターロイキン15受容体α鎖の複合体を投与する方法を用いた.インターロイキン2をSTIMダブルノックアウトマウスに投与した場合,成熟した制御性T細胞の比率,数,増殖能は対照となるマウスと同じレベルにまで回復したが,Foxp3の発現の上昇は誘導されなかった.この回復した制御性T細胞では抑制機能は回復しておらず,その原因は抑制機能をもつ細胞表面タンパク質であるCTLA-4やTIGITの発現レベルが低下したままであるためだと考えられた.すなわち,ストア作動性Ca2+流入は,制御性T細胞の前駆細胞の増殖や成熟だけでなく,抑制機能の発現も制御していることが明らかになった.一方,インターロイキン15を投与した場合はアゴニスト選択性T細胞の比率にある程度の上昇が認められた程度であったが,インターロイキン15の存在のもと腸管上皮細胞間リンパ球の前駆細胞を培養したところ成熟した細胞へと分化した.この結果は,ストア作動性Ca2+流入は腸管上皮細胞間リンパ球の前駆細胞の胸腺からインターロイキン15の豊富な腸リンパ組織への移動を制御している可能性を示唆した.
5.アゴニスト選択性T細胞の分化には完全なNFATシグナルが必要である
ここまでの結果は,アゴニスト選択性T細胞の成熟には持続的かつ大きなCa2+の流入およびCa2+シグナルが必要であることを強く示唆した.そこで,それぞれの前駆細胞を抗CD3抗体により刺激しCa2+の流入を測定したところ,予想どおり,アゴニスト選択性T細胞の前駆細胞にはCD4陽性CD8陽性細胞に比べ大きなCa2+の流入が惹起された.Ca2+の流入に依存して活性化される転写因子NFATは,T細胞において3種類が発現している.そのうちNFAT4は,これを単独でノックアウトしたマウスにおいて通常のT細胞の分化に影響の生じることが報告されていたので,NFAT1とNFAT2のダブルノックアウトマウスを用いてアゴニスト選択性T細胞の分化を調べた.その結果,このダブルノックアウトマウスでは通常のT細胞の分化は正常であった一方,胸腺や末梢において制御性T細胞などの前駆細胞や成熟した細胞の比率や数が減少しており,すべてのアゴニスト選択性T細胞の分化が選択的に障害されていた.以上の結果から,大きなCa2+の流入によるNFATの完全な活性化がアゴニスト選択性T細胞の成熟に必須であることが明らかになった.
おわりに
これまでは,Ca2+依存性の脱リン酸化酵素であるカルシニューリンがT細胞の正の選択を制御しているという報告から9),その上流であるストア作動性Ca2+流入も同様に正の選択に重要であると予想されていた.しかし,今回の研究から,STIMにより惹起されるストア作動性Ca2+流入とそれによるCa2+シグナルは,T細胞の正の選択ではなく,自己反応性のT細胞受容体をもつアゴニスト選択性T細胞の最終分化および成熟に必須であることが明らかになった(図1).今後,正の選択を制御するCa2+の流入機構あるいはCa2+シグナルがマスター転写因子の発現したあとの分化を促進する分子機構を明らかにすることにより,それぞれのT細胞の分化に必要なT細胞受容体シグナルあるいはCa2+シグナルの強度,それぞれのT細胞の最終分化の機序やその性質の維持機構の解明へとつながることが期待される.
文 献
- Baldwin, T. A., Hogquist, K. A. & Jameson, S. C.: The fourth way? Harnessing aggressive tendencies in the thymus. J. Immunol., 173, 6515-6520 (2004)[PubMed]
- Stritesky, G. L., Jameson, S. C. & Hogquist, K. A.: Selection of self-reactive T cells in the thymus. Annu. Rev. Immunol., 30, 95-114 (2012)[PubMed]
- Feske, S.: Calcium signaling in lymphocyte activation and disease. Nat. Rev. Immunol., 7, 690-702 (2007)[PubMed]
- Hogan, P. G., Lewis, R. S., Rao, A.: Molecular basis of calcium signaling in lymphocytes: STIM and ORAI. Annu. Rev. Immunol., 28, 491-533 (2010)[PubMed]
- Daniels, M. A., Teixeiro, E., Gill, J. et al.: Thymic selection threshold defined by compartmentalization of Ras/MAPK signalling. Nature, 444, 724-729 (2006)[PubMed]
- Nakayama, T., Ueda, Y., Yamada, H. et al.: In vivo calcium elevations in thymocytes with T cell receptors that are specific for self ligands. Science, 257, 96-99 (1992)[PubMed]
- Bhakta, N. R., Oh, D. Y. & Lewis, R. S.: Calcium oscillations regulate thymocyte motility during positive selection in the three-dimensional thymic environment. Nat. Immunol., 6, 143-151 (2005)[PubMed]
- Oh-hora, M., Yamashita, M., Hogan, P. G. et al.: Dual functions for the endoplasmic reticulum calcium sensors STIM1 and STIM2 in T cell activation and tolerance. Nat. Immunol., 9, 432-443 (2008)[PubMed]
- Neilson, J. R., Winslow, M. M., Hur, E. M. et al.: Calcineurin B1 is essential for positive but not negative selection during thymocyte development. Immunity, 20, 255-266 (2004)[PubMed]
著者プロフィール
略歴:2000年 大阪大学大学院医学系研究科博士課程 修了,2002年 関西医科大学肝臓研究所 助手,2004年 米国Harvard大学 研究員,2007年 同 インストラクター,2008年 東京医科歯科大学 特任准教授を経て,2013年より九州大学生体防御医学研究所 准教授.
研究テーマ:免疫受容体からのシグナル伝達経路.
関心事:細胞の運命決定,その性質の維持とシグナル伝達系との関係.
© 2013 大洞 将嗣 Licensed under CC 表示 2.1 日本
