樹状細胞において転写因子STAT5は1型ヘルパーT細胞応答ではなく2型ヘルパーT細胞応答に重要である
北島雅之・Steven F. Ziegler
(米国Benaroya Research Institute at Virginia Mason,Immunology Program)
email:北島雅之
DOI: 10.7875/first.author.2013.027
The transcription factor STAT5 is critical in dendritic cells for the development of TH2 but not TH1 responses.
Bryan D. Bell, Masayuki Kitajima, Ryan P. Larson, Thomas A. Stoklasek, Kristen Dang, Kazuhito Sakamoto, Kay-Uwe Wagner, Boris Reizis, Lothar Hennighausen, Steven F. Ziegler
Nature Immunology, 14, 364-371 (2013)
樹状細胞は自然免疫と獲得免疫とをつなぎ免疫恒常性の維持に重要な役割をはたしているが,その分子機構にはいまだ不明な点が多く残されている.今回,筆者らは,樹状細胞において特異的に転写因子STAT5を欠損したマウスを用いた解析から,樹状細胞においてSTAT5は1型ヘルパーT細胞を必要とする炎症反応には必要なく,2型ヘルパーT細胞を必要とする皮膚や肺組織におけるアレルギー性の炎症反応に重要であることを見い出した.さらに,アレルギー性炎症部位の細胞により産生されSTAT5を活性化するサイトカインとして,TSLPが大きな役割をはたしていることを見い出した.樹状細胞におけるTSLPによるSTAT5の活性化は,1型ヘルパーT細胞の分化ではなく2型ヘルパーT細胞の分化を誘導すること,TSLP受容体を欠損したマウスは,樹状細胞に特異的にSTAT5を欠損したマウスと同様の表現型を示すことも実験的に証明した.これらの結果は,TSLPを介した樹状細胞におけるSTAT5の活性化という方向性が,表皮バリアの細胞におけるアレルギー性炎症に強く関与することを示唆する知見であると考えられた.
樹状細胞は外来性および内因性の抗原をT細胞に提示することにより,その抗原に特異的なT細胞の機能を制御して免疫恒常性の一端を担っている.樹状細胞に異常が生じると免疫恒常性は破たんをきたし,感染症,アレルギー性疾患,自己免疫疾患などの発症および増悪につながると考えられる.
転写因子は核において遺伝子の発現制御の役割を直接に担い,細胞の独自性を発揮するのに重要である.造血幹細胞から樹状細胞への分化の過程には,サイトカインFLT3Lを介した転写因子STAT3の機能がとくに重要であることが報告されている1).一方で,同じファミリーに属するSTAT5はコンベンショナルな樹状細胞への分化を亢進する一方で,形質細胞様の樹状細胞への分化を抑制することが報告されているが,生体における役割はいまだ完全には明らかでない2).樹状細胞への分化ののちも,STAT5はTSLP(thymic stromal lymphpoietin,胸腺間質性リンパ球新生因子),インターロイキン7,GM-CSFなどのサイトカインにより活性化されて細胞機能は制御される.とくに,TSLPは樹状細胞だけでなく,ランゲルハンス細胞,B細胞,T細胞などの免疫細胞に作用し,アレルギー性炎症の反応開始に重要であることが報告されている3).TSLPは細胞膜に存在するTSLP受容体とインターロイキン7受容体α鎖により認識され,その結果,STAT5だけでなくさまざまなシグナル伝達タンパク質が活性化される4,5).そのなかで,樹状細胞におけるSTAT5と疾患との直接的なかかわりは明らかになっていない.そこで筆者らは,樹状細胞におけるSTAT5の役割の解明がさまざまな疾患への治療につながることを期待して研究を開始した.
樹状細胞におけるSTAT5の役割を明らかにするため,樹状細胞において特異的にSTAT5を欠損させたコンディショナルノックアウトマウスを用いた.このノックアウトマウスの脾臓組織において,樹状細胞のサブセットおよびほかの免疫細胞は,野生型マウスと比較し正常に存在していた.また,皮膚においても,表皮,真皮,皮膚所属リンパ節における樹状細胞のサブセットの割合は正常であった.さらに,経時的に観察しても自己免疫疾患様の症状を示さなかったことから,この樹状細胞に特異的なSTAT5ノックアウトマウスでは樹状細胞への分化は正常であることが示唆された.
マウスの接触性過敏症はヒトにおける接触性皮膚炎のモデルである.抗原としてハプテンであるフルオレセインイソチオシアネートとフタル酸ジブチルを用いると,2型ヘルパーT細胞応答が優位な炎症反応の誘導されることが知られている.2型ヘルパーT細胞応答が優位な接触性過敏症モデルを誘導した樹状細胞に特異的なSTAT5ノックアウトマウスにおいて,野生型マウスと比較して皮膚の肥厚および炎症性細胞の浸潤が減少していた.さらに,2型ヘルパーT細胞が放出するサイトカインであるインターロイキン4の減少,および,CD4陽性T細胞の増殖の低下も認められた.これらの結果は,STAT5を欠損した樹状細胞では2型ヘルパーT細胞への分化が抑制されていることを示唆した.また,皮膚において抗原であるフルオレセインイソチオシアネートを取り込み所属リンパ節へと移動した樹状細胞の数は,樹状細胞に特異的なSTAT5ノックアウトマウスにおいて有意な減少を示し,樹状細胞においてT細胞刺激タンパク質であるCD86の発現の低下も認められたことから,樹状細胞の活性化が抑制されていることが示唆された.つぎに,ハプテンであるジニトロフルオロベンゼンを用いて,1型ヘルパーT細胞応答が優位な接触性過敏症モデルを誘導した.興味深いことに,このモデルでは樹状細胞に特異的なSTAT5ノックアウトマウスにおいて,野生型マウスと比較して皮膚の肥厚,1型ヘルパーT細胞が放出するサイトカインであるインターフェロンγ,および,CD4陽性T細胞とCD8陽性T細胞の増殖について有意な違いが認められなかった.
マウスの接触性過敏症モデルでは,樹状細胞の一種である表皮のランゲルハンス細胞と真皮の樹状細胞による抗原の取り込みと,それにつづく,所属リンパ節への移動が重要であることが知られている.2型ヘルパーT細胞応答が優位な炎症反応を誘導した接触性過敏症モデルで,どちらの樹状細胞においてSTAT5がより重要であるかを調べるため,ランゲルハンス細胞において特異的にSTAT5を欠損させたコンディショナルノックアウトマウスを用いた.このノックアウトマウスでも,表皮および真皮におけるランゲルハンス細胞,また,真皮の樹状細胞のサブセットは正常に存在していた.そして,2型ヘルパーT細胞応答が優位な炎症反応を誘導した接触性過敏症モデルにおいて,皮膚の肥厚やインターロイキン4の産生に減少はみられなかった.これらの結果は,真皮の樹状細胞において,STAT5が2型ヘルパーT細胞への分化およびアレルギー性炎症の発症に重要であることを示唆した.
肺組織の炎症反応での樹状細胞におけるSTAT5の役割を調べるため,2型ヘルパーT細胞に依存したアレルギー性の気道炎症モデルを用いて検討を行った.樹状細胞に特異的なSTAT5ノックアウトマウスでは,肺気道肺胞の洗浄液における好酸球の浸潤,および,所属リンパ節におけるインターロイキン4産生T細胞が著しく減少した.一方,インフルエンザAウイルスを介した1型ヘルパーT細胞に依存する肺組織の炎症モデルでは,体重の減少,抗原に特異的なCD4陽性T細胞,CD8陽性T細胞によるインターフェロンγの産生について,野生型マウスと比較して有意な違いがみられなかった.これらの結果は,肺組織の樹状細胞において,STAT5は2型ヘルパーT細胞応答を介した肺気道の炎症において重要であるが,1型ヘルパーT細胞応答においては重要ではないことを示唆した.
樹状細胞に特異的なSTAT5ノックアウトマウスでは樹状細胞への分化は正常であった一方で,2型ヘルパーT細胞応答が減少したことから,アレルギー性炎症の誘導タンパク質の刺激により表皮バリアの細胞から産生され,樹状細胞においてSTAT5を活性化するサイトカインが,2型ヘルパーT細胞応答の優位なアレルギー性炎症の発症に重要であるとの仮説をたてた.そこで,2型ヘルパーT細胞応答が優位な炎症反応を誘導した接触性過敏症モデルにおいて皮膚の局所でのサイトカインの産生を調べたところ,TSLPの著しい発現上昇が認められた一方で,1型ヘルパーT細胞応答の優位な炎症反応を誘導した接触性過敏症モデルの炎症組織ではTSLPの発現の上昇は認められなかった.同様に,アレルギー性炎症を誘導した肺組織においてもTSLPの発現が確認されている6).
TSLP受容体ノックアウトマウスでは,樹状細胞に特異的なSTAT5ノックアウトマウスと同様に,2型ヘルパーT細胞応答に依存した接触性過敏症およびアレルギー性気道炎症の減少することが,筆者らのグループによりすでに報告されている6,7).さらに直接的にTSLPの役割を検討するため,樹状細胞に特異的なSTAT5ノックアウトマウスに外来抗原とTSLPを直接に投与するモデルを用いてアレルギー性の気道炎症反応について調べた8).その結果,肺気道肺胞の洗浄液における好酸球の浸潤,および,インターロイキン4産生T細胞が,野生型マウスと比較して著しく減少した.これらの結果は,アレルギー性炎症の誘導タンパク質の刺激により皮膚や肺組織において産生されたTSLPが,樹状細胞においてSTAT5の活性化を促しアレルギー性炎症を発症させることを示唆した.
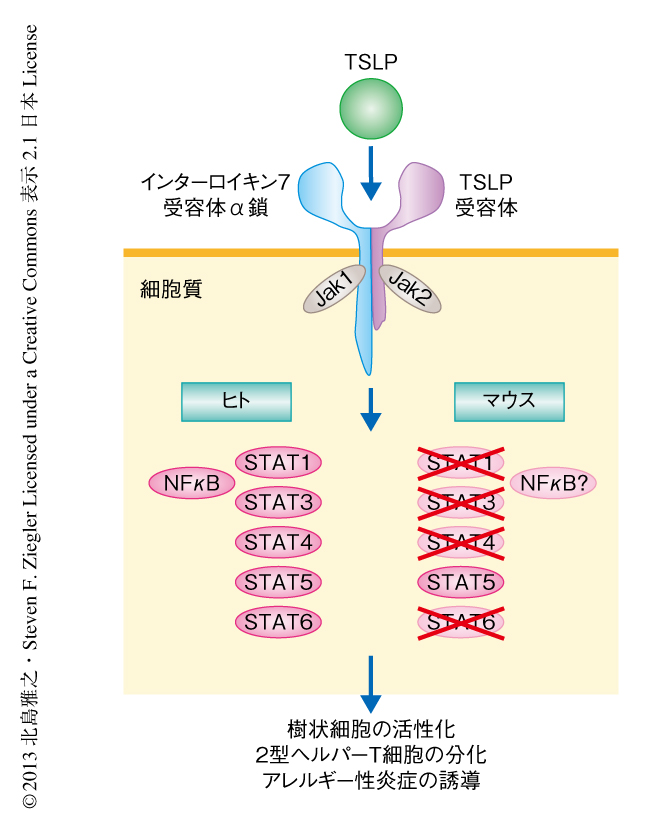
ヒトの末梢血に由来する樹状細胞では,TSLPがSTAT5のほか,STAT1,STAT3,STAT4,STAT6を活性化させること,そして,STAT5の活性化にはシグナル伝達タンパク質であるJak1およびJak2が関与していることが報告されている4).しかし,マウスの樹状細胞において,TSLPがSTAT5のほかの転写因子を活性化するかどうかは明らかでない.そこで以前,筆者らは,TSLPはマウスの骨髄細胞に由来する樹状細胞を活性化し,ヒトの樹状細胞と同様に,いくつかの共刺激分子,ケモカインCCL17の発現の上昇を誘導することを報告した6).また同じ年には,この骨髄細胞に由来する樹状細胞には3つの樹状細胞サブセットが含まれていることが報告された9).そこで,骨髄細胞に由来する樹状細胞を用いて,TSLPにより活性化される樹状細胞サブセットおよび転写因子について解析した.その結果,TSLPはCD11bを強く発現する樹状細胞においてのみJak1,Jak2,STAT5を活性化した.しかし,STAT1,STAT3,STAT4,STAT6はどの樹状細胞においても活性化されなかった.予期されたように,樹状細胞に特異的なSTAT5ノックアウトマウスに由来する樹状細胞は,野生型マウスに由来する樹状細胞と比較して,TSLPによるSTAT5の活性化,それにつづく共刺激分子,ケモカインCCL17の発現の上昇は著しく抑制されていた.STAT1,STAT4,STAT6のそれぞれを欠損したマウスに由来する樹状細胞では,そのような抑制はみられなかった.これらの結果は,TSLPによるマウスの樹状細胞の活性化においてはSTAT5がとくに重要であることを示唆した.
TSLPにより活性化された樹状細胞がアレルギー性炎症に重要な2型ヘルパー細胞の分化を誘導するかどうかを調べるため,骨髄に由来する樹状細胞と,外来抗原,および,その抗原に特異的なCD4陽性T細胞を共培養した.TSLPにより活性化された樹状細胞はCD4陽性T細胞から分化した2型ヘルパーT細胞の放出するサイトカインの産生を増加させたが,STAT5を欠損した樹状細胞ではこの増加は顕著にみられなかった.一方で,STAT5を欠損した樹状細胞は,1型ヘルパーT細胞あるいは2型ヘルパーT細胞を誘導する培養条件において,それぞれのヘルパーT細胞を正常に分化したことから,基本的なT細胞刺激能には問題のないことが示唆された.TSLPと抗原をマウスに投与する実験系を用いても,樹状細胞に特異的なSTAT5ノックアウトマウスでは2型ヘルパーT細胞への分化は抑制されていた.これらの結果は,TSLPを介した樹状細胞におけるSTAT5の活性化が2型ヘルパー細胞の分化に重要であることを示唆した.
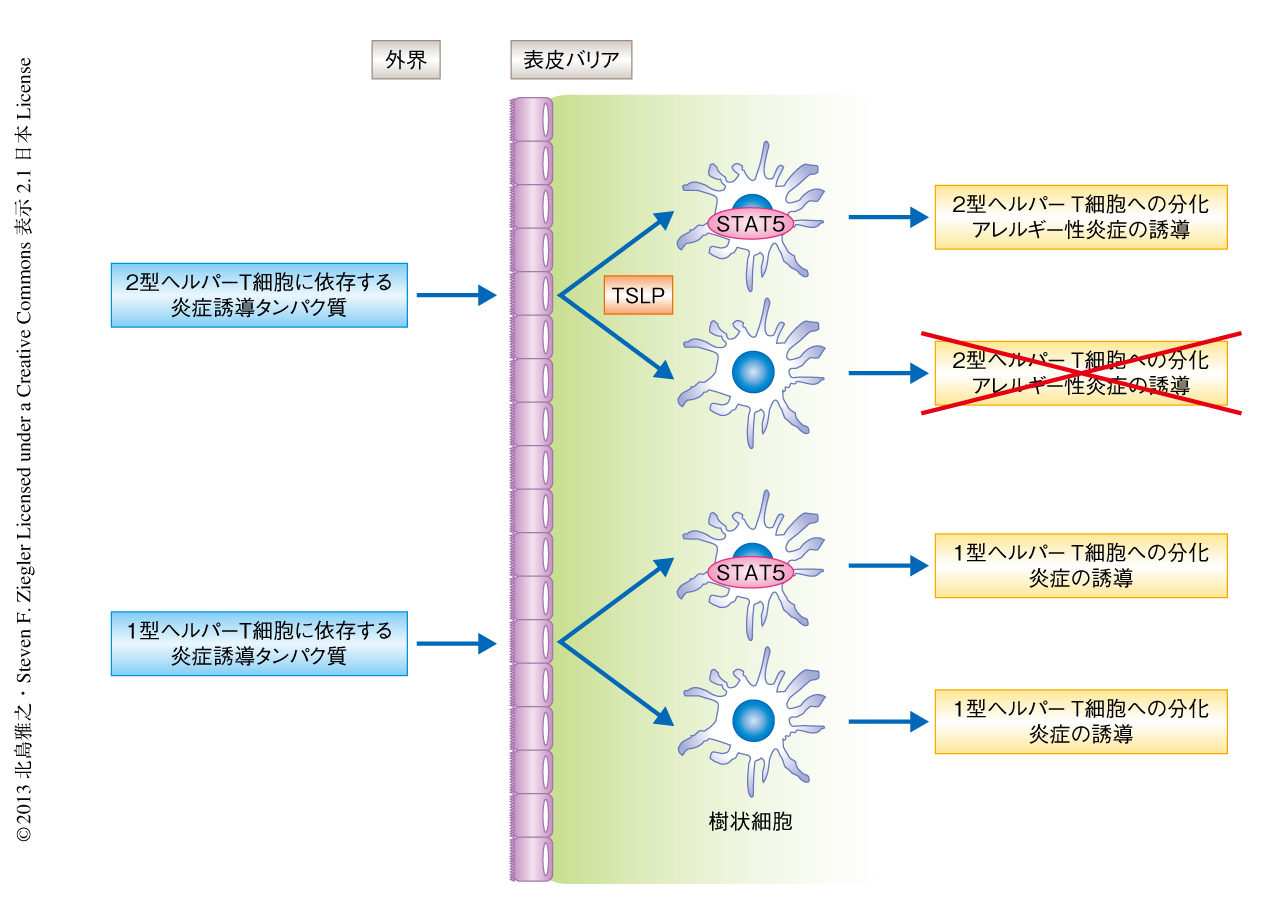
免疫恒常性を維持するため必要とされる樹状細胞によるT細胞の分化制御の分子機構は,多くの点が未解明である.今回の結果から,2型ヘルパーT細胞への分化をともなうアレルギー性炎症の発症には,表皮バリアの細胞から産生されるTSLPを介した樹状細胞におけるSTAT5の活性化が重要であり,1型ヘルパーT細胞に依存する炎症反応においてこれは重要ではないことが明らかにされた(図1).また,ヒトとマウスにおいて,TSLPはアレルギー性炎症の発症に関与することが示唆されているのにもかかわらず,樹状細胞において活性化される転写因子は同じではない可能性も示された(図2).これらの結果は,樹状細胞の存在する組織,あるいは,そのサブセットによりTSLPの作用の異なる可能性を示唆しているのかもしれない.今後,TSLPにより活性化されるヒトとマウスの樹状細胞のサブセット,および,シグナル伝達機構の詳細な解析,そして,活性化したSTAT5による樹状細胞の機能の制御機構を解明することにより,アレルギー性炎症の標的療法への発展も期待できる.
略歴:2005年 千葉工業大学大学院工学研究科 修了,同年 千葉大学大学院医学研究院 産学官連携研究員を経て,2008年より米国Benaroya Research Institute at Virginia MasonにてPostdoctoral Fellow.
研究テーマ:サイトカインTSLPを介した免疫応答.
関心事:免疫学全般.
Steven F. Ziegler
米国Benaroya Research Institute at Virginia MasonにてDirector.
© 2013 北島雅之・Steven F. Ziegler Licensed under CC 表示 2.1 日本
(米国Benaroya Research Institute at Virginia Mason,Immunology Program)
email:北島雅之
DOI: 10.7875/first.author.2013.027
The transcription factor STAT5 is critical in dendritic cells for the development of TH2 but not TH1 responses.
Bryan D. Bell, Masayuki Kitajima, Ryan P. Larson, Thomas A. Stoklasek, Kristen Dang, Kazuhito Sakamoto, Kay-Uwe Wagner, Boris Reizis, Lothar Hennighausen, Steven F. Ziegler
Nature Immunology, 14, 364-371 (2013)
要 約
樹状細胞は自然免疫と獲得免疫とをつなぎ免疫恒常性の維持に重要な役割をはたしているが,その分子機構にはいまだ不明な点が多く残されている.今回,筆者らは,樹状細胞において特異的に転写因子STAT5を欠損したマウスを用いた解析から,樹状細胞においてSTAT5は1型ヘルパーT細胞を必要とする炎症反応には必要なく,2型ヘルパーT細胞を必要とする皮膚や肺組織におけるアレルギー性の炎症反応に重要であることを見い出した.さらに,アレルギー性炎症部位の細胞により産生されSTAT5を活性化するサイトカインとして,TSLPが大きな役割をはたしていることを見い出した.樹状細胞におけるTSLPによるSTAT5の活性化は,1型ヘルパーT細胞の分化ではなく2型ヘルパーT細胞の分化を誘導すること,TSLP受容体を欠損したマウスは,樹状細胞に特異的にSTAT5を欠損したマウスと同様の表現型を示すことも実験的に証明した.これらの結果は,TSLPを介した樹状細胞におけるSTAT5の活性化という方向性が,表皮バリアの細胞におけるアレルギー性炎症に強く関与することを示唆する知見であると考えられた.
はじめに
樹状細胞は外来性および内因性の抗原をT細胞に提示することにより,その抗原に特異的なT細胞の機能を制御して免疫恒常性の一端を担っている.樹状細胞に異常が生じると免疫恒常性は破たんをきたし,感染症,アレルギー性疾患,自己免疫疾患などの発症および増悪につながると考えられる.
転写因子は核において遺伝子の発現制御の役割を直接に担い,細胞の独自性を発揮するのに重要である.造血幹細胞から樹状細胞への分化の過程には,サイトカインFLT3Lを介した転写因子STAT3の機能がとくに重要であることが報告されている1).一方で,同じファミリーに属するSTAT5はコンベンショナルな樹状細胞への分化を亢進する一方で,形質細胞様の樹状細胞への分化を抑制することが報告されているが,生体における役割はいまだ完全には明らかでない2).樹状細胞への分化ののちも,STAT5はTSLP(thymic stromal lymphpoietin,胸腺間質性リンパ球新生因子),インターロイキン7,GM-CSFなどのサイトカインにより活性化されて細胞機能は制御される.とくに,TSLPは樹状細胞だけでなく,ランゲルハンス細胞,B細胞,T細胞などの免疫細胞に作用し,アレルギー性炎症の反応開始に重要であることが報告されている3).TSLPは細胞膜に存在するTSLP受容体とインターロイキン7受容体α鎖により認識され,その結果,STAT5だけでなくさまざまなシグナル伝達タンパク質が活性化される4,5).そのなかで,樹状細胞におけるSTAT5と疾患との直接的なかかわりは明らかになっていない.そこで筆者らは,樹状細胞におけるSTAT5の役割の解明がさまざまな疾患への治療につながることを期待して研究を開始した.
1.STAT5は樹状細胞においてその恒常性の維持に必要ではない
樹状細胞におけるSTAT5の役割を明らかにするため,樹状細胞において特異的にSTAT5を欠損させたコンディショナルノックアウトマウスを用いた.このノックアウトマウスの脾臓組織において,樹状細胞のサブセットおよびほかの免疫細胞は,野生型マウスと比較し正常に存在していた.また,皮膚においても,表皮,真皮,皮膚所属リンパ節における樹状細胞のサブセットの割合は正常であった.さらに,経時的に観察しても自己免疫疾患様の症状を示さなかったことから,この樹状細胞に特異的なSTAT5ノックアウトマウスでは樹状細胞への分化は正常であることが示唆された.
2.樹状細胞においてSTAT5は1型ヘルパー細胞ではなく2型ヘルパー細胞を介した接触性過敏症に必要である
マウスの接触性過敏症はヒトにおける接触性皮膚炎のモデルである.抗原としてハプテンであるフルオレセインイソチオシアネートとフタル酸ジブチルを用いると,2型ヘルパーT細胞応答が優位な炎症反応の誘導されることが知られている.2型ヘルパーT細胞応答が優位な接触性過敏症モデルを誘導した樹状細胞に特異的なSTAT5ノックアウトマウスにおいて,野生型マウスと比較して皮膚の肥厚および炎症性細胞の浸潤が減少していた.さらに,2型ヘルパーT細胞が放出するサイトカインであるインターロイキン4の減少,および,CD4陽性T細胞の増殖の低下も認められた.これらの結果は,STAT5を欠損した樹状細胞では2型ヘルパーT細胞への分化が抑制されていることを示唆した.また,皮膚において抗原であるフルオレセインイソチオシアネートを取り込み所属リンパ節へと移動した樹状細胞の数は,樹状細胞に特異的なSTAT5ノックアウトマウスにおいて有意な減少を示し,樹状細胞においてT細胞刺激タンパク質であるCD86の発現の低下も認められたことから,樹状細胞の活性化が抑制されていることが示唆された.つぎに,ハプテンであるジニトロフルオロベンゼンを用いて,1型ヘルパーT細胞応答が優位な接触性過敏症モデルを誘導した.興味深いことに,このモデルでは樹状細胞に特異的なSTAT5ノックアウトマウスにおいて,野生型マウスと比較して皮膚の肥厚,1型ヘルパーT細胞が放出するサイトカインであるインターフェロンγ,および,CD4陽性T細胞とCD8陽性T細胞の増殖について有意な違いが認められなかった.
マウスの接触性過敏症モデルでは,樹状細胞の一種である表皮のランゲルハンス細胞と真皮の樹状細胞による抗原の取り込みと,それにつづく,所属リンパ節への移動が重要であることが知られている.2型ヘルパーT細胞応答が優位な炎症反応を誘導した接触性過敏症モデルで,どちらの樹状細胞においてSTAT5がより重要であるかを調べるため,ランゲルハンス細胞において特異的にSTAT5を欠損させたコンディショナルノックアウトマウスを用いた.このノックアウトマウスでも,表皮および真皮におけるランゲルハンス細胞,また,真皮の樹状細胞のサブセットは正常に存在していた.そして,2型ヘルパーT細胞応答が優位な炎症反応を誘導した接触性過敏症モデルにおいて,皮膚の肥厚やインターロイキン4の産生に減少はみられなかった.これらの結果は,真皮の樹状細胞において,STAT5が2型ヘルパーT細胞への分化およびアレルギー性炎症の発症に重要であることを示唆した.
3.樹状細胞においてSTAT5は2型ヘルパー細胞を介した肺炎症反応に必要である
肺組織の炎症反応での樹状細胞におけるSTAT5の役割を調べるため,2型ヘルパーT細胞に依存したアレルギー性の気道炎症モデルを用いて検討を行った.樹状細胞に特異的なSTAT5ノックアウトマウスでは,肺気道肺胞の洗浄液における好酸球の浸潤,および,所属リンパ節におけるインターロイキン4産生T細胞が著しく減少した.一方,インフルエンザAウイルスを介した1型ヘルパーT細胞に依存する肺組織の炎症モデルでは,体重の減少,抗原に特異的なCD4陽性T細胞,CD8陽性T細胞によるインターフェロンγの産生について,野生型マウスと比較して有意な違いがみられなかった.これらの結果は,肺組織の樹状細胞において,STAT5は2型ヘルパーT細胞応答を介した肺気道の炎症において重要であるが,1型ヘルパーT細胞応答においては重要ではないことを示唆した.
4.樹状細胞に特異的なSTAT5ノックアウトマウスではTSLPへの応答が減少する
樹状細胞に特異的なSTAT5ノックアウトマウスでは樹状細胞への分化は正常であった一方で,2型ヘルパーT細胞応答が減少したことから,アレルギー性炎症の誘導タンパク質の刺激により表皮バリアの細胞から産生され,樹状細胞においてSTAT5を活性化するサイトカインが,2型ヘルパーT細胞応答の優位なアレルギー性炎症の発症に重要であるとの仮説をたてた.そこで,2型ヘルパーT細胞応答が優位な炎症反応を誘導した接触性過敏症モデルにおいて皮膚の局所でのサイトカインの産生を調べたところ,TSLPの著しい発現上昇が認められた一方で,1型ヘルパーT細胞応答の優位な炎症反応を誘導した接触性過敏症モデルの炎症組織ではTSLPの発現の上昇は認められなかった.同様に,アレルギー性炎症を誘導した肺組織においてもTSLPの発現が確認されている6).
TSLP受容体ノックアウトマウスでは,樹状細胞に特異的なSTAT5ノックアウトマウスと同様に,2型ヘルパーT細胞応答に依存した接触性過敏症およびアレルギー性気道炎症の減少することが,筆者らのグループによりすでに報告されている6,7).さらに直接的にTSLPの役割を検討するため,樹状細胞に特異的なSTAT5ノックアウトマウスに外来抗原とTSLPを直接に投与するモデルを用いてアレルギー性の気道炎症反応について調べた8).その結果,肺気道肺胞の洗浄液における好酸球の浸潤,および,インターロイキン4産生T細胞が,野生型マウスと比較して著しく減少した.これらの結果は,アレルギー性炎症の誘導タンパク質の刺激により皮膚や肺組織において産生されたTSLPが,樹状細胞においてSTAT5の活性化を促しアレルギー性炎症を発症させることを示唆した.
5.樹状細胞においてSTAT5はTSLPにより活性化される
ヒトの末梢血に由来する樹状細胞では,TSLPがSTAT5のほか,STAT1,STAT3,STAT4,STAT6を活性化させること,そして,STAT5の活性化にはシグナル伝達タンパク質であるJak1およびJak2が関与していることが報告されている4).しかし,マウスの樹状細胞において,TSLPがSTAT5のほかの転写因子を活性化するかどうかは明らかでない.そこで以前,筆者らは,TSLPはマウスの骨髄細胞に由来する樹状細胞を活性化し,ヒトの樹状細胞と同様に,いくつかの共刺激分子,ケモカインCCL17の発現の上昇を誘導することを報告した6).また同じ年には,この骨髄細胞に由来する樹状細胞には3つの樹状細胞サブセットが含まれていることが報告された9).そこで,骨髄細胞に由来する樹状細胞を用いて,TSLPにより活性化される樹状細胞サブセットおよび転写因子について解析した.その結果,TSLPはCD11bを強く発現する樹状細胞においてのみJak1,Jak2,STAT5を活性化した.しかし,STAT1,STAT3,STAT4,STAT6はどの樹状細胞においても活性化されなかった.予期されたように,樹状細胞に特異的なSTAT5ノックアウトマウスに由来する樹状細胞は,野生型マウスに由来する樹状細胞と比較して,TSLPによるSTAT5の活性化,それにつづく共刺激分子,ケモカインCCL17の発現の上昇は著しく抑制されていた.STAT1,STAT4,STAT6のそれぞれを欠損したマウスに由来する樹状細胞では,そのような抑制はみられなかった.これらの結果は,TSLPによるマウスの樹状細胞の活性化においてはSTAT5がとくに重要であることを示唆した.
6.樹状細胞におけるTSLPを介したSTAT5の活性化は2型ヘルパーT細胞反応を促進する
TSLPにより活性化された樹状細胞がアレルギー性炎症に重要な2型ヘルパー細胞の分化を誘導するかどうかを調べるため,骨髄に由来する樹状細胞と,外来抗原,および,その抗原に特異的なCD4陽性T細胞を共培養した.TSLPにより活性化された樹状細胞はCD4陽性T細胞から分化した2型ヘルパーT細胞の放出するサイトカインの産生を増加させたが,STAT5を欠損した樹状細胞ではこの増加は顕著にみられなかった.一方で,STAT5を欠損した樹状細胞は,1型ヘルパーT細胞あるいは2型ヘルパーT細胞を誘導する培養条件において,それぞれのヘルパーT細胞を正常に分化したことから,基本的なT細胞刺激能には問題のないことが示唆された.TSLPと抗原をマウスに投与する実験系を用いても,樹状細胞に特異的なSTAT5ノックアウトマウスでは2型ヘルパーT細胞への分化は抑制されていた.これらの結果は,TSLPを介した樹状細胞におけるSTAT5の活性化が2型ヘルパー細胞の分化に重要であることを示唆した.
おわりに
免疫恒常性を維持するため必要とされる樹状細胞によるT細胞の分化制御の分子機構は,多くの点が未解明である.今回の結果から,2型ヘルパーT細胞への分化をともなうアレルギー性炎症の発症には,表皮バリアの細胞から産生されるTSLPを介した樹状細胞におけるSTAT5の活性化が重要であり,1型ヘルパーT細胞に依存する炎症反応においてこれは重要ではないことが明らかにされた(図1).また,ヒトとマウスにおいて,TSLPはアレルギー性炎症の発症に関与することが示唆されているのにもかかわらず,樹状細胞において活性化される転写因子は同じではない可能性も示された(図2).これらの結果は,樹状細胞の存在する組織,あるいは,そのサブセットによりTSLPの作用の異なる可能性を示唆しているのかもしれない.今後,TSLPにより活性化されるヒトとマウスの樹状細胞のサブセット,および,シグナル伝達機構の詳細な解析,そして,活性化したSTAT5による樹状細胞の機能の制御機構を解明することにより,アレルギー性炎症の標的療法への発展も期待できる.
文 献
- Shortman, K. & Naik, S. H.: Steady-state and inflammatory dendritic-cell development. Nat. Rev. Immunol., 7, 19-30 (2007)[PubMed]
- Esashi, E., Wang, Y. H., Perng, O. et al.: The signal transducer STAT5 inhibits plasmacytoid dendritic cell development by suppressing transcription factor IRF8. Immunity, 28, 509-520 (2008)[PubMed]
- Ziegler, S. F. & Artis, D.: Sensing the outside world: TSLP regulates barrier immunity. Nat. Immunol., 11, 289-293 (2010)[PubMed]
- Arima, K., Watanabe, N., Hanabuchi, S. et al.: Distinct signal codes generate dendritic cell functional plasticity. Sci. Signal., 3, ra4 (2010)[PubMed]
- Rochman, Y., Kashyap, M., Robinson, G. W. et al.: Thymic stromal lymphopoietin-mediated STAT5 phosphorylation via kinases JAK1 and JAK2 reveals a key difference from IL-7-induced signaling. Proc. Natl. Acad. Sci. USA, 107, 19455-19460 (2010)[PubMed]
- Zhou, B., Comeau, M. R., De Smedt, T. et al.: Thymic stromal lymphopoietin as a key initiator of allergic airway inflammation in mice. Nat. Immunol., 6, 1047-1053 (2005)[PubMed]
- Larson, R. P., Zimmerli, S. C., Comeau, M. R. et al.: Dibutyl phthalate-induced thymic stromal lymphopoietin is required for Th2 contact hypersensitivity responses. J. Immunol., 184, 2974-2984 (2010)[PubMed]
- Headley, M. B., Zhou, B., Shih, W. X. et al.: TSLP conditions the lung immune environment for the generation of pathogenic innate and antigen-specific adaptive immune responses. J. Immunol., 182, 1641-1647 (2009)[PubMed]
- Naik, S. H., Proietto, A. I., Wilson, N. S. et al.: Cutting edge: generation of splenic CD8+ and CD8- dendritic cell equivalents in Fms-like tyrosine kinase 3 ligand bone marrow cultures. J. Immunol., 174, 6592-6597 (2005)[PubMed]
著者プロフィール
略歴:2005年 千葉工業大学大学院工学研究科 修了,同年 千葉大学大学院医学研究院 産学官連携研究員を経て,2008年より米国Benaroya Research Institute at Virginia MasonにてPostdoctoral Fellow.
研究テーマ:サイトカインTSLPを介した免疫応答.
関心事:免疫学全般.
Steven F. Ziegler
米国Benaroya Research Institute at Virginia MasonにてDirector.
© 2013 北島雅之・Steven F. Ziegler Licensed under CC 表示 2.1 日本
