ヌタウナギの頭部の発生と脊椎動物の進化
大石康博・倉谷 滋
(理化学研究所発生再生科学総合研究センター 形態進化研究グループ)
email:大石康博,倉谷 滋
DOI: 10.7875/first.author.2013.004
Craniofacial development of hagfishes and the evolution of vertebrates.
Yasuhiro Oisi, Kinya G. Ota, Shigehiro Kuraku, Satoko Fujimoto, Shigeru Kuratani
Nature, 493, 175-180 (2013)
現生の無顎類であるヌタウナギとヤツメウナギは,脊椎動物のもっとも原始的な系統である円口類を構成する.最近の分子的な解析は円口類の単系統性をくり返し支持してきているが,一方で,形態学的な観察から,ヌタウナギが脊椎動物の基本的な特徴を欠き,さらに,その腺性下垂体は内胚葉に由来すると報告されており,ヤツメウナギを含むすべての脊椎動物のなかでもっとも祖先的な系統であるとも解釈されてきた.また,ヌタウナギとヤツメウナギとで,成体の頭部の形態は容易には比較できない.そこで,筆者らは,ヌタウナギの胚の発生系列を用い,その頭部の構造の発生過程を詳細に観察した.その結果,ヌタウナギの腺性下垂体は外胚葉に由来することが明らかになった.また,ヌタウナギとヤツメウナギは顎口類とは異なる汎円口類パターンを共有することが個体発生の比較から明らかになった.この結果は,分子系統学の示す系統関係と整合的であり,さらに,古生代の化石魚類の知見を考慮すると,汎円口類パターンは現生する脊椎動物全体の祖先に共有されていた可能性を示唆した.
最近の分子系統学的な研究成果から,ヌタウナギとヤツメウナギは,約5億年前に顎口類のグループから分岐した単系統群(ひとつの共通祖先から進化した生物群)を構成すると考えられている1,2).このグループは,顎の代わりに丸い口をもち,鼻孔が1つしかないという特徴から円口類とよばれる3).対して,現生するほとんどの脊椎動物である顎口類は,顎の存在と対をなす鼻孔により特徴づけられる3).しかし,形態学者の見解は分子系統学が提示した系統関係とは矛盾しており,ヤツメウナギと顎口類の共通祖先から最初にヌタウナギが分かれたと考えてきた.つまり,ヌタウナギは現生するもっとも祖先的な脊椎動物であり,むしろ,ヤツメウナギと顎口類が近縁であることになる4).その根拠として,ヌタウナギは脊椎動物を特徴づけるいくつかの形態を欠いていることがあげられる.ヌタウナギは明瞭な背骨や眼のレンズをもたない4).また,ほかの脊椎動物の腺性下垂体は外胚葉に由来するのに対し,ヌタウナギの腺性下垂体は,原始的な脊索動物であるナメクジウオの腺性下垂体に相当する構造と同様に,内胚葉に由来すると報告されている5,6).さらに,成体のヌタウナギとヤツメウナギの頭部の構造は著しく異なるため,解剖学的な比較も容易ではない.
そのため,ヌタウナギを比較発生的に解析する重要性は以前から認識されていた.しかし,ヌタウナギは深海性で,生殖に関する生理および生態はほとんど不明であり,発生に関する情報はかぎられたものだった.筆者らの研究室では,2007年,実験室において日本産のヌタウナギ(Eptatretus burgeri)の受精卵を得る方法,また,組織学的および分子生物学的な実験の系を世界にさきがけて確立し7),その成果として,ヌタウナギの胚は,脊椎動物の体制を特徴づける移動性の神経堤細胞を分化し7),さらに,背骨の原基となる硬節細胞をもつことを明らかにした8).今回,筆者らは,ヌタウナギの頭部の発生に注目し,その腺性下垂体の由来を観察し,さらに,ヌタウナギの頭部の構造を比較発生学的に解析することにより,初期の脊椎動物における頭部の形態の進化における理解を深めることを試みた.
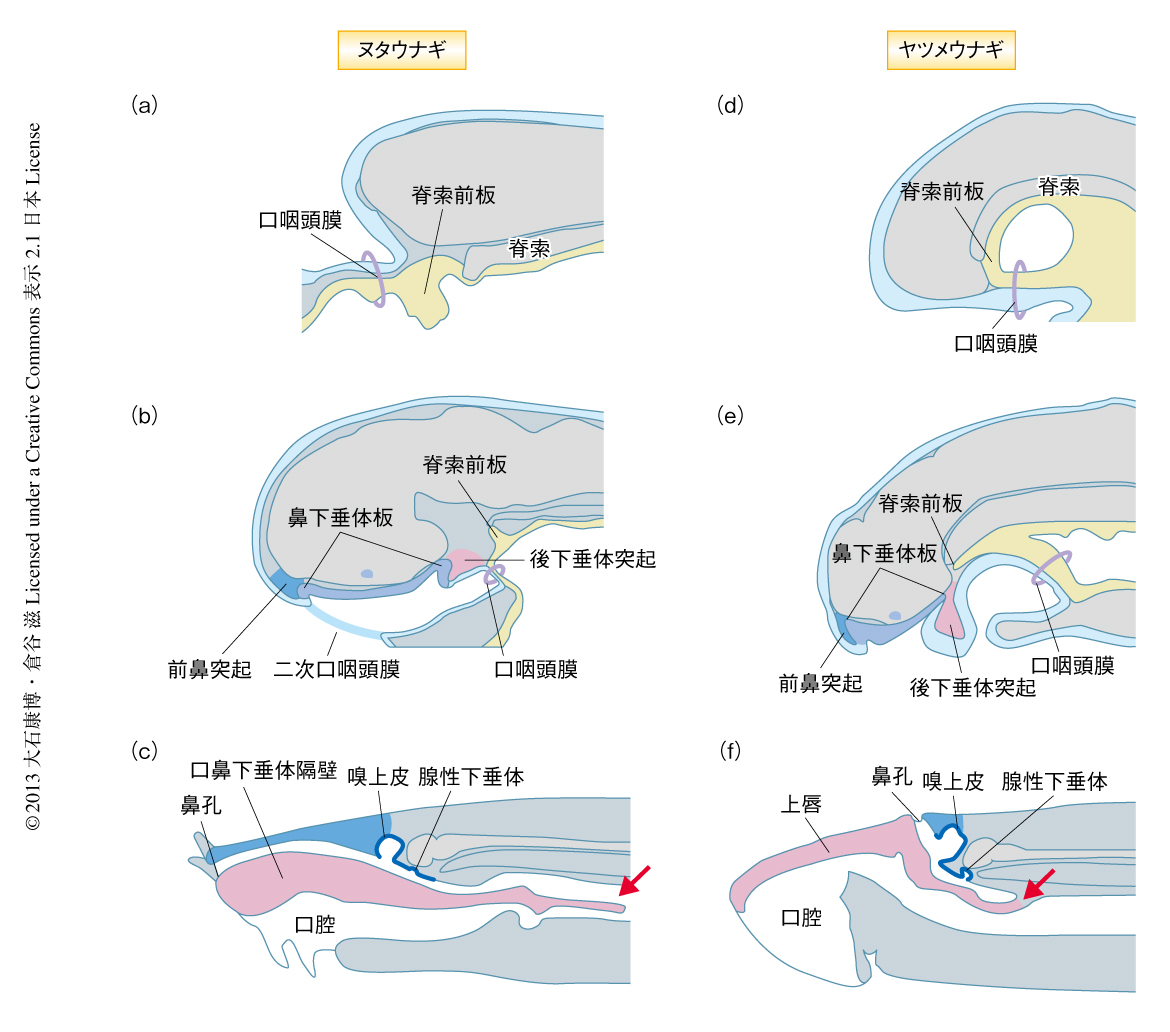
ヌタウナギの腺性下垂体の由来を明らかにするため,ほかのすべての脊椎動物の胚の頭部において外胚葉と内胚葉との境界を形成する口咽頭膜6) をヌタウナギの胚において同定し,口咽頭膜と腺性下垂体原基(下垂体プラコード)との位置関係を調べた.その結果,発生の初期に,前脳の前方に外胚葉と内胚葉の上皮とが隣接した口咽頭膜が観察された(図1a).つづいて,口咽頭膜は前脳の下部へ陥入し,その前方に二次的な膜が形成された(図1b).以前の報告では,この二次的な膜(二次口咽頭膜)が口咽頭膜であると誤って同定され,そのため,ヌタウナギの下垂体プラコードは内胚葉に由来すると解釈された5).しかし,ヌタウナギの下垂体プラコードは明らかに真の口咽頭膜の前方に位置していた(図1b).これらの結果から,ヌタウナギの腺性下垂体は外胚葉に由来すると思われた(図1c).
現在まで調べられたすべての脊椎動物において,下垂体は神経性下垂体と腺性下垂体よりなる.そのうち,神経性下垂体の発生には前脳の腹側に位置する視床下部におけるShh/Nkx2.1遺伝子の発現が必須であることがわかっている.一方で,腺性下垂体は視床下部に隣接して発生する外胚葉性の肥厚(下垂体プラコード)より生じ,Pitx2遺伝子とLhx3/4遺伝子の発現がその発生に重要な役割をはたすことが報告されている.そこで,下垂体の発生部位を明らかにすることにより,腺性下垂体の由来につきより詳細な解析を試みた.
ヌタウナギから1種類のShh遺伝子,Nkx2.1遺伝子,Pitx2遺伝子,Lhx3/4遺伝子の相同遺伝子をクローニングし,それぞれ,EbHh1遺伝子,EbNkx2.1遺伝子,EbPitxA遺伝子,EbLhx3/4A遺伝子と名づけた.in situハイブリダイゼーション法によりヌタウナギの胚におけるこれら遺伝子の発現を調べたところ,EbHh1遺伝子およびEbNkx2.1遺伝子は前脳の腹側の視床下部において発現していた.また,視床と隣接する下垂体プラコードにおいてEbPitxA遺伝子の発現および弱いEbLhx3/4遺伝子の発現が観察された.これらの観察から,ヌタウナギの腺性下垂体はほかの脊椎動物と同様な遺伝子の発現機構にもとづき外胚葉から生じることが示唆された.
つづいて,発生をつうじて隣接していた視床下部と下垂体プラコードから,神経性下垂体と腺性下垂体が生じた(図1c).これらの結果から,ヌタウナギの腺性下垂体は,ほかの脊椎動物と同様に,外胚葉に由来することが明らかになった.
以前の研究では,ヌタウナギの腺性下垂体は脊索の前端において視床に隣接する内胚葉性の肥厚に由来すると報告された5).それをうけて,ヌタウナギの内胚葉の細胞が腺性下垂体の形成に関与するかどうかを調べた.組織学的には,この内胚葉の部位はほかの脊椎動物の胚にも観察され脊索前板とよばれる(図1d).脊索前板のマーカーとしては,Shh遺伝子およびSix3/6遺伝子の発現が報告されている.そこで,ヌタウナギにおけるこれらの相同遺伝子の発現を調べたところ,確かに内胚葉性の肥厚はEbHh1遺伝子およびEbSix3/6A遺伝子を発現していた.さらに,発生の初期,ヌタウナギの脊索前板は腺性下垂体に隣接するが(図1a),つづいてそれは視床下部から分離し(図1b),消失する.これらの結果から,ヌタウナギの内胚葉は腺性下垂体の形成には関与しないことが明らかになった.
ヤツメウナギと顎口類の頭部は形態学的に異なる発生パターンを示すことが知られている.なかでも,鼻プラコードと下垂体プラコード(嗅上皮と腺性下垂体の原基に相当する外胚葉性の肥厚)との位置関係が重要である.ヤツメウナギでは鼻プラコードと下垂体プラコードとが連続した鼻下垂体板を形成する(図1e).一方で,顎口類では2つの鼻プラコードと下垂体プラコードは離れて位置する9).以前の研究より,ヌタウナギの胚の前脳の下部中央において前後に連続した外胚葉性の肥厚が観察され,鼻下垂体板に相当すると考えられていた(図1b).そこで,ヌタウナギの下垂体プラコードと鼻プラコードの発生パターンを調べることにした.
Fgf8/17遺伝子は顎口類の鼻プラコード,あるいは,ヤツメウナギの鼻下垂体板の前方に発現し,その分化に重要な役割をはたす.そこで,ヌタウナギからFgf8/17遺伝子の相同遺伝子をクローニングしEbFgf8/17遺伝子と名づけた.EbFgf8/17遺伝子の発現を解析したところ,鼻下垂体板の前部に強く発現していた.一方,ヌタウナギの下垂体プラコードに発現するEbPitxA遺伝子は,鼻下垂体板の後部において発現していた.また,顎口類において鼻プラコードおよび下垂体プラコードの両方に発現することが知られるSix3/6遺伝子およびSox2/3遺伝子のヌタウナギにおける相同遺伝子であるEbSix3/6A遺伝子およびEbSoxB1遺伝子は,鼻下垂体板の全域に発現していた.これらの結果は,ヌタウナギにおいても鼻プラコードと下垂体プラコードとが連続し,鼻下垂体板を形成していることを示唆した.
また,ヌタウナギの鼻下垂体板は,ヤツメウナギと同様の分化を示した.ヤツメウナギおよびヌタウナギにおいて,鼻下垂体板は1つの鼻孔につながる鼻腔(鼻下垂体腔)を形成した.また,鼻腔の背側では無対の嗅上皮と腺性下垂体が隣接して形成された(図1 c, f).したがって,ヌタウナギとヤツメウナギの鼻下垂体板は形態発生学的にも相同であることが示唆された.
鼻下垂体板と密接な関係をもつ頭部の構造のパターンも,ヌタウナギの胚とヤツメウナギの胚において相同である可能性がある9).そこで,両者の頭部の構造の発生を比較することにした.まず,組織切片によりヌタウナギとヤツメウナギの頭部の内部構造を解析したところ,鼻下垂体板の前後に間葉からなる突起が観察された.そのうち,鼻下垂体板の前方を前鼻突起,鼻下垂体板の後方を後下垂体突起とよんだ(図1 b, e).つぎに,三次元的に観察したところ,ヌタウナギとヤツメウナギの後下垂体突起は鼻下垂体板の後部(下垂体プラコード)の後方に一対の突起として生じた.つづいて,左右の後下垂体突起は正中で癒合をはじめ,鼻下垂体板を含む鼻腔と口腔が分離した(図1 b, e).つづいて,ヌタウナギでは後下垂体突起の前部が4対の触手を形成した(図1c)が,一方で,ヤツメウナギの後下垂体突起は上唇を形成した(図1f).ヌタウナギの触手とヤツメウナギの上唇の構造は大きく異なるが,三叉神経による神経支配のパターンは両者が等価な構造であることを支持した.また,後下垂体突起の後方において,ヌタウナギの鼻腔は咽頭へつながり,その後端が盲嚢となっているヤツメウナギとは異なるパターンをもっていた(図1 c, f).形態発生的な観察から,ヌタウナギに特有なこの連絡孔は,後下垂体突起に由来する構造(触手へとつづく口鼻下垂体隔壁)の後端が二次的に破れることによりもたらされることがわかった(図1 b, c).したがって,ヌタウナギの触手および口鼻下垂体隔壁と,ヤツメウナギの上唇は,後下垂体突起に由来する相同な構造であった.
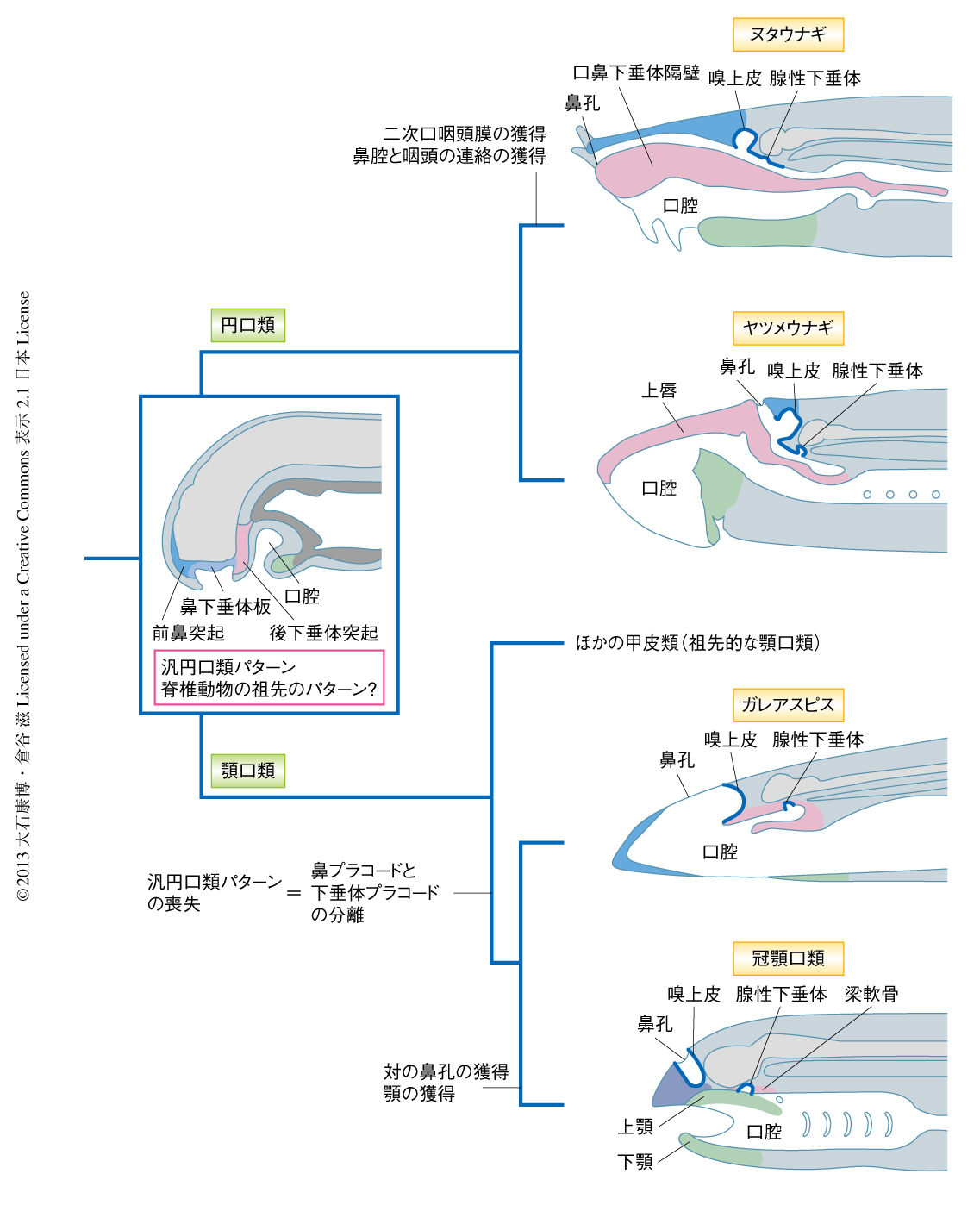
ヌタウナギの胚とヤツメウナギの胚に共有される頭部の構造における基本パターンは,顎口類とは異なっていた.ヌタウナギとヤツメウナギの後下垂体突起は口部の構造を形成する一方,顎口類の胚において対応する位置をしめる顎前領域からは頭蓋を構成する梁軟骨が生じる.このことは,ヌタウナギの胚とヤツメウナギの胚は,顎口類の胚とは異なり,独特の汎円口類パターンを基盤に発生することを示唆した(図2).
筆者らは,ヌタウナギの腺性下垂体がほかの脊椎動物と同様に外胚葉の由来であることを明らかにした.最近,ヌタウナギの腺性下垂体はほかの脊椎動物の腺性下垂体と等価の形態と機能をもち,ヌタウナギの祖先的な形質の多くは二次的な変形や消失にもとづくことが示されてきた2,8).今回の結果は,ヌタウナギの形態についての数々の再解釈とともに,ヌタウナギとヤツメウナギが1つのグループである円口類を構成することを分子系統学のデータと整合的に支持した.
ヌタウナギとヤツメウナギの成体の頭部の形態は大きく異なっている.しかし,今回の比較発生学的な解析により,両者は鼻下垂体板,前鼻突起,後下垂体突起という頭部の基本原基からなる汎円口類パターンを共有することが明らかにされた.ヌタウナギとヤツメウナギでは腺性下垂体は鼻下垂体板にもとづいており,無対の嗅上皮に隣接していた.一方で,顎口類の胚では,下垂体プラコードは対の鼻プラコードから分離し,二次的に口腔に取り込まれる.絶滅した無顎類である甲皮類は,軟骨性の骨格しかもたない円口類とは異なり体を硬骨でおおっていたことから,円口類よりも顎口類と近縁な系統とみなされている3).なかでも,ガレアスピスは注目すべき形質を示している10).ガレアスピスの鼻孔は円口類のように正中に1つしかない.一方で,ガレアスピスは対の嗅上皮をもち,さらに,腺性下垂体と嗅上皮とが顎口類と同じように分離していた可能性が示されている.今回の発生学的な研究と化石の記録から,汎円口類パターンがすべての脊椎動物の祖先的な発生プログラムを反映していると仮定すると,ガレアスピスは円口類パターンの部分的な喪失をともない,顎口類へといたる遷移段階を示している可能性が浮上する(図2).今後さらに,円口類と顎口類との比較発生学,また,化石の記録の解析が進展することにより,初期の脊椎動物が対鼻孔や顎を獲得した進化機構が明らかになることが期待される.
略歴:神戸大学大学院理学研究科博士課程 在学中.
研究テーマ:動物の形態要素,構造からみた発生と進化.
倉谷 滋(Shigeru Kuratani)
理化学研究所発生再生科学総合研究センター グループディレクター.
研究室URL:http://www.cdb.riken.jp/emo/japanese/indexj.html
© 2013 大石康博・倉谷 滋 Licensed under CC 表示 2.1 日本
(理化学研究所発生再生科学総合研究センター 形態進化研究グループ)
email:大石康博,倉谷 滋
DOI: 10.7875/first.author.2013.004
Craniofacial development of hagfishes and the evolution of vertebrates.
Yasuhiro Oisi, Kinya G. Ota, Shigehiro Kuraku, Satoko Fujimoto, Shigeru Kuratani
Nature, 493, 175-180 (2013)
要 約
現生の無顎類であるヌタウナギとヤツメウナギは,脊椎動物のもっとも原始的な系統である円口類を構成する.最近の分子的な解析は円口類の単系統性をくり返し支持してきているが,一方で,形態学的な観察から,ヌタウナギが脊椎動物の基本的な特徴を欠き,さらに,その腺性下垂体は内胚葉に由来すると報告されており,ヤツメウナギを含むすべての脊椎動物のなかでもっとも祖先的な系統であるとも解釈されてきた.また,ヌタウナギとヤツメウナギとで,成体の頭部の形態は容易には比較できない.そこで,筆者らは,ヌタウナギの胚の発生系列を用い,その頭部の構造の発生過程を詳細に観察した.その結果,ヌタウナギの腺性下垂体は外胚葉に由来することが明らかになった.また,ヌタウナギとヤツメウナギは顎口類とは異なる汎円口類パターンを共有することが個体発生の比較から明らかになった.この結果は,分子系統学の示す系統関係と整合的であり,さらに,古生代の化石魚類の知見を考慮すると,汎円口類パターンは現生する脊椎動物全体の祖先に共有されていた可能性を示唆した.
はじめに
最近の分子系統学的な研究成果から,ヌタウナギとヤツメウナギは,約5億年前に顎口類のグループから分岐した単系統群(ひとつの共通祖先から進化した生物群)を構成すると考えられている1,2).このグループは,顎の代わりに丸い口をもち,鼻孔が1つしかないという特徴から円口類とよばれる3).対して,現生するほとんどの脊椎動物である顎口類は,顎の存在と対をなす鼻孔により特徴づけられる3).しかし,形態学者の見解は分子系統学が提示した系統関係とは矛盾しており,ヤツメウナギと顎口類の共通祖先から最初にヌタウナギが分かれたと考えてきた.つまり,ヌタウナギは現生するもっとも祖先的な脊椎動物であり,むしろ,ヤツメウナギと顎口類が近縁であることになる4).その根拠として,ヌタウナギは脊椎動物を特徴づけるいくつかの形態を欠いていることがあげられる.ヌタウナギは明瞭な背骨や眼のレンズをもたない4).また,ほかの脊椎動物の腺性下垂体は外胚葉に由来するのに対し,ヌタウナギの腺性下垂体は,原始的な脊索動物であるナメクジウオの腺性下垂体に相当する構造と同様に,内胚葉に由来すると報告されている5,6).さらに,成体のヌタウナギとヤツメウナギの頭部の構造は著しく異なるため,解剖学的な比較も容易ではない.
そのため,ヌタウナギを比較発生的に解析する重要性は以前から認識されていた.しかし,ヌタウナギは深海性で,生殖に関する生理および生態はほとんど不明であり,発生に関する情報はかぎられたものだった.筆者らの研究室では,2007年,実験室において日本産のヌタウナギ(Eptatretus burgeri)の受精卵を得る方法,また,組織学的および分子生物学的な実験の系を世界にさきがけて確立し7),その成果として,ヌタウナギの胚は,脊椎動物の体制を特徴づける移動性の神経堤細胞を分化し7),さらに,背骨の原基となる硬節細胞をもつことを明らかにした8).今回,筆者らは,ヌタウナギの頭部の発生に注目し,その腺性下垂体の由来を観察し,さらに,ヌタウナギの頭部の構造を比較発生学的に解析することにより,初期の脊椎動物における頭部の形態の進化における理解を深めることを試みた.
1.ヌタウナギの腺性下垂体の由来:組織の観察
ヌタウナギの腺性下垂体の由来を明らかにするため,ほかのすべての脊椎動物の胚の頭部において外胚葉と内胚葉との境界を形成する口咽頭膜6) をヌタウナギの胚において同定し,口咽頭膜と腺性下垂体原基(下垂体プラコード)との位置関係を調べた.その結果,発生の初期に,前脳の前方に外胚葉と内胚葉の上皮とが隣接した口咽頭膜が観察された(図1a).つづいて,口咽頭膜は前脳の下部へ陥入し,その前方に二次的な膜が形成された(図1b).以前の報告では,この二次的な膜(二次口咽頭膜)が口咽頭膜であると誤って同定され,そのため,ヌタウナギの下垂体プラコードは内胚葉に由来すると解釈された5).しかし,ヌタウナギの下垂体プラコードは明らかに真の口咽頭膜の前方に位置していた(図1b).これらの結果から,ヌタウナギの腺性下垂体は外胚葉に由来すると思われた(図1c).
2.ヌタウナギの腺性下垂体の由来:遺伝子発現の観察
現在まで調べられたすべての脊椎動物において,下垂体は神経性下垂体と腺性下垂体よりなる.そのうち,神経性下垂体の発生には前脳の腹側に位置する視床下部におけるShh/Nkx2.1遺伝子の発現が必須であることがわかっている.一方で,腺性下垂体は視床下部に隣接して発生する外胚葉性の肥厚(下垂体プラコード)より生じ,Pitx2遺伝子とLhx3/4遺伝子の発現がその発生に重要な役割をはたすことが報告されている.そこで,下垂体の発生部位を明らかにすることにより,腺性下垂体の由来につきより詳細な解析を試みた.
ヌタウナギから1種類のShh遺伝子,Nkx2.1遺伝子,Pitx2遺伝子,Lhx3/4遺伝子の相同遺伝子をクローニングし,それぞれ,EbHh1遺伝子,EbNkx2.1遺伝子,EbPitxA遺伝子,EbLhx3/4A遺伝子と名づけた.in situハイブリダイゼーション法によりヌタウナギの胚におけるこれら遺伝子の発現を調べたところ,EbHh1遺伝子およびEbNkx2.1遺伝子は前脳の腹側の視床下部において発現していた.また,視床と隣接する下垂体プラコードにおいてEbPitxA遺伝子の発現および弱いEbLhx3/4遺伝子の発現が観察された.これらの観察から,ヌタウナギの腺性下垂体はほかの脊椎動物と同様な遺伝子の発現機構にもとづき外胚葉から生じることが示唆された.
つづいて,発生をつうじて隣接していた視床下部と下垂体プラコードから,神経性下垂体と腺性下垂体が生じた(図1c).これらの結果から,ヌタウナギの腺性下垂体は,ほかの脊椎動物と同様に,外胚葉に由来することが明らかになった.
3.ヌタウナギの内胚葉の発生
以前の研究では,ヌタウナギの腺性下垂体は脊索の前端において視床に隣接する内胚葉性の肥厚に由来すると報告された5).それをうけて,ヌタウナギの内胚葉の細胞が腺性下垂体の形成に関与するかどうかを調べた.組織学的には,この内胚葉の部位はほかの脊椎動物の胚にも観察され脊索前板とよばれる(図1d).脊索前板のマーカーとしては,Shh遺伝子およびSix3/6遺伝子の発現が報告されている.そこで,ヌタウナギにおけるこれらの相同遺伝子の発現を調べたところ,確かに内胚葉性の肥厚はEbHh1遺伝子およびEbSix3/6A遺伝子を発現していた.さらに,発生の初期,ヌタウナギの脊索前板は腺性下垂体に隣接するが(図1a),つづいてそれは視床下部から分離し(図1b),消失する.これらの結果から,ヌタウナギの内胚葉は腺性下垂体の形成には関与しないことが明らかになった.
4.ヌタウナギの鼻下垂体の発生
ヤツメウナギと顎口類の頭部は形態学的に異なる発生パターンを示すことが知られている.なかでも,鼻プラコードと下垂体プラコード(嗅上皮と腺性下垂体の原基に相当する外胚葉性の肥厚)との位置関係が重要である.ヤツメウナギでは鼻プラコードと下垂体プラコードとが連続した鼻下垂体板を形成する(図1e).一方で,顎口類では2つの鼻プラコードと下垂体プラコードは離れて位置する9).以前の研究より,ヌタウナギの胚の前脳の下部中央において前後に連続した外胚葉性の肥厚が観察され,鼻下垂体板に相当すると考えられていた(図1b).そこで,ヌタウナギの下垂体プラコードと鼻プラコードの発生パターンを調べることにした.
Fgf8/17遺伝子は顎口類の鼻プラコード,あるいは,ヤツメウナギの鼻下垂体板の前方に発現し,その分化に重要な役割をはたす.そこで,ヌタウナギからFgf8/17遺伝子の相同遺伝子をクローニングしEbFgf8/17遺伝子と名づけた.EbFgf8/17遺伝子の発現を解析したところ,鼻下垂体板の前部に強く発現していた.一方,ヌタウナギの下垂体プラコードに発現するEbPitxA遺伝子は,鼻下垂体板の後部において発現していた.また,顎口類において鼻プラコードおよび下垂体プラコードの両方に発現することが知られるSix3/6遺伝子およびSox2/3遺伝子のヌタウナギにおける相同遺伝子であるEbSix3/6A遺伝子およびEbSoxB1遺伝子は,鼻下垂体板の全域に発現していた.これらの結果は,ヌタウナギにおいても鼻プラコードと下垂体プラコードとが連続し,鼻下垂体板を形成していることを示唆した.
また,ヌタウナギの鼻下垂体板は,ヤツメウナギと同様の分化を示した.ヤツメウナギおよびヌタウナギにおいて,鼻下垂体板は1つの鼻孔につながる鼻腔(鼻下垂体腔)を形成した.また,鼻腔の背側では無対の嗅上皮と腺性下垂体が隣接して形成された(図1 c, f).したがって,ヌタウナギとヤツメウナギの鼻下垂体板は形態発生学的にも相同であることが示唆された.
5.ヌタウナギとヤツメウナギの顔面の構造の相同性
鼻下垂体板と密接な関係をもつ頭部の構造のパターンも,ヌタウナギの胚とヤツメウナギの胚において相同である可能性がある9).そこで,両者の頭部の構造の発生を比較することにした.まず,組織切片によりヌタウナギとヤツメウナギの頭部の内部構造を解析したところ,鼻下垂体板の前後に間葉からなる突起が観察された.そのうち,鼻下垂体板の前方を前鼻突起,鼻下垂体板の後方を後下垂体突起とよんだ(図1 b, e).つぎに,三次元的に観察したところ,ヌタウナギとヤツメウナギの後下垂体突起は鼻下垂体板の後部(下垂体プラコード)の後方に一対の突起として生じた.つづいて,左右の後下垂体突起は正中で癒合をはじめ,鼻下垂体板を含む鼻腔と口腔が分離した(図1 b, e).つづいて,ヌタウナギでは後下垂体突起の前部が4対の触手を形成した(図1c)が,一方で,ヤツメウナギの後下垂体突起は上唇を形成した(図1f).ヌタウナギの触手とヤツメウナギの上唇の構造は大きく異なるが,三叉神経による神経支配のパターンは両者が等価な構造であることを支持した.また,後下垂体突起の後方において,ヌタウナギの鼻腔は咽頭へつながり,その後端が盲嚢となっているヤツメウナギとは異なるパターンをもっていた(図1 c, f).形態発生的な観察から,ヌタウナギに特有なこの連絡孔は,後下垂体突起に由来する構造(触手へとつづく口鼻下垂体隔壁)の後端が二次的に破れることによりもたらされることがわかった(図1 b, c).したがって,ヌタウナギの触手および口鼻下垂体隔壁と,ヤツメウナギの上唇は,後下垂体突起に由来する相同な構造であった.
ヌタウナギの胚とヤツメウナギの胚に共有される頭部の構造における基本パターンは,顎口類とは異なっていた.ヌタウナギとヤツメウナギの後下垂体突起は口部の構造を形成する一方,顎口類の胚において対応する位置をしめる顎前領域からは頭蓋を構成する梁軟骨が生じる.このことは,ヌタウナギの胚とヤツメウナギの胚は,顎口類の胚とは異なり,独特の汎円口類パターンを基盤に発生することを示唆した(図2).
おわりに
筆者らは,ヌタウナギの腺性下垂体がほかの脊椎動物と同様に外胚葉の由来であることを明らかにした.最近,ヌタウナギの腺性下垂体はほかの脊椎動物の腺性下垂体と等価の形態と機能をもち,ヌタウナギの祖先的な形質の多くは二次的な変形や消失にもとづくことが示されてきた2,8).今回の結果は,ヌタウナギの形態についての数々の再解釈とともに,ヌタウナギとヤツメウナギが1つのグループである円口類を構成することを分子系統学のデータと整合的に支持した.
ヌタウナギとヤツメウナギの成体の頭部の形態は大きく異なっている.しかし,今回の比較発生学的な解析により,両者は鼻下垂体板,前鼻突起,後下垂体突起という頭部の基本原基からなる汎円口類パターンを共有することが明らかにされた.ヌタウナギとヤツメウナギでは腺性下垂体は鼻下垂体板にもとづいており,無対の嗅上皮に隣接していた.一方で,顎口類の胚では,下垂体プラコードは対の鼻プラコードから分離し,二次的に口腔に取り込まれる.絶滅した無顎類である甲皮類は,軟骨性の骨格しかもたない円口類とは異なり体を硬骨でおおっていたことから,円口類よりも顎口類と近縁な系統とみなされている3).なかでも,ガレアスピスは注目すべき形質を示している10).ガレアスピスの鼻孔は円口類のように正中に1つしかない.一方で,ガレアスピスは対の嗅上皮をもち,さらに,腺性下垂体と嗅上皮とが顎口類と同じように分離していた可能性が示されている.今回の発生学的な研究と化石の記録から,汎円口類パターンがすべての脊椎動物の祖先的な発生プログラムを反映していると仮定すると,ガレアスピスは円口類パターンの部分的な喪失をともない,顎口類へといたる遷移段階を示している可能性が浮上する(図2).今後さらに,円口類と顎口類との比較発生学,また,化石の記録の解析が進展することにより,初期の脊椎動物が対鼻孔や顎を獲得した進化機構が明らかになることが期待される.
文 献
- Kuraku, S.: Insights into cyclostome phylogenomics: pre-2R or post-2R. Zool. Sci., 25, 960-968 (2008)[PubMed]
- Heimberg, A. M., Cowper-Sal-lari, R., Peterson, K. J. et al.: microRNAs reveal the interrelationships of hagfish, lampreys, and gnathostomes and the nature of the ancestral vertebrate. Proc. Natl. Acad. Sci. USA, 107, 19379-19383 (2010)[PubMed]
- Janvier, P.: Early Vertebrates. Oxford University Press, New York (1996)
- Gess, R. W., Coates, M. I. & Rubidge, B. S.: A lamprey from the Devonian period of South Africa. Nature, 443, 981-984 (2006)[PubMed]
- Gorbman, A.: Early development of the hagfish pituitary gland: evidence for the endodermal origin of the adenohypophysis. Am. Zool., 23, 639-654 (1983)
- Soukup, V., Horacek, I. & Cerny, R.: Development and evolution of the vertebrate primary mouth. J. Anat., 222, 79-99 (2013)[PubMed]
- Ota, K. G., Kuraku, S. & Kuratani, S.: Hagfish embryology with reference to the evolution of the neural crest. Nature, 446, 672-675 (2007)[PubMed]
- Ota, K. G., Fujimoto, S., Kuratani, S. et al.: Identification of vertebra-like elements and their possible differentiation from sclerotomes in the hagfish. Nat. Commun., 2, 373 (2011)[PubMed]
- Kuratani, S.: Evolution of the vertebrate jaw from developmental perspectives. Evol. Dev., 14, 76-92 (2012)[PubMed]
- Gai, Z., Donoghue, P. C., Stampanoni, M. et al.: Fossil jawless fish from China foreshadows early jawed vertebrate anatomy. Nature, 476, 324-327 (2011)[PubMed]
著者プロフィール
略歴:神戸大学大学院理学研究科博士課程 在学中.
研究テーマ:動物の形態要素,構造からみた発生と進化.
倉谷 滋(Shigeru Kuratani)
理化学研究所発生再生科学総合研究センター グループディレクター.
研究室URL:http://www.cdb.riken.jp/emo/japanese/indexj.html
© 2013 大石康博・倉谷 滋 Licensed under CC 表示 2.1 日本
