分子モータータンパク質KIF5AはGABAA受容体のシナプスへの輸送に必須でありその欠損はてんかんをひき起こす
武井陽介・廣川信隆
(東京大学大学院医学系研究科 分子構造・動態学講座)
email:廣川信隆
DOI: 10.7875/first.author.2013.005
Molecular motor KIF5A is essential for GABAA receptor transport, and KIF5A deletion causes epilepsy.
Kazuo Nakajima, Xiling Yin, Yosuke Takei, Dae-Hyun Seog, Noriko Homma, Nobutaka Hirokawa
Neuron, 76, 945-961 (2012)
KIF5Aはキネシンスーパーファミリータンパク質に属する分子モータータンパク質であり,ニューロンにおいて多量に発現していることが知られていたが,その特異的な機能は不明であった.今回,筆者らが,KIF5Aノックアウトマウスを作製し観察したところ,生後数週間でてんかんを発症した.KIF5AノックアウトマウスのニューロンにおいてはGABAA受容体の輸送が低下しており,そのため,抑制性シナプスの後部におけるGABAA受容体の量の減少と抑制性神経伝達の低下が起こっていた.この原因を調べるためKIF5A結合タンパク質をスクリーニングしたところ,GABAA受容体結合タンパク質であるGABARAPがKIF5AのC末端側領域と結合することがわかった.以上より,GABAA受容体はKIF5AとGABARAPとの複合体によりシナプスへ輸送されており,この輸送が破綻すると抑制性神経伝達と興奮性神経伝達とのバランスがくずれ,てんかんの発症にいたるものと考えられた.
キネシンスーパーファミリータンパク質(kinesin superfamily protein:KIF)は細胞の内部において微小管にそって膜オルガネラ,タンパク質複合体,mRNAなど種々の荷物(カーゴ)を輸送する分子モータータンパク質であり,細胞分化,形態形成,機能制御などさまざまな局面において重要な役割をはたしている1,2).とりわけ,ニューロンにおいてはこれらのタンパク質がさまざまな神経疾患や記憶障害に関与していることが示されてきた3-5).今回,筆者らは,キネシンスーパーファミリータンパク質のなかで神経系に豊富に存在するKIF5Aに注目した.KIF5Aは類似したタンパク質KIF5BおよびKIF5CとともにKIF5サブファミリーを形成するが6),KIF5Aだけに特異的なカーゴおよび機能は不明であり,くわしい解明が待たれていた.KIF5Aを機能面から理解するためKIF5Aノックアウトマウスを作製し解析したところ,てんかん症状が観察された.
てんかんとは,大脳におけるニューロンの過剰な活動の結果,発作性の神経症状をくり返す慢性の脳障害である.もっとも頻度の高い精神神経疾患のひとつであり,世界で5000万人に及ぶ患者がてんかんの発作と発作が原因で起こるさまざまな生活障害に苦しんでいる7).てんかんの原因の解明はまだ不十分であり,さまざまな薬物療法や外科手術などが行われているが根本治療は確立されていない.今回,筆者らは,KIF5Aノックアウトマウスのニューロンの細胞生物学的な解析とKIF5A結合タンパク質およびKIF5Aのカーゴの同定により,KIF5AによるGABAA受容体の輸送の破綻によりてんかんがひき起こされることを明らかにした.
胎生期からKIF5Aを欠失させると生後すぐにそのマウスは死んでしまい,以後の解析ができない.そこで,生後1~2週間をへてKIF5Aの欠失を開始するノックアウトマウスを作製した.このマウスは生後数週間するとしばしば発作的に転倒および無動のエピソードをくり返し,成長の遅れがめだつようになり,生後4週齢までにすべて死亡した.KIF5Aノックアウトマウスの海馬に電極を刺入して脳波を記録したところ,棘波や棘徐波結合など突発性の異常がくり返し出現し,てんかんであることが判明した.KIF5Aノックアウトマウスの脳波を周波数解析したところ,すべての周波数帯域において脳波の振幅は減少しており,脳波のベースラインの異常も明らかになった.
神経伝達には興奮性と抑制性の2種類があり,抑制性神経伝達の障害がてんかんの原因であることが知られている8).KIF5Aノックアウトマウスの抑制性神経伝達について調べるため,海馬急速切片を作製しパッチクランプ法によりKIF5Aノックアウトマウスと野生型マウスとで比較した.その結果,KIF5Aノックアウトマウスのニューロンでは微小な抑制性後シナプス電流の振幅が野生型マウスの半分程度に減少しており,電気刺激に対する抑制性後シナプス電流の応答も減弱していることが明らかになった.以上の結果から,KIF5Aノックアウトマウスの脳には抑制性シナプスの機能障害があり,その結果,てんかんをひき起こしているものと考えられた.
脳における抑制性神経伝達をつかさどる主要な受容体であるGABAA受容体について検討した.GABAA受容体はγ-アミノ酪酸(GABA)を神経伝達物質とするイオンチャネル内蔵型の受容体である8).KIF5Aノックアウトマウスのニューロンにおいて,GABAA受容体の総量は変わらなかったが,細胞表面に発現するGABAA受容体は著しく減少しており,より多くのGABAA受容体は細胞の内部にとどまっていた.免疫染色法により樹状突起におけるGABAA受容体の減少が確認された.KIF5A,KIF5B,KIF5Cのそれぞれをコードする遺伝子をKIF5Aノックアウトマウスのニューロンに導入することでレスキュー実験を行ったところ,KIF5Aを発現した場合のみGABAA受容体の細胞表面における発現が回復したため,KIF5Aだけが特異的にGABAA受容体を細胞表面へと輸送していることが示唆された.
KIF5AとGABAA受容体がどのように分子的な関連をもつのか調べるため,酵母ツーハイブリッド法を用いてKIF5Aに特異的なC末端領域に結合するタンパク質をスクリーニングしたところ,GABAA受容体結合タンパク質であるGABARAP(GABAA receptor-associated protein)が同定された.KIF5AとGABARAPとの結合は組換えタンパク質どうしの直接結合により確認された.このKIF5AとGABARAPとの結合と,GABARAPがGABAA受容体のγサブユニットに結合すること9) をあわせると,細胞においてKIF5A-GABARAP-GABAA受容体という複合体が形成されているものと考えられた.免疫沈降法および二重免疫染色法によりKIF5AとGABARAPとの細胞における結合が確認された.
KIF5Aノックアウトマウスのニューロンでは樹状突起におけるGABARAPの局在が変化し,突起の末端においてGABARAPが減少していた.miRNAノックダウン法によりGABARAPの発現を低下させると,GABAA受容体の細胞表面およびシナプスにおける発現が低下し,また,KIF5AのGABARAP結合ドメイン断片をニューロンに発現すると,ドミナントネガティブ効果により細胞表面のGABAA受容体の量が低下した.これらの結果から,KIF5AとGABARAPとの複合体はGABAA受容体の細胞表面への輸送に必須であると考えられた.
GFPで標識したGABAA受容体をニューロンに発現させ,生きたニューロンの内部においてGABAA受容体の樹状突起における輸送を可視化しシグナルの動態を解析した.KIF5AノックアウトマウスのニューロンおよびGABARAPをノックダウンしたニューロンにおいて,GABAA受容体の輸送は著明に低下していた.さらに,GFP標識GABAA受容体を発現させたニューロンを,小胞体からゴルジ体へのタンパク質輸送の特異的な阻害剤であるブレフェルディンAにより処理し,つづいてブレフェルディンAを除きGFP標識GABAA受容体の動態をみることにより蛍光シグナルが小胞体からゴルジ体をへて樹状突起の内部を移動するようすを観察したところ,KIF5Aノックアウトマウスのニューロンではゴルジ体から放出され樹状突起の末端へと輸送される過程が著しく遅れていることが判明した.これらの結果は,KIF5AとGABARAPとの複合体がGABAA受容体の細胞表面およびシナプスへの輸送に重要であることを示した.
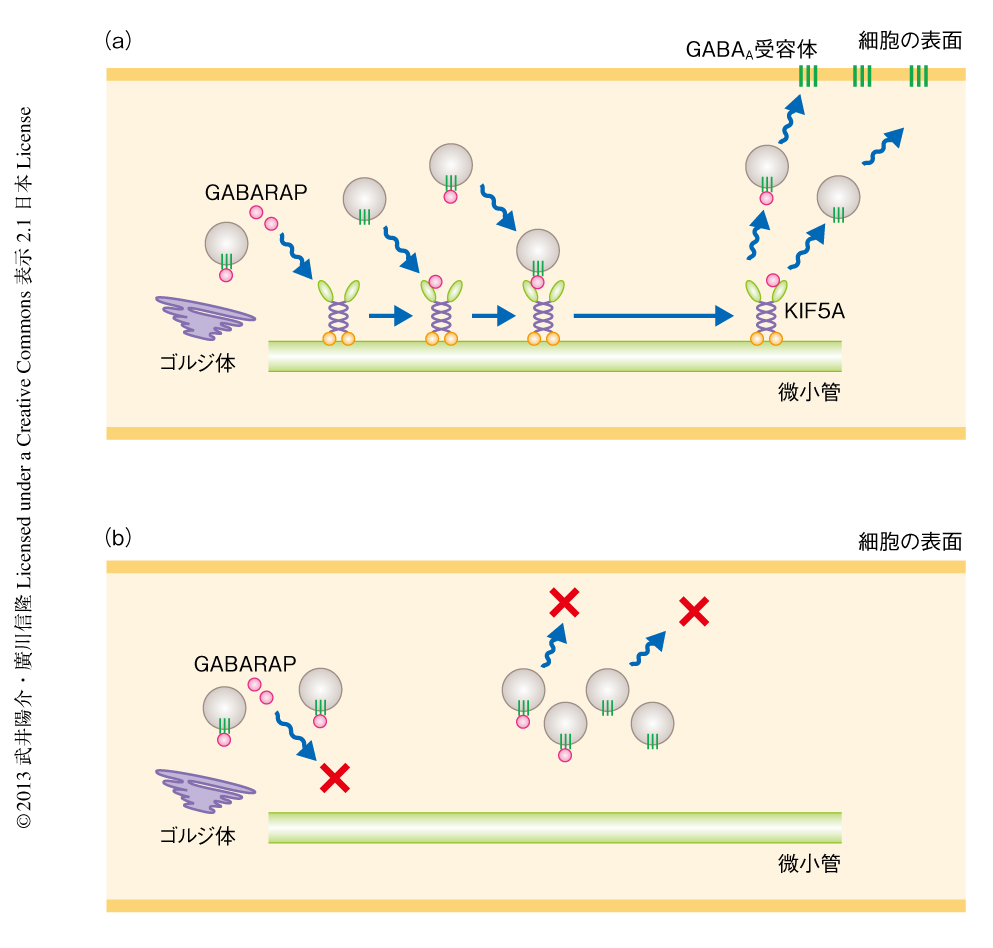
この研究において,筆者らは,KIF5Aの結合タンパク質としてGABAA受容体結合タンパク質GABARAPを同定した.KIF5Aの一次構造はKIF5BおよびKIF5Cときわめて類似しており6),これまで,これらKIF5サブファミリーのカーゴはすべて同一だと考えられていた.ところが,KIF5Aに特異的な結合タンパク質の発見により,KIF5AがGABARAPをなかだちとする独自の輸送系(図1a)によりGABAA受容体を輸送していることが判明した.KIF5Aノックアウトマウスの脳ではGABAA受容体が正しくシナプスへと輸送されないため抑制性神経伝達の障害が起こる(図1b).神経伝達には興奮性と抑制性と2種類があり,抑制性神経伝達が低下することで両者のバランスが興奮側へとかたむき,てんかんの発症につながるものと考えられた.
この研究は,細胞におけるGABAA受容体の輸送の破綻によるてんかんの発症という新しい機序を明らかにした.今後,さらにこの輸送系の制御機構における詳細を明らかにし,将来的には,てんかんの予防や治療法の開発にむけたアプローチへとつなげていきたい.
略歴:東京大学大学院医学系研究科 准教授.
研究テーマ:神経系の細胞骨格とその関連タンパク質.
廣川 信隆(Nobutaka Hirokawa)
東京大学大学院医学系研究科 特任教授.
© 2013 武井陽介・廣川信隆 Licensed under CC 表示 2.1 日本
(東京大学大学院医学系研究科 分子構造・動態学講座)
email:廣川信隆
DOI: 10.7875/first.author.2013.005
Molecular motor KIF5A is essential for GABAA receptor transport, and KIF5A deletion causes epilepsy.
Kazuo Nakajima, Xiling Yin, Yosuke Takei, Dae-Hyun Seog, Noriko Homma, Nobutaka Hirokawa
Neuron, 76, 945-961 (2012)
要 約
KIF5Aはキネシンスーパーファミリータンパク質に属する分子モータータンパク質であり,ニューロンにおいて多量に発現していることが知られていたが,その特異的な機能は不明であった.今回,筆者らが,KIF5Aノックアウトマウスを作製し観察したところ,生後数週間でてんかんを発症した.KIF5AノックアウトマウスのニューロンにおいてはGABAA受容体の輸送が低下しており,そのため,抑制性シナプスの後部におけるGABAA受容体の量の減少と抑制性神経伝達の低下が起こっていた.この原因を調べるためKIF5A結合タンパク質をスクリーニングしたところ,GABAA受容体結合タンパク質であるGABARAPがKIF5AのC末端側領域と結合することがわかった.以上より,GABAA受容体はKIF5AとGABARAPとの複合体によりシナプスへ輸送されており,この輸送が破綻すると抑制性神経伝達と興奮性神経伝達とのバランスがくずれ,てんかんの発症にいたるものと考えられた.
はじめに
キネシンスーパーファミリータンパク質(kinesin superfamily protein:KIF)は細胞の内部において微小管にそって膜オルガネラ,タンパク質複合体,mRNAなど種々の荷物(カーゴ)を輸送する分子モータータンパク質であり,細胞分化,形態形成,機能制御などさまざまな局面において重要な役割をはたしている1,2).とりわけ,ニューロンにおいてはこれらのタンパク質がさまざまな神経疾患や記憶障害に関与していることが示されてきた3-5).今回,筆者らは,キネシンスーパーファミリータンパク質のなかで神経系に豊富に存在するKIF5Aに注目した.KIF5Aは類似したタンパク質KIF5BおよびKIF5CとともにKIF5サブファミリーを形成するが6),KIF5Aだけに特異的なカーゴおよび機能は不明であり,くわしい解明が待たれていた.KIF5Aを機能面から理解するためKIF5Aノックアウトマウスを作製し解析したところ,てんかん症状が観察された.
てんかんとは,大脳におけるニューロンの過剰な活動の結果,発作性の神経症状をくり返す慢性の脳障害である.もっとも頻度の高い精神神経疾患のひとつであり,世界で5000万人に及ぶ患者がてんかんの発作と発作が原因で起こるさまざまな生活障害に苦しんでいる7).てんかんの原因の解明はまだ不十分であり,さまざまな薬物療法や外科手術などが行われているが根本治療は確立されていない.今回,筆者らは,KIF5Aノックアウトマウスのニューロンの細胞生物学的な解析とKIF5A結合タンパク質およびKIF5Aのカーゴの同定により,KIF5AによるGABAA受容体の輸送の破綻によりてんかんがひき起こされることを明らかにした.
1.KIF5Aノックアウトマウスはてんかんを発症する
胎生期からKIF5Aを欠失させると生後すぐにそのマウスは死んでしまい,以後の解析ができない.そこで,生後1~2週間をへてKIF5Aの欠失を開始するノックアウトマウスを作製した.このマウスは生後数週間するとしばしば発作的に転倒および無動のエピソードをくり返し,成長の遅れがめだつようになり,生後4週齢までにすべて死亡した.KIF5Aノックアウトマウスの海馬に電極を刺入して脳波を記録したところ,棘波や棘徐波結合など突発性の異常がくり返し出現し,てんかんであることが判明した.KIF5Aノックアウトマウスの脳波を周波数解析したところ,すべての周波数帯域において脳波の振幅は減少しており,脳波のベースラインの異常も明らかになった.
2.KIF5Aノックアウトマウスのニューロンにおける抑制性神経伝達の低下
神経伝達には興奮性と抑制性の2種類があり,抑制性神経伝達の障害がてんかんの原因であることが知られている8).KIF5Aノックアウトマウスの抑制性神経伝達について調べるため,海馬急速切片を作製しパッチクランプ法によりKIF5Aノックアウトマウスと野生型マウスとで比較した.その結果,KIF5Aノックアウトマウスのニューロンでは微小な抑制性後シナプス電流の振幅が野生型マウスの半分程度に減少しており,電気刺激に対する抑制性後シナプス電流の応答も減弱していることが明らかになった.以上の結果から,KIF5Aノックアウトマウスの脳には抑制性シナプスの機能障害があり,その結果,てんかんをひき起こしているものと考えられた.
3.KIF5AノックアウトマウスのニューロンではGABAA受容体の細胞表面における発現が低下している
脳における抑制性神経伝達をつかさどる主要な受容体であるGABAA受容体について検討した.GABAA受容体はγ-アミノ酪酸(GABA)を神経伝達物質とするイオンチャネル内蔵型の受容体である8).KIF5Aノックアウトマウスのニューロンにおいて,GABAA受容体の総量は変わらなかったが,細胞表面に発現するGABAA受容体は著しく減少しており,より多くのGABAA受容体は細胞の内部にとどまっていた.免疫染色法により樹状突起におけるGABAA受容体の減少が確認された.KIF5A,KIF5B,KIF5Cのそれぞれをコードする遺伝子をKIF5Aノックアウトマウスのニューロンに導入することでレスキュー実験を行ったところ,KIF5Aを発現した場合のみGABAA受容体の細胞表面における発現が回復したため,KIF5Aだけが特異的にGABAA受容体を細胞表面へと輸送していることが示唆された.
4.GABAA受容体結合タンパク質GABARAPはKIF5AのC末端領域に結合する
KIF5AとGABAA受容体がどのように分子的な関連をもつのか調べるため,酵母ツーハイブリッド法を用いてKIF5Aに特異的なC末端領域に結合するタンパク質をスクリーニングしたところ,GABAA受容体結合タンパク質であるGABARAP(GABAA receptor-associated protein)が同定された.KIF5AとGABARAPとの結合は組換えタンパク質どうしの直接結合により確認された.このKIF5AとGABARAPとの結合と,GABARAPがGABAA受容体のγサブユニットに結合すること9) をあわせると,細胞においてKIF5A-GABARAP-GABAA受容体という複合体が形成されているものと考えられた.免疫沈降法および二重免疫染色法によりKIF5AとGABARAPとの細胞における結合が確認された.
5.KIF5AとGABARAPとの複合体はGABAA受容体の輸送に重要である
KIF5Aノックアウトマウスのニューロンでは樹状突起におけるGABARAPの局在が変化し,突起の末端においてGABARAPが減少していた.miRNAノックダウン法によりGABARAPの発現を低下させると,GABAA受容体の細胞表面およびシナプスにおける発現が低下し,また,KIF5AのGABARAP結合ドメイン断片をニューロンに発現すると,ドミナントネガティブ効果により細胞表面のGABAA受容体の量が低下した.これらの結果から,KIF5AとGABARAPとの複合体はGABAA受容体の細胞表面への輸送に必須であると考えられた.
GFPで標識したGABAA受容体をニューロンに発現させ,生きたニューロンの内部においてGABAA受容体の樹状突起における輸送を可視化しシグナルの動態を解析した.KIF5AノックアウトマウスのニューロンおよびGABARAPをノックダウンしたニューロンにおいて,GABAA受容体の輸送は著明に低下していた.さらに,GFP標識GABAA受容体を発現させたニューロンを,小胞体からゴルジ体へのタンパク質輸送の特異的な阻害剤であるブレフェルディンAにより処理し,つづいてブレフェルディンAを除きGFP標識GABAA受容体の動態をみることにより蛍光シグナルが小胞体からゴルジ体をへて樹状突起の内部を移動するようすを観察したところ,KIF5Aノックアウトマウスのニューロンではゴルジ体から放出され樹状突起の末端へと輸送される過程が著しく遅れていることが判明した.これらの結果は,KIF5AとGABARAPとの複合体がGABAA受容体の細胞表面およびシナプスへの輸送に重要であることを示した.
おわりに
この研究において,筆者らは,KIF5Aの結合タンパク質としてGABAA受容体結合タンパク質GABARAPを同定した.KIF5Aの一次構造はKIF5BおよびKIF5Cときわめて類似しており6),これまで,これらKIF5サブファミリーのカーゴはすべて同一だと考えられていた.ところが,KIF5Aに特異的な結合タンパク質の発見により,KIF5AがGABARAPをなかだちとする独自の輸送系(図1a)によりGABAA受容体を輸送していることが判明した.KIF5Aノックアウトマウスの脳ではGABAA受容体が正しくシナプスへと輸送されないため抑制性神経伝達の障害が起こる(図1b).神経伝達には興奮性と抑制性と2種類があり,抑制性神経伝達が低下することで両者のバランスが興奮側へとかたむき,てんかんの発症につながるものと考えられた.
この研究は,細胞におけるGABAA受容体の輸送の破綻によるてんかんの発症という新しい機序を明らかにした.今後,さらにこの輸送系の制御機構における詳細を明らかにし,将来的には,てんかんの予防や治療法の開発にむけたアプローチへとつなげていきたい.
文 献
- Hirokawa, N., Noda Y., Tanaka, Y. et al.: Kinesin superfamily motor proteins and intracellular transport. Nat. Rev. Mol. Cell Biol., 10, 682-696 (2009)[PubMed]
- Hirokawa, N., Niwa, S. & Tanaka, Y.: Molecular motors in neurons: transport mechanisms and roles in brain function, development, and disease. Neuron, 68, 610-638 (2010)[PubMed]
- Yin, X., Takei, Y., Kido, M. A. et al.: Molecular motor KIF17 is fundamental for memory and learning via differential support of synaptic NR2A/2B levels. Neuron, 70, 310-325 (2011)[PubMed]
- Yin, X., Feng, X., Takei, Y. et al.: Regulation of NMDA receptor transport: a KIF17-cargo binding/releasing underlies synaptic plasticity and memory in vivo. J. Neurosci., 32, 5486-5499 (2012)[PubMed]
- Kondo, M., Takei, Y. & Hirokawa, N.: Motor protein KIF1A is essential for hippocampal synaptogenesis and learning enhancement in an enriched environment. Neuron, 73, 743-757 (2012)[PubMed] [新着論文レビュー]
- Kanai, Y., Okada, Y., Tanaka, Y. et al.: KIF5C, a novel neuronal kinesin enriched in motor neurons. J. Neurosci., 20, 6374-6384 (2000)[PubMed]
- Rakhade, S. N. & Jensen, F. E.: Epileptogenesis in the immature brain: emerging mechanisms. Nat. Rev. Neurol., 5, 380-391 (2009)[PubMed]
- Jacob, T. C., Moss, S. J. & Jurd, R.: GABAA receptor trafficking and its role in the dynamic modulation of neuronal inhibition. Nat. Rev. Neurosci., 9, 331-343 (2008)[PubMed]
- Wang, H., Bedford, F. K., Brandon, N. J. et al.: GABAA-receptor-associated protein links GABAA receptors and the cytoskeleton. Nature, 397, 69-72 (1999)[PubMed]
著者プロフィール
略歴:東京大学大学院医学系研究科 准教授.
研究テーマ:神経系の細胞骨格とその関連タンパク質.
廣川 信隆(Nobutaka Hirokawa)
東京大学大学院医学系研究科 特任教授.
© 2013 武井陽介・廣川信隆 Licensed under CC 表示 2.1 日本
