フルクトース受容体は脳において栄養センサーとして機能する
宮本徹也・Hubert Amrein
(米国Texas A&M Health Science Center,Department of Molecular and Cellular Medicine)
email:宮本徹也
DOI: 10.7875/first.author.2012.154
A fructose receptor functions as a nutrient sensor in the Drosophila brain.
Tetsuya Miyamoto, Jesse Slone, Xiangyu Song, Hubert Amrein
Cell, 151, 1113-1125 (2012)
食物の栄養価は摂食行動に大きな影響を及ぼすが,その感知機構は未解明である.筆者らは,Ca2+イメージングと行動実験により,ショウジョウバエの味覚受容体Gr43aはフルクトース受容体であり,味覚器官にくわえ脳においても機能していることを明らかにした.体内のフルクトースのレベルは栄養価のある糖を摂取すると著しく上昇することから,摂食実験の結果とあわせて,脳に存在するGr43aは栄養センサーとしてはたらいていることが示された.興味深いことに,その機能は体内の栄養状態に依存しており,空腹状態においては摂食行動を促進するが,満腹状態では逆にこれを抑制していた.古典的条件付けによりGr43aを発現するニューロンのひき起こす知覚について調べたところ,実際に空腹状態あるいは満腹状態に応じ快から不快へと正反対に変化することが明らかになった.これらの結果から,Gr43aは血中のフルクトースを介した栄養センサーであり,知覚の形成をつうじ摂食行動を制御していることが示された.
動物のゲノムには約100種類もの味覚受容体の遺伝子が存在し,それぞれの受容体は化学物質に対し独自の結合特性をもつ1).その多くは味覚器官でのみはたらいているが,少数は脳や腸においても発現がみられることから,味覚分子が体内で感知されていることを示している2,3).また,最近の研究から,動物は味覚にたよらず栄養価のみを指標に食物を識別できることが明らかになり,体内に栄養センサーをもつことが提唱されていた4-6).しかし,その分子実体や機能している器官,感知する栄養分子についてはまったく不明であった.筆者らは,ショウジョウバエの味覚受容体Gr43aが栄養センサーとして機能していることを明らかにし,その作動機構の解析を行った.
ショウジョウバエの味覚受容体ファミリー遺伝子は68個の遺伝子からなるが,機能の明らかになったものはわずか10個程度である.そこで,味覚受容体がはたす未知の役割を発見するため,味覚受容体ファミリー遺伝子のなかでもっとも保存されたもののひとつ,Gr43a遺伝子の解析を行った.まず,その発現部位を調べたところ,口器および脚の味覚器官にくわえ,脳,腸,子宮といった内部器官でも発現しており,さまざまな生体プロセスにかかわっていることが示唆された.
味覚受容体がその認識する分子と結合すると,細胞質へのCa2+の流入をひき起こしニューロンを興奮させる.そこで,さまざまな物質をあたえたときの,Gr43aを発現するニューロンの内部におけるCa2+濃度の変化を測定した.その結果,脚および脳においてGr43aはフルクトース受容体として機能していることが明らかになった.これはほかの実験系による結果とよく一致しており7),昆虫は体外だけではなく体内のフルクトースをも感知していることが示された.興味深いことに,脳においてGr43aを発現するニューロンは昆虫におけるおもな血糖であるグルコースやその二糖であるトレハロースにはまったく反応しなかった.
フルクトースは代表的な単糖のひとつである.同じ単糖のグルコースとともに野菜や果物に豊富に含まれており,動物の重要なエネルギー源となっている.しかし,体内においてはまったく状況が異なり,グルコースがエネルギー代謝の基本分子として高濃度に蓄積されているのに対し,フルクトースはそのわずか1%程度しか存在しない.そのため,これまでその役割が注目されることはほとんどなく,フルクトースに特異的に反応するニューロンが脳に存在していることは驚きであった.
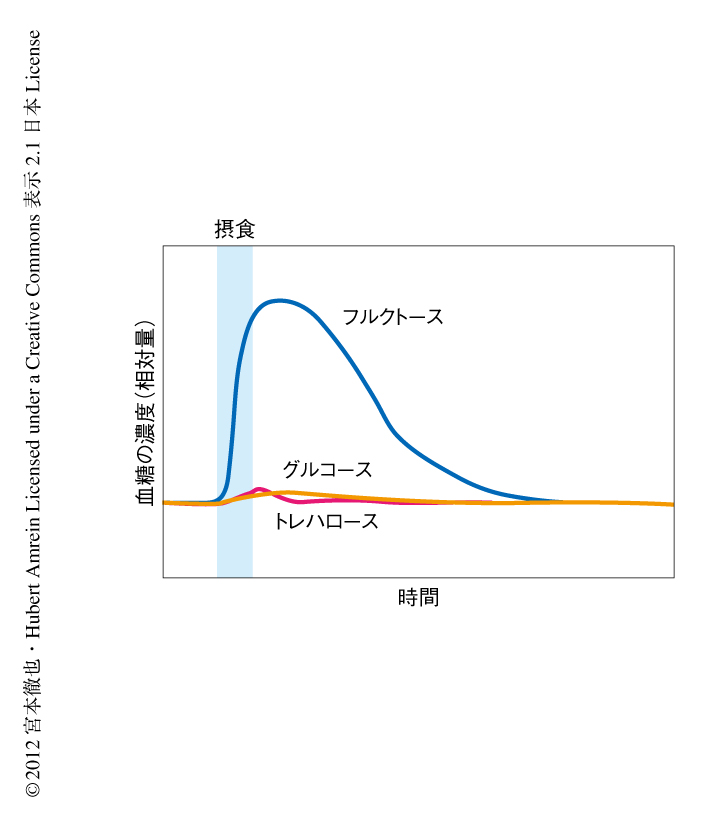
血中のフルクトースの機能を明らかにするため,その代謝ダイナミクスを調べたところ,摂食の直後にフルクトース濃度は著しく上昇することがわかった(図1).興味深いことに,この現象はフルクトースを摂取したときにかぎらず,グルコースなどほかの糖を摂取したときも同様であった.これは,過剰な糖分の一部が代謝によりフルクトースへと変換されたためと考えられた.それに対し,甘味料として使われるシュークラロースなど代謝されない糖を摂取してもフルクトース濃度に変化はなかった.また,おもな血糖であるグルコースやトレハロースは体内に大量に蓄えられているため,摂食ののちの相対的な変化はきわめて小さかった(図1).これらのことから,血中のフルクトース濃度は栄養価のある糖の摂取を反映し,きわめて大きく変動することが明らかになった.
最近の研究から,ショウジョウバエは糖類の一種ソルビトールを識別できることが明らかになっていた5,6).ソルビトールには栄養価があるが,ショウジョウバエはその味を感じることができないため,この結果は栄養センサーの存在を示唆するものであった.さきに述べた血中のフルクトースの代謝ダイナミクスの知見から,フルクトース受容体であるGr43aは栄養センサーの候補であると考えられた.それを確かめるため,水とソルビトール溶液とを同時にあたえてその摂取量を比較したところ,野生型ショウジョウバエは水よりもソルビトールを好んだが,Gr43a遺伝子に変異をもつショウジョウバエはソルビトールへの嗜好性をまったく示さなかった.このことから,Gr43aは実際に栄養センサーとして機能しており,ショウジョウバエは食物中の栄養価を血中のフルクトース濃度をつうじ感知していることが明らかになった.
さきに述べた摂食実験では,Gr43a変異ショウジョウバエのソルビトールの摂取量は野生型ショウジョウバエよりも少なかった.ところが,甘くて栄養価のあるシュークロースやグルコースをあたえた場合,Gr43a変異体における摂取量は逆に野生型より数十%も多かった.シュークロースの摂取量は味のないソルビトールの約3倍にもなったことから,Gr43aを発現するニューロンの作用は空腹状態あるいは満腹状態に応じ正反対になると考えられた.それを明確に示すため,甘いが栄養価のないシュークラロースをソルビトールに混ぜ,栄養価に対し影響をあたえることなくソルビトールの摂取量を増加させた.すると,野生型では摂取量が2倍程度になったが,Gr43a変異体では3倍以上に増加し野生型よりも著しく多くなった.このことから,Gr43aを発現するニューロンは空腹状態では摂食行動を促進するが,満腹状態では逆に抑制的にはたらくことが明らかになった.同一のニューロンが正反対の役割をはたすのはきわめて例外的である.
Gr43aは栄養センサーとして摂食行動をどのように制御しているのだろうか.Gr43aを発現するニューロンは食欲に直接にはたらきかけているのではなく,摂取した食物に対する知覚をつうじ間接的に影響を及ぼしているという仮説をたてた.それを確かめるため,パブロフの犬で有名な古典的条件付けを利用して,Gr43aを発現するニューロンが知覚を生じさせる可能性を検討した.そのために,まず,Gr43aを発現するニューロンを強制的に興奮させ,同時に特定の匂いをかがせることにより,その経験と匂いを関連づけて記憶させた.もし,Gr43aを発現するニューロンの興奮が知覚を生じさせるならば,それに応じ,ショウジョウバエはその匂いに対してなんらかの反応(接近または回避)を示すはずである.実験の結果,空腹状態ではGr43aを発現するニューロンの興奮は実際に正の知覚をひき起こしていることが明らかになった.重要なことに,満腹状態においてその興奮は逆に負にとらえられていた.
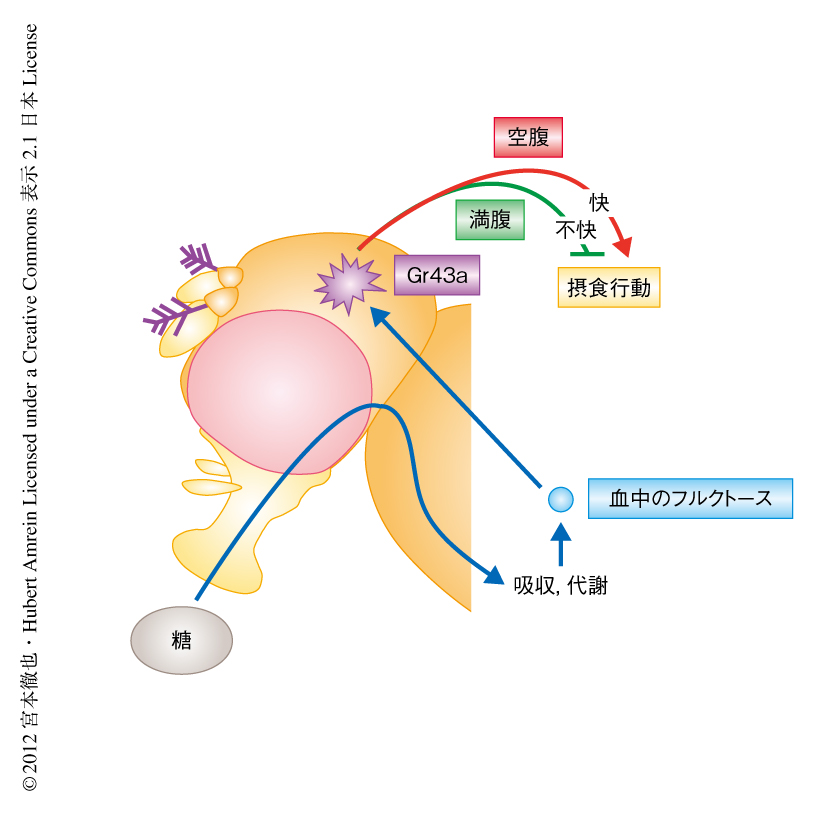
以上の結果は,栄養センサーは空腹状態では摂食行動を促進し,満腹状態では摂食行動を抑制することと完全に一致していた(図2).すなわち,栄養価のある食物を摂取するとGr43aを発現するニューロンが興奮し,空腹の場合にはそれを正に知覚するため,その食物の摂取に対し肯定的な評価がくだされる.すると,その行動をくり返し,結果的に栄養価のある食物の摂取量のみが増加することになる.反対に,満腹のときには栄養価のある食物を摂取すると負の知覚が生じるため,栄養価のある食物の摂取量は減少する.これらのことから,Gr43aを発現するニューロンは食物の摂取に対し栄養状態に応じた評価をあたえることにより,飽食あるいは飢餓を防いでいることが明らかになった.
グルコース(および,昆虫ではその二糖であるトレハロース)はエネルギー代謝の基本分子であり,体内につねに高濃度で維持されているため,食物の摂取による変化はきわめて小さい.この研究から明らかになった栄養価の感知機構は,その問題を見事に解決していた.この機構のもっとも大きな長所は,栄養分子として用いるフルクトースは食物には豊富に含まれているが体内にはほとんど存在しないため,わずかな食事であってもその血中の濃度が大きく変動することである.それにくわえて,フルクトースはグルコースから容易に変換されうるため,原理的には栄養価のある(グルコースに変換されうる)すべての分子の検出が可能である.
Gr43a遺伝子は昆虫の種間でもっとも保存された味覚受容体の遺伝子のひとつであり,そのことはこの機構が昆虫にとりきわめて重要であることを示している.それでは,ヒトにも栄養センサーは存在しているのだろうか.味覚受容体ファミリー遺伝子は脊椎動物と無脊椎動物とで独自に進化したため,ヒトにはGr43a遺伝子は存在しない.しかしながら,血中におけるグルコースとフルクトースとの量比はヒトでもショウジョウバエと同じ100対1程度であり,マウスにおいては食物の栄養価を感知する能力も示されている4).栄養センサーの存在の直接の証拠は得られていないが,今後の研究が待たれるところである.
略歴:2004年 名古屋大学大学院理学研究科 修了,同年 名古屋大学遺伝子実験施設 研究員,2005年 米国Duke大学Postdoctoral fellow,2009年 米国Texas A&M Health Science Center Postdoctoral fellowを経て,2010年より同Postdoctoral Research Associate.
研究テーマ:ショウジョウバエの味覚.
抱負:個別の現象をとおして脳の機能の本質を探っていきたい.
Hubert Amrein
米国Texas A&M Health Science CenterにてProfessor.
研究室URL:http://amreinlab.wordpress.com/
© 2012 宮本徹也・Hubert Amrein Licensed under CC 表示 2.1 日本
(米国Texas A&M Health Science Center,Department of Molecular and Cellular Medicine)
email:宮本徹也
DOI: 10.7875/first.author.2012.154
A fructose receptor functions as a nutrient sensor in the Drosophila brain.
Tetsuya Miyamoto, Jesse Slone, Xiangyu Song, Hubert Amrein
Cell, 151, 1113-1125 (2012)
要 約
食物の栄養価は摂食行動に大きな影響を及ぼすが,その感知機構は未解明である.筆者らは,Ca2+イメージングと行動実験により,ショウジョウバエの味覚受容体Gr43aはフルクトース受容体であり,味覚器官にくわえ脳においても機能していることを明らかにした.体内のフルクトースのレベルは栄養価のある糖を摂取すると著しく上昇することから,摂食実験の結果とあわせて,脳に存在するGr43aは栄養センサーとしてはたらいていることが示された.興味深いことに,その機能は体内の栄養状態に依存しており,空腹状態においては摂食行動を促進するが,満腹状態では逆にこれを抑制していた.古典的条件付けによりGr43aを発現するニューロンのひき起こす知覚について調べたところ,実際に空腹状態あるいは満腹状態に応じ快から不快へと正反対に変化することが明らかになった.これらの結果から,Gr43aは血中のフルクトースを介した栄養センサーであり,知覚の形成をつうじ摂食行動を制御していることが示された.
はじめに
動物のゲノムには約100種類もの味覚受容体の遺伝子が存在し,それぞれの受容体は化学物質に対し独自の結合特性をもつ1).その多くは味覚器官でのみはたらいているが,少数は脳や腸においても発現がみられることから,味覚分子が体内で感知されていることを示している2,3).また,最近の研究から,動物は味覚にたよらず栄養価のみを指標に食物を識別できることが明らかになり,体内に栄養センサーをもつことが提唱されていた4-6).しかし,その分子実体や機能している器官,感知する栄養分子についてはまったく不明であった.筆者らは,ショウジョウバエの味覚受容体Gr43aが栄養センサーとして機能していることを明らかにし,その作動機構の解析を行った.
1.味覚受容体Gr43aは味覚器官にくわえ体内でも発現している
ショウジョウバエの味覚受容体ファミリー遺伝子は68個の遺伝子からなるが,機能の明らかになったものはわずか10個程度である.そこで,味覚受容体がはたす未知の役割を発見するため,味覚受容体ファミリー遺伝子のなかでもっとも保存されたもののひとつ,Gr43a遺伝子の解析を行った.まず,その発現部位を調べたところ,口器および脚の味覚器官にくわえ,脳,腸,子宮といった内部器官でも発現しており,さまざまな生体プロセスにかかわっていることが示唆された.
2.Gr43aはフルクトース受容体である
味覚受容体がその認識する分子と結合すると,細胞質へのCa2+の流入をひき起こしニューロンを興奮させる.そこで,さまざまな物質をあたえたときの,Gr43aを発現するニューロンの内部におけるCa2+濃度の変化を測定した.その結果,脚および脳においてGr43aはフルクトース受容体として機能していることが明らかになった.これはほかの実験系による結果とよく一致しており7),昆虫は体外だけではなく体内のフルクトースをも感知していることが示された.興味深いことに,脳においてGr43aを発現するニューロンは昆虫におけるおもな血糖であるグルコースやその二糖であるトレハロースにはまったく反応しなかった.
3.血中のフルクトースは摂取した食物の栄養価を反映する
フルクトースは代表的な単糖のひとつである.同じ単糖のグルコースとともに野菜や果物に豊富に含まれており,動物の重要なエネルギー源となっている.しかし,体内においてはまったく状況が異なり,グルコースがエネルギー代謝の基本分子として高濃度に蓄積されているのに対し,フルクトースはそのわずか1%程度しか存在しない.そのため,これまでその役割が注目されることはほとんどなく,フルクトースに特異的に反応するニューロンが脳に存在していることは驚きであった.
血中のフルクトースの機能を明らかにするため,その代謝ダイナミクスを調べたところ,摂食の直後にフルクトース濃度は著しく上昇することがわかった(図1).興味深いことに,この現象はフルクトースを摂取したときにかぎらず,グルコースなどほかの糖を摂取したときも同様であった.これは,過剰な糖分の一部が代謝によりフルクトースへと変換されたためと考えられた.それに対し,甘味料として使われるシュークラロースなど代謝されない糖を摂取してもフルクトース濃度に変化はなかった.また,おもな血糖であるグルコースやトレハロースは体内に大量に蓄えられているため,摂食ののちの相対的な変化はきわめて小さかった(図1).これらのことから,血中のフルクトース濃度は栄養価のある糖の摂取を反映し,きわめて大きく変動することが明らかになった.
4.Gr43aは栄養センサーである
最近の研究から,ショウジョウバエは糖類の一種ソルビトールを識別できることが明らかになっていた5,6).ソルビトールには栄養価があるが,ショウジョウバエはその味を感じることができないため,この結果は栄養センサーの存在を示唆するものであった.さきに述べた血中のフルクトースの代謝ダイナミクスの知見から,フルクトース受容体であるGr43aは栄養センサーの候補であると考えられた.それを確かめるため,水とソルビトール溶液とを同時にあたえてその摂取量を比較したところ,野生型ショウジョウバエは水よりもソルビトールを好んだが,Gr43a遺伝子に変異をもつショウジョウバエはソルビトールへの嗜好性をまったく示さなかった.このことから,Gr43aは実際に栄養センサーとして機能しており,ショウジョウバエは食物中の栄養価を血中のフルクトース濃度をつうじ感知していることが明らかになった.
5.Gr43aを発現するニューロンは空腹状態では摂食行動を促進し満腹状態では抑制する
さきに述べた摂食実験では,Gr43a変異ショウジョウバエのソルビトールの摂取量は野生型ショウジョウバエよりも少なかった.ところが,甘くて栄養価のあるシュークロースやグルコースをあたえた場合,Gr43a変異体における摂取量は逆に野生型より数十%も多かった.シュークロースの摂取量は味のないソルビトールの約3倍にもなったことから,Gr43aを発現するニューロンの作用は空腹状態あるいは満腹状態に応じ正反対になると考えられた.それを明確に示すため,甘いが栄養価のないシュークラロースをソルビトールに混ぜ,栄養価に対し影響をあたえることなくソルビトールの摂取量を増加させた.すると,野生型では摂取量が2倍程度になったが,Gr43a変異体では3倍以上に増加し野生型よりも著しく多くなった.このことから,Gr43aを発現するニューロンは空腹状態では摂食行動を促進するが,満腹状態では逆に抑制的にはたらくことが明らかになった.同一のニューロンが正反対の役割をはたすのはきわめて例外的である.
6.Gr43aを発現するニューロンの興奮は栄養状態により正反対の知覚をひき起こす
Gr43aは栄養センサーとして摂食行動をどのように制御しているのだろうか.Gr43aを発現するニューロンは食欲に直接にはたらきかけているのではなく,摂取した食物に対する知覚をつうじ間接的に影響を及ぼしているという仮説をたてた.それを確かめるため,パブロフの犬で有名な古典的条件付けを利用して,Gr43aを発現するニューロンが知覚を生じさせる可能性を検討した.そのために,まず,Gr43aを発現するニューロンを強制的に興奮させ,同時に特定の匂いをかがせることにより,その経験と匂いを関連づけて記憶させた.もし,Gr43aを発現するニューロンの興奮が知覚を生じさせるならば,それに応じ,ショウジョウバエはその匂いに対してなんらかの反応(接近または回避)を示すはずである.実験の結果,空腹状態ではGr43aを発現するニューロンの興奮は実際に正の知覚をひき起こしていることが明らかになった.重要なことに,満腹状態においてその興奮は逆に負にとらえられていた.
以上の結果は,栄養センサーは空腹状態では摂食行動を促進し,満腹状態では摂食行動を抑制することと完全に一致していた(図2).すなわち,栄養価のある食物を摂取するとGr43aを発現するニューロンが興奮し,空腹の場合にはそれを正に知覚するため,その食物の摂取に対し肯定的な評価がくだされる.すると,その行動をくり返し,結果的に栄養価のある食物の摂取量のみが増加することになる.反対に,満腹のときには栄養価のある食物を摂取すると負の知覚が生じるため,栄養価のある食物の摂取量は減少する.これらのことから,Gr43aを発現するニューロンは食物の摂取に対し栄養状態に応じた評価をあたえることにより,飽食あるいは飢餓を防いでいることが明らかになった.
おわりに
グルコース(および,昆虫ではその二糖であるトレハロース)はエネルギー代謝の基本分子であり,体内につねに高濃度で維持されているため,食物の摂取による変化はきわめて小さい.この研究から明らかになった栄養価の感知機構は,その問題を見事に解決していた.この機構のもっとも大きな長所は,栄養分子として用いるフルクトースは食物には豊富に含まれているが体内にはほとんど存在しないため,わずかな食事であってもその血中の濃度が大きく変動することである.それにくわえて,フルクトースはグルコースから容易に変換されうるため,原理的には栄養価のある(グルコースに変換されうる)すべての分子の検出が可能である.
Gr43a遺伝子は昆虫の種間でもっとも保存された味覚受容体の遺伝子のひとつであり,そのことはこの機構が昆虫にとりきわめて重要であることを示している.それでは,ヒトにも栄養センサーは存在しているのだろうか.味覚受容体ファミリー遺伝子は脊椎動物と無脊椎動物とで独自に進化したため,ヒトにはGr43a遺伝子は存在しない.しかしながら,血中におけるグルコースとフルクトースとの量比はヒトでもショウジョウバエと同じ100対1程度であり,マウスにおいては食物の栄養価を感知する能力も示されている4).栄養センサーの存在の直接の証拠は得られていないが,今後の研究が待たれるところである.
文 献
- Scott, K.: Taste recognition: food for thought. Neuron, 48, 455-464 (2005)[PubMed]
- Margolskee, R. F., Dyer, J., Kokrashvili, Z. et al.: T1R3 and gustducin in gut sense sugars to regulate expression of Na+-glucose cotransporter 1. Proc. Natl. Acad. Sci. USA, 104, 15075-15080 (2007)[PubMed]
- Singh, N., Vrontakis, M., Parkinson, F. et al.: Functional bitter taste receptors are expressed in brain cells. Biochem. Biophys. Res. Commun., 406, 146-151 (2011)[PubMed]
- de Araujo, I. E., Oliveira-Maia, A. J., Sotnikova, T. D. et al.: Food reward in the absence of taste receptor signaling. Neuron, 57, 930-941 (2008)[PubMed]
- Burke, C. J. & Waddell, S.: Remembering nutrient quality of sugar in Drosophila. Curr. Biol., 21, 746-750 (2011)[PubMed]
- Fujita, M. & Tanimura, T.: Drosophila evaluates and learns the nutritional value of sugars. Curr. Biol., 21, 751-755 (2011)[PubMed]
- Sato, K., Tanaka, K. & Touhara, K.: Sugar-regulated cation channel formed by an insect gustatory receptor. Proc. Natl. Acad. Sci. USA, 108, 11680-11685 (2011)[PubMed]
著者プロフィール
略歴:2004年 名古屋大学大学院理学研究科 修了,同年 名古屋大学遺伝子実験施設 研究員,2005年 米国Duke大学Postdoctoral fellow,2009年 米国Texas A&M Health Science Center Postdoctoral fellowを経て,2010年より同Postdoctoral Research Associate.
研究テーマ:ショウジョウバエの味覚.
抱負:個別の現象をとおして脳の機能の本質を探っていきたい.
Hubert Amrein
米国Texas A&M Health Science CenterにてProfessor.
研究室URL:http://amreinlab.wordpress.com/
© 2012 宮本徹也・Hubert Amrein Licensed under CC 表示 2.1 日本
