睡眠と覚醒を制御するドーパミン神経回路の同定
上野太郎・粂 和彦
(熊本大学発生医学研究所 多能性幹細胞分野)
email:上野太郎,粂 和彦
DOI: 10.7875/first.author.2012.135
Identification of a dopamine pathway that regulates sleep and arousal in Drosophila.
Taro Ueno, Jun Tomita, Hiromu Tanimoto, Keita Endo, Kei Ito, Shoen Kume, Kazuhiko Kume
Nature Neuroscience, 15, 1516-1523 (2012)
睡眠は昆虫から哺乳類にまで広く観察される生理現象であり,その分子機構も系統学的に保存されていることが報告されている.モノアミンであるドーパミンには覚醒作用があるが,ドーパミンは記憶の形成や注意などほかの機能も担っており,これら異なる生理機能がどのように制御されているのかはこれまで不明であった.筆者らは,モデル生物としてショウジョウバエを用いて,睡眠と覚醒を制御するドーパミン神経回路を同定した.ドーパミントランスポーターに変異をもつfumin変異体を用いた解析により,ドーパミンによる覚醒作用はD1型ドーパミン受容体であるDA1受容体を介していることがわかった.また,ドーパミンによる覚醒作用には,睡眠中枢のひとつとされる扇状体におけるDA1受容体の発現が必要であることがわかった.一方,DA1受容体は記憶の形成にかかわるとされるキノコ体において睡眠覚醒の制御には関与していなかった.また,扇状体のニューロンはドーパミンニューロンとシナプス結合しており,ドーパミンシグナルに対しcAMP濃度の上昇を示すことが確認された.さらに,モザイク個体解析法により,扇状体に投射するPPM3クラスターとよばれるドーパミンニューロンの活性化により覚醒が誘導されることが明らかになった.一方で,キノコ体に投射するドーパミンニューロンを活性化しても睡眠と覚醒に変化は認められなかった.以上のことから,扇状体に投射するドーパミン神経回路が覚醒作用を担っており,記憶の形成にかかわるドーパミン神経回路とは独立してはたらいているものと考えられた.
睡眠は動物界において広く観察される現象であるが,その機能および制御機構については不明な点が多い.睡眠覚醒の制御につき細胞レベルでの分子機構の解明において,哺乳類は神経系が複雑なため困難が多い.ショウジョウバエは比較的単純な神経系をもち,遺伝学的なツールが豊富なことから,特定の行動を制御する細胞レベルでの分子機構の解析が可能である1).ショウジョウバエにも睡眠のあることが報告され2,3),そののちの分子遺伝学的な解析により,睡眠と覚醒の分子基盤の多くは昆虫と哺乳類とのあいだで保存されていることが明らかになっている.
モノアミンであるドーパミンは哺乳類において覚醒作用をもつことが知られるが,筆者らは,2005年,ドーパミントランスポーターに変異をもつショウジョウバエが短時間睡眠の表現型を示すことを報告し,これをfumin(不眠)変異体と名づけた4).ドーパミントランスポーターはシナプス間隙に放出されたドーパミンを再取り込みする作用があり,fumin変異体ではドーパミンシグナルが増強した結果,短時間睡眠となっていると考えられた.コカインやアンフェタミンなどの覚醒剤はドーパミントランスポーターをブロックすることが知られ,同様の分子機構により覚醒作用をひき起こす薬剤である.
ドーパミンは覚醒作用を示すが,一方で,学習記憶や注意などさまざまな生理機能をもつことも知られている.ショウジョウバエにおいてもドーパミンは学習記憶に重要な神経伝達物質であり,記憶中枢とされるキノコ体に投射するドーパミンニューロンが記憶の形成を担うことが報告されている5).しかしながら,これらさまざまな生理機能がどのように実現されているのかについては,これまで明らかになっていなかった.
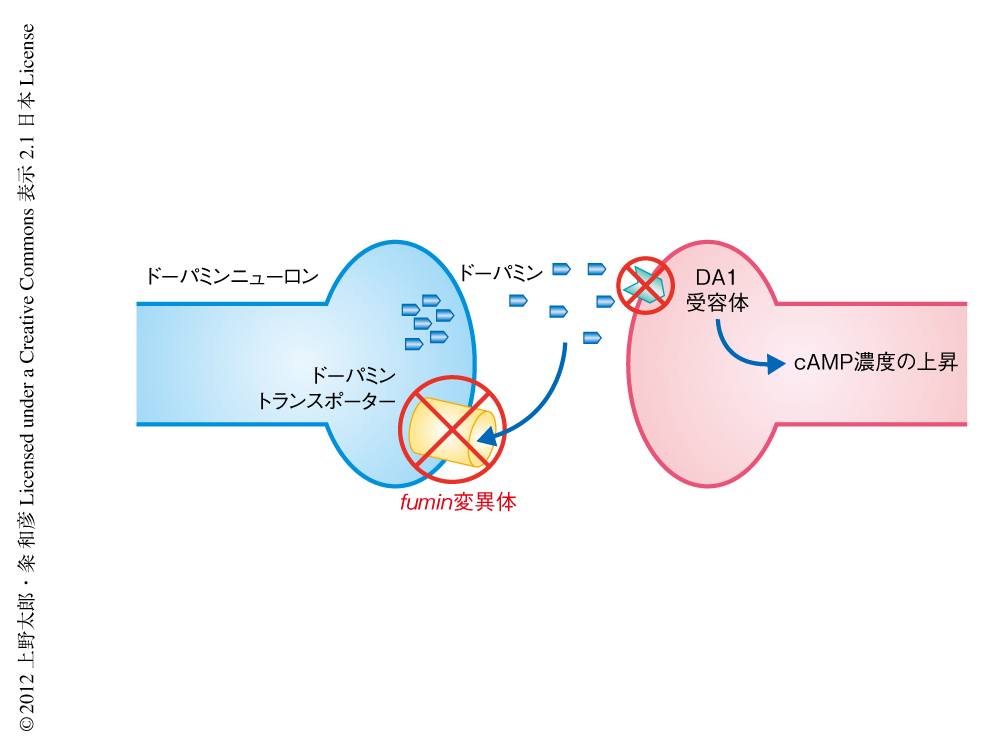
ショウジョウバエにおいても複数のドーパミン受容体が存在するが,覚醒作用に重要なドーパミン受容体を明らかにするため,ドーパミンシグナルの増強したfumin変異体を用いた解析を行った.fumin変異体は短時間睡眠の表現型を示すが,D1型ドーパミン受容体であるDA1受容体の変異を追加することにより,この短時間睡眠の表現型は消失した(図1).DA1受容体は脳においてキノコ体や扇状体などに発現している.そこで,覚醒作用に重要な脳組織を同定するため,このDA1受容体にも変異をもつfumin変異体においてDA1受容体を組織特異的に発現させた.キノコ体においてDA1受容体を発現させたところ,睡眠時間に影響はなかった.一方,扇状体においてDA1受容体を発現させたところ,睡眠時間はfumin変異体と同様に短時間睡眠となった.
最近の研究では扇状体は睡眠中枢のひとつであることが報告されており,扇状体におけるニューロンの活性化により睡眠が誘導されることが知られている6).温度依存性チャネルであるTrpA1を用いて扇状体のニューロンを活性化したところ,野生型と同様に,fumin変異体においても睡眠の誘導が起こることが確認された.
ショウジョウバエの概日時計は脳の一部のニューロンにより担われている.この時計ニューロンのうち,PDFニューロンは概日周期の調整のみならず覚醒作用をもつことが報告されている.そこで,ドーパミンによる覚醒作用がPDFニューロンに依存しているかどうかを調べるため,その遺伝学的な除去を行った.PDFニューロンに特異的にアポトーシス誘導遺伝子を発現させ,野生型とfumin変異体における睡眠時間を調べた.その結果,PDFニューロンを除去してもfumin変異体の短時間睡眠の表現型は保持され,ドーパミンによる覚醒作用はPDFニューロンに依存していないものと考えられた.
扇状体のニューロンがドーパミンニューロンとシナプスを形成していることを確認するため,GRASP(GFP reconstitution across synaptic partners)法を用いて解析した.この方法では,蛍光タンパク質GFPを2つの断片に分け,それぞれをドーパミンニューロンと扇状体のニューロンに発現させて,GFPの再構成により蛍光が生じるかどうかを観察する.その結果,扇状体においてGFPのシグナルが観察され,ドーパミンニューロンと扇状体のニューロンとはシナプス結合しているものと考えられた.
DA1受容体はD1型ドーパミン受容体であり,セカンドメッセンジャーとしてcAMP濃度を上昇させる.そこで,扇状体のニューロンがドーパミンに反応を示すかどうかをFRET(fluorescence resonance energy transfer,蛍光共鳴エネルギー移動)法により検討した.扇状体にcAMPのFRETセンサーを発現させたショウジョウバエの脳を摘出し,培養下においてドーパミンを投与した.顕微鏡下においてドーパミンを投与することにより,FRETシグナルの増強がみられcAMP濃度の上昇が確認された.また,テトロドトキシンの前処理によってもcAMP濃度の上昇は消失しなかったことから,別のニューロンの活動を介した二次的な影響ではなく,扇状体のニューロンがドーパミンに反応しているものと考えられた.さらに,扇状体においてDA1受容体をノックダウンしたショウジョウバエでは,ドーパミンに対するcAMP濃度の上昇は消失することがわかった.
覚醒作用を担うドーパミンシグナルを受け取る脳領域はわかったが,では,どのドーパミンニューロンが睡眠覚醒を制御しているのだろうか? 細胞レベルでの同定のため,ドーパミンニューロンを時期特異的に活性化した.温度依存性チャネルであるTrpA1をドーパミンニューロンの全体に発現させ,温度の上昇によりドーパミンニューロンを活性化したところ,睡眠時間の著しい減少がみられた.つぎに,記憶の形成にかかわるとされるドーパミンニューロンにおいてTrpA1チャネルを発現させ同様の実験を行った.その結果,睡眠時間の短縮はみられず,これらのドーパミンニューロンは睡眠と覚醒には関与しないことがわかった.さらに,ドーパミンニューロンはエタノールに対する反応にもかかわることが知られているが,この反応に重要なドーパミンニューロンを活性化させた場合も同様に睡眠時間の短縮は認められなかった.
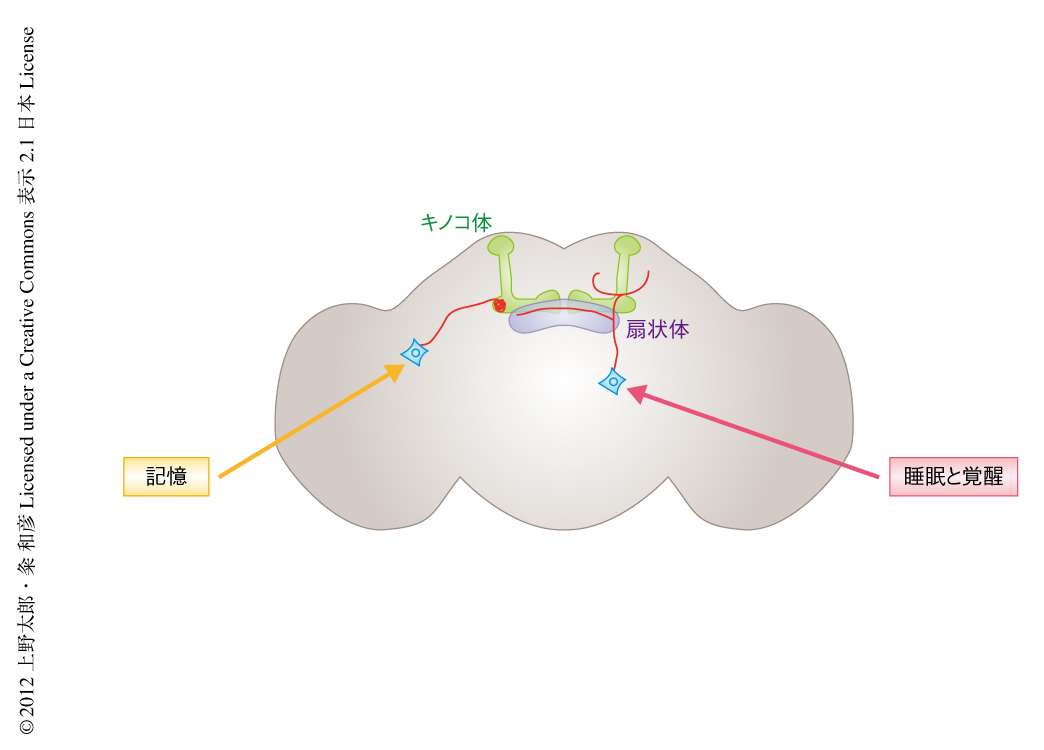
睡眠と覚醒を制御するドーパミンニューロンを1細胞レベルで同定するため,モザイク個体解析法を用いた絞り込みを行った.TrpA1チャネルと同時にGFPの発現を誘導し,細胞分裂のときに染色体の組換えによりモザイク個体を作製した.これら多数のモザイク個体において1匹ずつ,温度の上昇によりニューロンを活性化したときの睡眠を解析した.そののち,解剖によりGFPの発現を確認し,どのドーパミンニューロンが活性化したのかを同定した.細胞体の位置によりクラスター分類して睡眠時間を比較したところ,PPM3クラスターとよばれるドーパミンニューロンの活性化により睡眠時間の短縮することがわかった.さらに,PPM3クラスターのドーパミンニューロンは扇状体と楕円体に投射しているが,このうち,扇状体に投射するドーパミンニューロンを活性化したときにのみ,睡眠時間の短縮することがわかった(図2).
睡眠には覚醒しているとき形成された記憶を定着させる作用が知られており,ショウジョウバエにおいても睡眠の奪取は記憶の低下をひき起こすことが知られている7).一方,扇状体のニューロンの活性化により睡眠を人工的に誘導したショウジョウバエでは長期記憶の増強することが報告されている.睡眠が記憶を増強させる分子機構の解明が今後の課題のひとつである.
睡眠は記憶の増強をひき起こすが,睡眠中に外部からの刺激は遮断されるため新たな記憶を形成することは困難であると考えられてきた.しかし,ヒトにおける最近の研究では,睡眠中に匂いと音の連合学習が成立するという報告がある8).今回の筆者らの研究は,ショウジョウバエを用いた研究であるが,記憶を形成するドーパミン神経回路と睡眠覚醒を制御するドーパミン神経回路とは独立であることが明らかになった.このため,少なくともショウジョウバエにおいては,睡眠中に記憶の形成にかかわるドーパミンニューロンを活性化させることにより記憶の書き込みを制御することが可能であると考えられる.
睡眠が記憶を増強する一方,覚醒しているときの経験が睡眠に影響をあたえることも知られ,シナプス可塑性の関係からシナプスホメオスタシス仮説がとなえられている9).ショウジョウバエを用いたシナプスのイメージング法により,断眠されたショウジョウバエにおいてはシナプスタンパク質が増加しており,睡眠によりシナプス強度は適正化されていると考えられている10).睡眠の機能や分子機構にはまだ謎が多く,その分子基盤の解明に分子遺伝学の牽引役としてショウジョウバエのはたす役割が期待される.
略歴:2012年 熊本大学大学院医学系研究科 修了,同年より熊本大学発生医学研究所 博士研究員.
研究テーマ:睡眠.
粂 和彦(Kazuhiko Kume)
熊本大学発生医学研究所 准教授.
© 2012 上野太郎・粂 和彦Licensed under CC 表示 2.1 日本
(熊本大学発生医学研究所 多能性幹細胞分野)
email:上野太郎,粂 和彦
DOI: 10.7875/first.author.2012.135
Identification of a dopamine pathway that regulates sleep and arousal in Drosophila.
Taro Ueno, Jun Tomita, Hiromu Tanimoto, Keita Endo, Kei Ito, Shoen Kume, Kazuhiko Kume
Nature Neuroscience, 15, 1516-1523 (2012)
この論文に出現する遺伝子・タンパク質のUniprot ID
要 約
睡眠は昆虫から哺乳類にまで広く観察される生理現象であり,その分子機構も系統学的に保存されていることが報告されている.モノアミンであるドーパミンには覚醒作用があるが,ドーパミンは記憶の形成や注意などほかの機能も担っており,これら異なる生理機能がどのように制御されているのかはこれまで不明であった.筆者らは,モデル生物としてショウジョウバエを用いて,睡眠と覚醒を制御するドーパミン神経回路を同定した.ドーパミントランスポーターに変異をもつfumin変異体を用いた解析により,ドーパミンによる覚醒作用はD1型ドーパミン受容体であるDA1受容体を介していることがわかった.また,ドーパミンによる覚醒作用には,睡眠中枢のひとつとされる扇状体におけるDA1受容体の発現が必要であることがわかった.一方,DA1受容体は記憶の形成にかかわるとされるキノコ体において睡眠覚醒の制御には関与していなかった.また,扇状体のニューロンはドーパミンニューロンとシナプス結合しており,ドーパミンシグナルに対しcAMP濃度の上昇を示すことが確認された.さらに,モザイク個体解析法により,扇状体に投射するPPM3クラスターとよばれるドーパミンニューロンの活性化により覚醒が誘導されることが明らかになった.一方で,キノコ体に投射するドーパミンニューロンを活性化しても睡眠と覚醒に変化は認められなかった.以上のことから,扇状体に投射するドーパミン神経回路が覚醒作用を担っており,記憶の形成にかかわるドーパミン神経回路とは独立してはたらいているものと考えられた.
はじめに
睡眠は動物界において広く観察される現象であるが,その機能および制御機構については不明な点が多い.睡眠覚醒の制御につき細胞レベルでの分子機構の解明において,哺乳類は神経系が複雑なため困難が多い.ショウジョウバエは比較的単純な神経系をもち,遺伝学的なツールが豊富なことから,特定の行動を制御する細胞レベルでの分子機構の解析が可能である1).ショウジョウバエにも睡眠のあることが報告され2,3),そののちの分子遺伝学的な解析により,睡眠と覚醒の分子基盤の多くは昆虫と哺乳類とのあいだで保存されていることが明らかになっている.
モノアミンであるドーパミンは哺乳類において覚醒作用をもつことが知られるが,筆者らは,2005年,ドーパミントランスポーターに変異をもつショウジョウバエが短時間睡眠の表現型を示すことを報告し,これをfumin(不眠)変異体と名づけた4).ドーパミントランスポーターはシナプス間隙に放出されたドーパミンを再取り込みする作用があり,fumin変異体ではドーパミンシグナルが増強した結果,短時間睡眠となっていると考えられた.コカインやアンフェタミンなどの覚醒剤はドーパミントランスポーターをブロックすることが知られ,同様の分子機構により覚醒作用をひき起こす薬剤である.
ドーパミンは覚醒作用を示すが,一方で,学習記憶や注意などさまざまな生理機能をもつことも知られている.ショウジョウバエにおいてもドーパミンは学習記憶に重要な神経伝達物質であり,記憶中枢とされるキノコ体に投射するドーパミンニューロンが記憶の形成を担うことが報告されている5).しかしながら,これらさまざまな生理機能がどのように実現されているのかについては,これまで明らかになっていなかった.
1.ドーパミンは扇状体のDA1受容体を介し覚醒を制御する
ショウジョウバエにおいても複数のドーパミン受容体が存在するが,覚醒作用に重要なドーパミン受容体を明らかにするため,ドーパミンシグナルの増強したfumin変異体を用いた解析を行った.fumin変異体は短時間睡眠の表現型を示すが,D1型ドーパミン受容体であるDA1受容体の変異を追加することにより,この短時間睡眠の表現型は消失した(図1).DA1受容体は脳においてキノコ体や扇状体などに発現している.そこで,覚醒作用に重要な脳組織を同定するため,このDA1受容体にも変異をもつfumin変異体においてDA1受容体を組織特異的に発現させた.キノコ体においてDA1受容体を発現させたところ,睡眠時間に影響はなかった.一方,扇状体においてDA1受容体を発現させたところ,睡眠時間はfumin変異体と同様に短時間睡眠となった.
最近の研究では扇状体は睡眠中枢のひとつであることが報告されており,扇状体におけるニューロンの活性化により睡眠が誘導されることが知られている6).温度依存性チャネルであるTrpA1を用いて扇状体のニューロンを活性化したところ,野生型と同様に,fumin変異体においても睡眠の誘導が起こることが確認された.
ショウジョウバエの概日時計は脳の一部のニューロンにより担われている.この時計ニューロンのうち,PDFニューロンは概日周期の調整のみならず覚醒作用をもつことが報告されている.そこで,ドーパミンによる覚醒作用がPDFニューロンに依存しているかどうかを調べるため,その遺伝学的な除去を行った.PDFニューロンに特異的にアポトーシス誘導遺伝子を発現させ,野生型とfumin変異体における睡眠時間を調べた.その結果,PDFニューロンを除去してもfumin変異体の短時間睡眠の表現型は保持され,ドーパミンによる覚醒作用はPDFニューロンに依存していないものと考えられた.
2.ドーパミンニューロンは扇状体のニューロンとシナプス結合している
扇状体のニューロンがドーパミンニューロンとシナプスを形成していることを確認するため,GRASP(GFP reconstitution across synaptic partners)法を用いて解析した.この方法では,蛍光タンパク質GFPを2つの断片に分け,それぞれをドーパミンニューロンと扇状体のニューロンに発現させて,GFPの再構成により蛍光が生じるかどうかを観察する.その結果,扇状体においてGFPのシグナルが観察され,ドーパミンニューロンと扇状体のニューロンとはシナプス結合しているものと考えられた.
3.扇状体のニューロンはドーパミンに反応する
DA1受容体はD1型ドーパミン受容体であり,セカンドメッセンジャーとしてcAMP濃度を上昇させる.そこで,扇状体のニューロンがドーパミンに反応を示すかどうかをFRET(fluorescence resonance energy transfer,蛍光共鳴エネルギー移動)法により検討した.扇状体にcAMPのFRETセンサーを発現させたショウジョウバエの脳を摘出し,培養下においてドーパミンを投与した.顕微鏡下においてドーパミンを投与することにより,FRETシグナルの増強がみられcAMP濃度の上昇が確認された.また,テトロドトキシンの前処理によってもcAMP濃度の上昇は消失しなかったことから,別のニューロンの活動を介した二次的な影響ではなく,扇状体のニューロンがドーパミンに反応しているものと考えられた.さらに,扇状体においてDA1受容体をノックダウンしたショウジョウバエでは,ドーパミンに対するcAMP濃度の上昇は消失することがわかった.
4.記憶の形成を制御するドーパミンニューロンの睡眠と覚醒への影響
覚醒作用を担うドーパミンシグナルを受け取る脳領域はわかったが,では,どのドーパミンニューロンが睡眠覚醒を制御しているのだろうか? 細胞レベルでの同定のため,ドーパミンニューロンを時期特異的に活性化した.温度依存性チャネルであるTrpA1をドーパミンニューロンの全体に発現させ,温度の上昇によりドーパミンニューロンを活性化したところ,睡眠時間の著しい減少がみられた.つぎに,記憶の形成にかかわるとされるドーパミンニューロンにおいてTrpA1チャネルを発現させ同様の実験を行った.その結果,睡眠時間の短縮はみられず,これらのドーパミンニューロンは睡眠と覚醒には関与しないことがわかった.さらに,ドーパミンニューロンはエタノールに対する反応にもかかわることが知られているが,この反応に重要なドーパミンニューロンを活性化させた場合も同様に睡眠時間の短縮は認められなかった.
5.扇状体に投射するPPM3ドーパミンニューロンは覚醒を制御する
睡眠と覚醒を制御するドーパミンニューロンを1細胞レベルで同定するため,モザイク個体解析法を用いた絞り込みを行った.TrpA1チャネルと同時にGFPの発現を誘導し,細胞分裂のときに染色体の組換えによりモザイク個体を作製した.これら多数のモザイク個体において1匹ずつ,温度の上昇によりニューロンを活性化したときの睡眠を解析した.そののち,解剖によりGFPの発現を確認し,どのドーパミンニューロンが活性化したのかを同定した.細胞体の位置によりクラスター分類して睡眠時間を比較したところ,PPM3クラスターとよばれるドーパミンニューロンの活性化により睡眠時間の短縮することがわかった.さらに,PPM3クラスターのドーパミンニューロンは扇状体と楕円体に投射しているが,このうち,扇状体に投射するドーパミンニューロンを活性化したときにのみ,睡眠時間の短縮することがわかった(図2).
おわりに
睡眠には覚醒しているとき形成された記憶を定着させる作用が知られており,ショウジョウバエにおいても睡眠の奪取は記憶の低下をひき起こすことが知られている7).一方,扇状体のニューロンの活性化により睡眠を人工的に誘導したショウジョウバエでは長期記憶の増強することが報告されている.睡眠が記憶を増強させる分子機構の解明が今後の課題のひとつである.
睡眠は記憶の増強をひき起こすが,睡眠中に外部からの刺激は遮断されるため新たな記憶を形成することは困難であると考えられてきた.しかし,ヒトにおける最近の研究では,睡眠中に匂いと音の連合学習が成立するという報告がある8).今回の筆者らの研究は,ショウジョウバエを用いた研究であるが,記憶を形成するドーパミン神経回路と睡眠覚醒を制御するドーパミン神経回路とは独立であることが明らかになった.このため,少なくともショウジョウバエにおいては,睡眠中に記憶の形成にかかわるドーパミンニューロンを活性化させることにより記憶の書き込みを制御することが可能であると考えられる.
睡眠が記憶を増強する一方,覚醒しているときの経験が睡眠に影響をあたえることも知られ,シナプス可塑性の関係からシナプスホメオスタシス仮説がとなえられている9).ショウジョウバエを用いたシナプスのイメージング法により,断眠されたショウジョウバエにおいてはシナプスタンパク質が増加しており,睡眠によりシナプス強度は適正化されていると考えられている10).睡眠の機能や分子機構にはまだ謎が多く,その分子基盤の解明に分子遺伝学の牽引役としてショウジョウバエのはたす役割が期待される.
文 献
- Kohatsu, S., Koganezawa, M. & Yamamoto, D.: Female contact activates male-specific interneurons that trigger stereotypic courtship behavior in Drosophila. Neuron, 69, 498-508 (2011)[PubMed ] [新着論文レビュー]
- Hendricks, J. C., Finn, S. M., Panckeri, K. A. et al.: Rest in Drosophila is a sleep-like state. Neuron, 25, 129-138 (2000)[PubMed]
- Shaw, P. J., Cirelli, C., Greenspan, R. J. et al.: Correlates of sleep and waking in Drosophila melanogaster. Science, 287, 1834-1837 (2000)[PubMed]
- Kume, K., Kume, S., Park, S. K. et al.: Dopamine is a regulator of arousal in the fruit fly. J. Neurosci., 25, 7377-7384 (2005)[PubMed]
- Aso, Y., Siwanowicz, I., Bracker, L. et al.: Specific dopaminergic neurons for the formation of labile aversive memory. Curr. Biol., 20, 1445-1451 (2010)[PubMed]
- Donlea, J. M., Thimgan, M. S., Suzuki, Y. et al.: Inducing sleep by remote control facilitates memory consolidation in Drosophila. Science, 332, 1571-1576 (2011)[PubMed]
- Li, X., Yu, F. & Guo, A.: Sleep deprivation specifically impairs short-term olfactory memory in Drosophila. Sleep, 32, 1417-1424 (2009)[PubMed]
- Arzi, A., Shedlesky, L., Ben-Shaul, M. et al.: Humans can learn new information during sleep. Nat. Neurosci., 15, 1460-1465 (2012)[PubMed]
- Tononi, G. & Cirelli, C.: Sleep function and synaptic homeostasis. Sleep Med. Rev., 10, 49-62 (2006)[PubMed]
- Gilestro, G. F., Tononi, G. & Cirelli, C.: Widespread changes in synaptic markers as a function of sleep and wakefulness in Drosophila. Science, 324, 109-112 (2009)[PubMed]
著者プロフィール
略歴:2012年 熊本大学大学院医学系研究科 修了,同年より熊本大学発生医学研究所 博士研究員.
研究テーマ:睡眠.
粂 和彦(Kazuhiko Kume)
熊本大学発生医学研究所 准教授.
© 2012 上野太郎・粂 和彦Licensed under CC 表示 2.1 日本
