腸管上皮においてDll1を発現する分泌系の前駆細胞は腸管への傷害により幹細胞へと脱分化する
佐藤俊朗・Hans Clevers
(オランダHubrecht Institute)
email:佐藤俊朗
DOI: 10.7875/first.author.2012.133
Dll1+ secretory progenitor cells revert to stem cells upon crypt damage.
Johan H. van Es, Toshiro Sato, Marc van de Wetering, Anna Lyubimova, Annie Ng Yee Nee, Alex Gregorieff, Nobuo Sasaki, Laura Zeinstra, Maaike van den Born, Jeroen Korving, Anton C. M. Martens, Nick Barker, Alexander van Oudenaarden, Hans Clevers
Nature Cell Biology, 14, 1099-1104 (2012)
腸管上皮はLgr5幹細胞から分化した子孫細胞により構成されている.筆者らは,トランスジェニックマウスを用いてDll1を発現する細胞の子孫細胞の解析を行った.Dll1発現細胞は一過性に増殖し,杯細胞,内分泌細胞,Paneth細胞など分泌系の分化細胞を産生したが,Lgr5幹細胞のようにすべての系統の細胞を永続的に産生することはなかった.このことから,Dll1発現細胞は幹細胞ではなく分泌系の前駆細胞であることがわかった.in vitroにおいてDll1発現細胞をWnt3Aとともに培養するとLgr5幹細胞を産み出すことがわかった.また,in vivoにおいても,放射線の照射によりLgr5幹細胞を除去するとDll1発現細胞は幹細胞へと脱分化することがわかった.これらのことから,腸管上皮幹細胞のヒエラルキーは細胞のおかれた微小環境により可塑性を示すことがわかった.
腸管上皮は,腸管の内腔へと突出した絨毛と,腸管の粘膜へと湾入した陰窩から構成されている.陰窩はおもに未分化な細胞により構成され,その底部に存在する幹細胞は自己複製により幹細胞それ自体と分化した娘細胞とを産生している.娘細胞は数回の分裂とともに陰窩を上行し,絨毛に移行すると増殖を停止して成熟し分化する.成熟した腸管上皮の大部分は吸収上皮細胞であり,それ以外の分化した上皮細胞は分泌構造をもち分泌系細胞とよばれている.分泌系細胞には,杯細胞,腸管内分泌細胞,Paneth細胞がある.分泌系細胞への運命決定は前駆細胞のレベルで起こっており,Notchシグナルの活性化した前駆細胞ではHes1が活性化し吸収上皮細胞へと分化していく.一方,Notchシグナルが不活性化しAtoh1を発現するようになった前駆細胞は分泌系細胞へと分化することがわかっている1).Notchシグナルが不活性化したことによりAtoh1を発現する細胞ではNotchリガンドの発現が上昇し,近接する細胞のNotchシグナルを活性化するため,分泌系細胞のまわりの細胞は吸収上皮細胞へと分化する(図1).
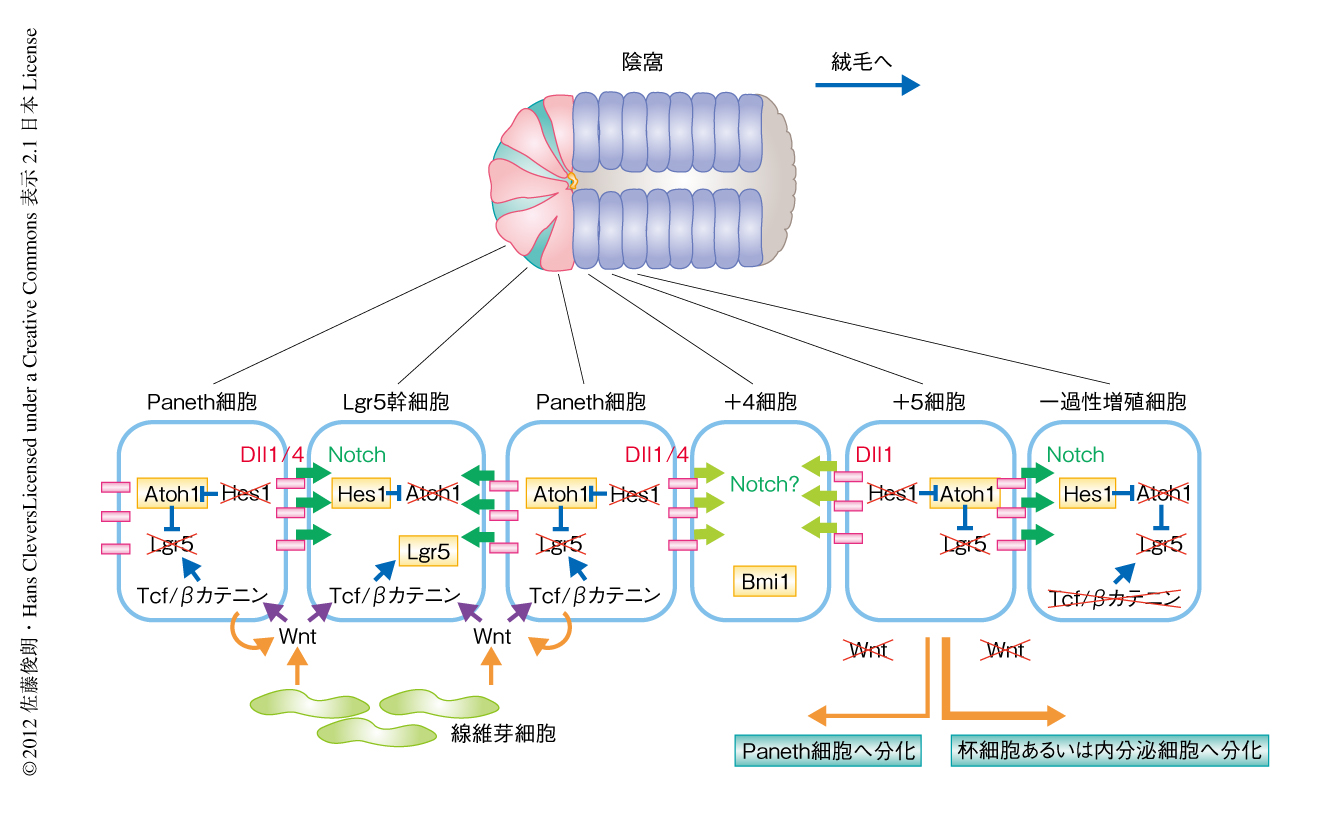
腸管上皮幹細胞はLgr5を発現し,Paneth細胞にはさまれた局在パターンをとる2).筆者らは,Lgr5を発現する細胞を単離し,EGF,Noggin,R-spondin1と,細胞外基質であるマトリジェルとともに培養することで,陰窩-絨毛構造を擬似した組織構造体(オルガノイド)を形成することを報告した3).この培養は無血清培養であったため,幹細胞維持タンパク質がほぼ解明された.幹細胞からのオルガノイドの形成がPaneth細胞の存在により飛躍的に向上したこと,また,Paneth細胞は幹細胞維持タンパク質であるWnt3,NotchリガンドであるDll4,EGFを発現していたことから,Paneth細胞は腸管上皮幹細胞のニッチであることがわかった4)(新着論文レビュー でも掲載).このように,腸管上皮幹細胞はその自己複製の機構やニッチがつぎつぎと解明されたが,幹細胞ニッチまたは幹細胞維持タンパク質は,非幹細胞を幹細胞へと脱分化させることができるかどうかわかっていない.今回,筆者らは,非幹細胞のマーカーであるDll1を用い,幹細胞の可塑性につき研究した.
まず,in situハイブリダイゼーション法により,Dll1は陰窩の底部より5~6番目の細胞(+5細胞)に強く発現しているが,Lgr5幹細胞には発現していないことを確認した.つづいて,Dll1遺伝子座に蛍光タンパク質GFPの遺伝子とタモキシフェン誘導型Creリコンビナーゼの遺伝子を組み込んだノックインマウスを作製した.さらに,このノックインマウスをCreリコンビナーゼに依存的にLacZを発現するトランスジェニックマウスと交配した.得られたマウスはタモキシフェンの投与によるCreリコンビナーゼの誘導により,Dll1発現細胞とその子孫細胞がLacZの発現により標識され,遺伝学的な細胞系譜の解析が可能となった.Dll1遺伝子座にノックインされたGFPは,in situハイブリダイゼーション法によりDll1の発現が検出された+5細胞のほか,成熟した分泌系細胞である杯細胞,内分泌細胞,Paneth細胞において発現した.一方,タモキシフェンの投与では+5細胞にのみLacZの発現が誘導され,ほかの分泌系細胞ではLacZは発現しなかった.腸管上皮においては通常の感度のin situハイブリダイゼーション法よりGFP遺伝子のノックインでのGFPの検出のほうが高感度であることがわかっており,+5細胞はほかの分泌系細胞よりDll1の発現レベルが高いことが示唆された.このため,Creリコンビナーゼの発現は+5細胞でのみ遺伝子組換えに十分なレベルにまで達していたものと考えられた.タモキシフェンの投与ののちのLacZの発現を経時的にを追跡すると,2日目まで+5細胞の一過性の増殖が認められたが,それ以降は新たなLacZ陽性細胞の産生は認められなかった.このことは,永続的な自己複製を示すLgr5幹細胞とは対照的であった.
LacZ陽性細胞には,経時的に絨毛へと上行するものと,陰窩の底部へと下行するものの2種類が認められた.腸管上皮細胞の分化マーカーとの二重染色により,これらの細胞はすべて分泌系細胞であることがわかった.つまり,絨毛においては杯細胞,内分泌細胞,Tuft細胞であり,吸収上皮細胞においてLacZによる標識は認められなかった.また,陰窩の底部の細胞はPaneth細胞であった(図1).
+5細胞は分泌系の前駆細胞であり,一過性の増殖ののち複数の分泌系細胞に分化する多分化能をもつと考えられていたが,その存在ははっきり証明されてはいなかった.さきのLacZの発現による細胞系譜の追跡実験の結果からは,+5細胞は単一の細胞系譜にしか分化を示さない未熟な分泌細胞(たとえば,未熟な杯細胞から成熟した杯細胞への分化)である可能性を除外できなかった.そこで,Creリコンビナーゼの活性化により4色の蛍光タンパク質からランダムに1色が遺伝学的に標識されるトランスジェニックマウス5) と,Dll1遺伝子座にGFP遺伝子とタモキシフェン誘導型Creリコンビナーゼの遺伝子を組み込んだノックインマウスとを交配し,さきと同様に細胞系譜の追跡実験を行った.このマウスにおいて,1色の蛍光タンパク質を発現した複数の分泌系細胞が同一の陰窩,または,同一の陰窩-絨毛軸において観察された.分泌系細胞は2種類の蛍光タンパク質を発現した細胞からなることもあり,+5細胞は分泌系の前駆細胞であり,一過性の増殖とともに多系統の分泌系細胞へと分化しうることが証明された.
Dll1遺伝子座にGFP遺伝子とタモキシフェン誘導型Creリコンビナーゼの遺伝子を組み込んだノックインマウスにおいて,GFP陽性細胞は+5細胞(分泌系の前駆細胞)のほか,Paneth細胞,内分泌細胞,杯細胞に認められたが,これらはCD24の発現レベルにより区別することが可能であった.つまり,CD24を高発現する細胞はPaneth細胞と内分泌細胞,CD24を発現しない細胞は杯細胞,CD24を低発現する細胞は一過性に増殖する細胞と幹細胞であり,+5細胞はCD24低発現GFP陽性細胞にあたると考えられた.実際に,このような細胞をフローサイトメトリーにより純化し,マイクロアレイを用いて遺伝子発現プロファイルを解析したところ,Neurogenin3などの内分泌系の前駆細胞マーカーを発現していたことから,+5細胞であると考えられた.+5細胞をWntの存在しない状態において培養したところコロニーを形成せず,in vivoにおける細胞系譜の追跡実験の結果と合致して,幹細胞としての機能をもたなかった.一方,+5細胞をWnt3Aの存在のもと培養すると,コロニーを形成し永続的な培養と多分化能が示され,また,このコロニーにはLgr5幹細胞が確認された.このことから,+5細胞はWntによる刺激により幹細胞としての機能を獲得することが示唆された.
in vitroにおいて観察された+5細胞の腸管上皮幹細胞への脱分化はin vivoにおいても観察されるのか,腸管上皮障害モデルを用いて検証した.6 Gyの放射線の照射はLgr5幹細胞を含む大部分の増殖細胞にアポトーシスを誘導することがわかっている.Dll1遺伝子座にGFP遺伝子とタモキシフェン誘導型Creリコンビナーゼの遺伝子を組み込んだノックインマウスとCreリコンビナーゼに依存的にLacZを発現するトランスジェニックマウスとを掛け合わせたマウスに,放射線の照射とともにタモキシフェンを投与し,腸管を傷害したときのDll1発現細胞の変化を解析した.放射線の照射をしない対照のマウスではLacZ陽性細胞は半減期の長いPaneth細胞を除き7日後には消失していた.一方,放射線を照射したマウスでは長期間にわたりLacZの発現が観察された.LacZの発現パターンは,Lgr5遺伝子座に対する同様のトランスジェニックマウスを用いた実験でみられた,陰窩-絨毛軸のすべてにLacZを発現するような幹細胞の細胞系譜を産生しうることがわかった.LacZ陽性に置き換わった陰窩にはLgr5を発現する細胞があり,Dll1発現細胞がin vivoにおいても腸管への傷害により幹細胞へと脱分化することがわかった.
腸管への傷害によりLgr5幹細胞が除去されたのち,3~4日後にはLgr5幹細胞が再出現することがわかっている.最近,Bmi1を発現する+4細胞は,Lgr5幹細胞を除去したとき活性化され新しいLgr5幹細胞を産み出す“予備の”幹細胞として機能することが報告された6).+4細胞は陰窩の底部より数えて4番目の細胞であり,Lgr5幹細胞とは異なり定常状態ではほとんど分裂しない別の幹細胞であると考えられてきた7).最近,より高感度のin situハイブリダイゼーション法により,Bmi1はより広い範囲に発現しておりLgr5幹細胞の75%はBmi1を発現していることが報告された8).さらに,Bmi1は陰窩においてさまざまな細胞に発現しており,それらの細胞の大部分は幹細胞としての細胞系譜を示さないことが報告された9).
Dll1発現細胞は定常状態では幹細胞としての細胞系譜を示さなかったことから,幹細胞にはDll1は発現していないことがわかった.一方,放射線の照射による腸管の傷害によりLgr5幹細胞を除去した場合,Dll1発現細胞はLgr5を発現する細胞を産み出し幹細胞となることがわかった.+4細胞は定常状態ではほとんど増殖せず,腸管への傷害により活性化しLgr5幹細胞を産生するのに対し,Dll1を発現する+5細胞は定常状態では前駆細胞としてはたらき,腸管への障害により幹細胞へと脱分化することがわかった.
in vitroにおいて,Dll1発現細胞はWntの刺激によりLgr5幹細胞を産生したことから,脱分化にはWntシグナルが重要であることが示された.腸管への傷害によりLgr5幹細胞が除去されると,+5細胞はWnt3を産生するPaneth細胞と接触できるようになると考えられ,前駆細胞において幹細胞ニッチの獲得が脱分化を誘導しているものと推察された.最近,Paneth細胞のほか,線維芽細胞がWnt2bを産生することにより幹細胞を維持する機能をもつことがわかり10),Dll1発現細胞における腸管への傷害による幹細胞としての機能の獲得に,線維芽細胞に由来するWntの関与している可能性も否定できない.
前駆細胞の幹細胞への脱分化はほかの組織幹細胞でも報告されており,組織幹細胞のヒエラルキーにはある程度の可塑性が許容されることがわかってきた11).いずれの報告も,前駆細胞から幹細胞への脱分化であり,分化の進んだ細胞から幹細胞への脱分化は確認されていない.脱分化により幹細胞としての機能を獲得する前駆細胞は,20年前に,潜在性の幹細胞として提唱されていたが,近年の研究により,実験的にその存在が確認されたことになる12).こうした潜在性の幹細胞はがん組織にも存在する可能性があり,がん幹細胞の標的治療を考えるうえで重要な概念になるだろう.
略歴:2004年 慶應義塾大学大学院医学研究科にて博士号取得,同年 慶應義塾大学医学部 研究員,2006年 米国Stowers Institute for Medical Research研究員,2007年 オランダHubrecht Institute研究員を経て,2011年4月より慶應義塾大学医学部 特任講師.
研究テーマ:腸管上皮幹細胞,炎症性腸疾患,大腸がん.
Hans Clevers
オランダHubrecht InstituteにてGroup Leader.
研究室URL:http://www.hubrecht.eu/research/clevers/index.html
© 2012 佐藤俊朗・Hans Clevers Licensed under CC 表示 2.1 日本
(オランダHubrecht Institute)
email:佐藤俊朗
DOI: 10.7875/first.author.2012.133
Dll1+ secretory progenitor cells revert to stem cells upon crypt damage.
Johan H. van Es, Toshiro Sato, Marc van de Wetering, Anna Lyubimova, Annie Ng Yee Nee, Alex Gregorieff, Nobuo Sasaki, Laura Zeinstra, Maaike van den Born, Jeroen Korving, Anton C. M. Martens, Nick Barker, Alexander van Oudenaarden, Hans Clevers
Nature Cell Biology, 14, 1099-1104 (2012)
要 約
腸管上皮はLgr5幹細胞から分化した子孫細胞により構成されている.筆者らは,トランスジェニックマウスを用いてDll1を発現する細胞の子孫細胞の解析を行った.Dll1発現細胞は一過性に増殖し,杯細胞,内分泌細胞,Paneth細胞など分泌系の分化細胞を産生したが,Lgr5幹細胞のようにすべての系統の細胞を永続的に産生することはなかった.このことから,Dll1発現細胞は幹細胞ではなく分泌系の前駆細胞であることがわかった.in vitroにおいてDll1発現細胞をWnt3Aとともに培養するとLgr5幹細胞を産み出すことがわかった.また,in vivoにおいても,放射線の照射によりLgr5幹細胞を除去するとDll1発現細胞は幹細胞へと脱分化することがわかった.これらのことから,腸管上皮幹細胞のヒエラルキーは細胞のおかれた微小環境により可塑性を示すことがわかった.
はじめに
腸管上皮は,腸管の内腔へと突出した絨毛と,腸管の粘膜へと湾入した陰窩から構成されている.陰窩はおもに未分化な細胞により構成され,その底部に存在する幹細胞は自己複製により幹細胞それ自体と分化した娘細胞とを産生している.娘細胞は数回の分裂とともに陰窩を上行し,絨毛に移行すると増殖を停止して成熟し分化する.成熟した腸管上皮の大部分は吸収上皮細胞であり,それ以外の分化した上皮細胞は分泌構造をもち分泌系細胞とよばれている.分泌系細胞には,杯細胞,腸管内分泌細胞,Paneth細胞がある.分泌系細胞への運命決定は前駆細胞のレベルで起こっており,Notchシグナルの活性化した前駆細胞ではHes1が活性化し吸収上皮細胞へと分化していく.一方,Notchシグナルが不活性化しAtoh1を発現するようになった前駆細胞は分泌系細胞へと分化することがわかっている1).Notchシグナルが不活性化したことによりAtoh1を発現する細胞ではNotchリガンドの発現が上昇し,近接する細胞のNotchシグナルを活性化するため,分泌系細胞のまわりの細胞は吸収上皮細胞へと分化する(図1).
腸管上皮幹細胞はLgr5を発現し,Paneth細胞にはさまれた局在パターンをとる2).筆者らは,Lgr5を発現する細胞を単離し,EGF,Noggin,R-spondin1と,細胞外基質であるマトリジェルとともに培養することで,陰窩-絨毛構造を擬似した組織構造体(オルガノイド)を形成することを報告した3).この培養は無血清培養であったため,幹細胞維持タンパク質がほぼ解明された.幹細胞からのオルガノイドの形成がPaneth細胞の存在により飛躍的に向上したこと,また,Paneth細胞は幹細胞維持タンパク質であるWnt3,NotchリガンドであるDll4,EGFを発現していたことから,Paneth細胞は腸管上皮幹細胞のニッチであることがわかった4)(新着論文レビュー でも掲載).このように,腸管上皮幹細胞はその自己複製の機構やニッチがつぎつぎと解明されたが,幹細胞ニッチまたは幹細胞維持タンパク質は,非幹細胞を幹細胞へと脱分化させることができるかどうかわかっていない.今回,筆者らは,非幹細胞のマーカーであるDll1を用い,幹細胞の可塑性につき研究した.
1.Dll1発現細胞は分泌系細胞へと分化する
まず,in situハイブリダイゼーション法により,Dll1は陰窩の底部より5~6番目の細胞(+5細胞)に強く発現しているが,Lgr5幹細胞には発現していないことを確認した.つづいて,Dll1遺伝子座に蛍光タンパク質GFPの遺伝子とタモキシフェン誘導型Creリコンビナーゼの遺伝子を組み込んだノックインマウスを作製した.さらに,このノックインマウスをCreリコンビナーゼに依存的にLacZを発現するトランスジェニックマウスと交配した.得られたマウスはタモキシフェンの投与によるCreリコンビナーゼの誘導により,Dll1発現細胞とその子孫細胞がLacZの発現により標識され,遺伝学的な細胞系譜の解析が可能となった.Dll1遺伝子座にノックインされたGFPは,in situハイブリダイゼーション法によりDll1の発現が検出された+5細胞のほか,成熟した分泌系細胞である杯細胞,内分泌細胞,Paneth細胞において発現した.一方,タモキシフェンの投与では+5細胞にのみLacZの発現が誘導され,ほかの分泌系細胞ではLacZは発現しなかった.腸管上皮においては通常の感度のin situハイブリダイゼーション法よりGFP遺伝子のノックインでのGFPの検出のほうが高感度であることがわかっており,+5細胞はほかの分泌系細胞よりDll1の発現レベルが高いことが示唆された.このため,Creリコンビナーゼの発現は+5細胞でのみ遺伝子組換えに十分なレベルにまで達していたものと考えられた.タモキシフェンの投与ののちのLacZの発現を経時的にを追跡すると,2日目まで+5細胞の一過性の増殖が認められたが,それ以降は新たなLacZ陽性細胞の産生は認められなかった.このことは,永続的な自己複製を示すLgr5幹細胞とは対照的であった.
LacZ陽性細胞には,経時的に絨毛へと上行するものと,陰窩の底部へと下行するものの2種類が認められた.腸管上皮細胞の分化マーカーとの二重染色により,これらの細胞はすべて分泌系細胞であることがわかった.つまり,絨毛においては杯細胞,内分泌細胞,Tuft細胞であり,吸収上皮細胞においてLacZによる標識は認められなかった.また,陰窩の底部の細胞はPaneth細胞であった(図1).
2.Dll1を発現する+5細胞は分泌系の前駆細胞である
+5細胞は分泌系の前駆細胞であり,一過性の増殖ののち複数の分泌系細胞に分化する多分化能をもつと考えられていたが,その存在ははっきり証明されてはいなかった.さきのLacZの発現による細胞系譜の追跡実験の結果からは,+5細胞は単一の細胞系譜にしか分化を示さない未熟な分泌細胞(たとえば,未熟な杯細胞から成熟した杯細胞への分化)である可能性を除外できなかった.そこで,Creリコンビナーゼの活性化により4色の蛍光タンパク質からランダムに1色が遺伝学的に標識されるトランスジェニックマウス5) と,Dll1遺伝子座にGFP遺伝子とタモキシフェン誘導型Creリコンビナーゼの遺伝子を組み込んだノックインマウスとを交配し,さきと同様に細胞系譜の追跡実験を行った.このマウスにおいて,1色の蛍光タンパク質を発現した複数の分泌系細胞が同一の陰窩,または,同一の陰窩-絨毛軸において観察された.分泌系細胞は2種類の蛍光タンパク質を発現した細胞からなることもあり,+5細胞は分泌系の前駆細胞であり,一過性の増殖とともに多系統の分泌系細胞へと分化しうることが証明された.
3.Dll1発現細胞はin vitroにおいて幹細胞としての機能を獲得する
Dll1遺伝子座にGFP遺伝子とタモキシフェン誘導型Creリコンビナーゼの遺伝子を組み込んだノックインマウスにおいて,GFP陽性細胞は+5細胞(分泌系の前駆細胞)のほか,Paneth細胞,内分泌細胞,杯細胞に認められたが,これらはCD24の発現レベルにより区別することが可能であった.つまり,CD24を高発現する細胞はPaneth細胞と内分泌細胞,CD24を発現しない細胞は杯細胞,CD24を低発現する細胞は一過性に増殖する細胞と幹細胞であり,+5細胞はCD24低発現GFP陽性細胞にあたると考えられた.実際に,このような細胞をフローサイトメトリーにより純化し,マイクロアレイを用いて遺伝子発現プロファイルを解析したところ,Neurogenin3などの内分泌系の前駆細胞マーカーを発現していたことから,+5細胞であると考えられた.+5細胞をWntの存在しない状態において培養したところコロニーを形成せず,in vivoにおける細胞系譜の追跡実験の結果と合致して,幹細胞としての機能をもたなかった.一方,+5細胞をWnt3Aの存在のもと培養すると,コロニーを形成し永続的な培養と多分化能が示され,また,このコロニーにはLgr5幹細胞が確認された.このことから,+5細胞はWntによる刺激により幹細胞としての機能を獲得することが示唆された.
4.Dll1発現細胞は腸管への傷害により幹細胞へと脱分化する
in vitroにおいて観察された+5細胞の腸管上皮幹細胞への脱分化はin vivoにおいても観察されるのか,腸管上皮障害モデルを用いて検証した.6 Gyの放射線の照射はLgr5幹細胞を含む大部分の増殖細胞にアポトーシスを誘導することがわかっている.Dll1遺伝子座にGFP遺伝子とタモキシフェン誘導型Creリコンビナーゼの遺伝子を組み込んだノックインマウスとCreリコンビナーゼに依存的にLacZを発現するトランスジェニックマウスとを掛け合わせたマウスに,放射線の照射とともにタモキシフェンを投与し,腸管を傷害したときのDll1発現細胞の変化を解析した.放射線の照射をしない対照のマウスではLacZ陽性細胞は半減期の長いPaneth細胞を除き7日後には消失していた.一方,放射線を照射したマウスでは長期間にわたりLacZの発現が観察された.LacZの発現パターンは,Lgr5遺伝子座に対する同様のトランスジェニックマウスを用いた実験でみられた,陰窩-絨毛軸のすべてにLacZを発現するような幹細胞の細胞系譜を産生しうることがわかった.LacZ陽性に置き換わった陰窩にはLgr5を発現する細胞があり,Dll1発現細胞がin vivoにおいても腸管への傷害により幹細胞へと脱分化することがわかった.
おわりに
腸管への傷害によりLgr5幹細胞が除去されたのち,3~4日後にはLgr5幹細胞が再出現することがわかっている.最近,Bmi1を発現する+4細胞は,Lgr5幹細胞を除去したとき活性化され新しいLgr5幹細胞を産み出す“予備の”幹細胞として機能することが報告された6).+4細胞は陰窩の底部より数えて4番目の細胞であり,Lgr5幹細胞とは異なり定常状態ではほとんど分裂しない別の幹細胞であると考えられてきた7).最近,より高感度のin situハイブリダイゼーション法により,Bmi1はより広い範囲に発現しておりLgr5幹細胞の75%はBmi1を発現していることが報告された8).さらに,Bmi1は陰窩においてさまざまな細胞に発現しており,それらの細胞の大部分は幹細胞としての細胞系譜を示さないことが報告された9).
Dll1発現細胞は定常状態では幹細胞としての細胞系譜を示さなかったことから,幹細胞にはDll1は発現していないことがわかった.一方,放射線の照射による腸管の傷害によりLgr5幹細胞を除去した場合,Dll1発現細胞はLgr5を発現する細胞を産み出し幹細胞となることがわかった.+4細胞は定常状態ではほとんど増殖せず,腸管への傷害により活性化しLgr5幹細胞を産生するのに対し,Dll1を発現する+5細胞は定常状態では前駆細胞としてはたらき,腸管への障害により幹細胞へと脱分化することがわかった.
in vitroにおいて,Dll1発現細胞はWntの刺激によりLgr5幹細胞を産生したことから,脱分化にはWntシグナルが重要であることが示された.腸管への傷害によりLgr5幹細胞が除去されると,+5細胞はWnt3を産生するPaneth細胞と接触できるようになると考えられ,前駆細胞において幹細胞ニッチの獲得が脱分化を誘導しているものと推察された.最近,Paneth細胞のほか,線維芽細胞がWnt2bを産生することにより幹細胞を維持する機能をもつことがわかり10),Dll1発現細胞における腸管への傷害による幹細胞としての機能の獲得に,線維芽細胞に由来するWntの関与している可能性も否定できない.
前駆細胞の幹細胞への脱分化はほかの組織幹細胞でも報告されており,組織幹細胞のヒエラルキーにはある程度の可塑性が許容されることがわかってきた11).いずれの報告も,前駆細胞から幹細胞への脱分化であり,分化の進んだ細胞から幹細胞への脱分化は確認されていない.脱分化により幹細胞としての機能を獲得する前駆細胞は,20年前に,潜在性の幹細胞として提唱されていたが,近年の研究により,実験的にその存在が確認されたことになる12).こうした潜在性の幹細胞はがん組織にも存在する可能性があり,がん幹細胞の標的治療を考えるうえで重要な概念になるだろう.
文 献
- Yang, Q., Bermingham, N. A., Finegold, M. J. et al.: Requirement of Math1 for secretory cell lineage commitment in the mouse intestine. Science, 294, 2155-2158 (2001)[PubMed]
- Barker, N., van Es, J. H., Kuipers, J. et al.: Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature, 449, 1003-1007 (2007)[PubMed]
- Sato, T., Vries, R. G., Snippert, H. J. et al.: Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature, 459, 262-265 (2009)[PubMed]
- Sato, T., van Es, J. H., Snippert, H. J. et al.: Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature, 469, 415-418 (2011)[PubMed] [新着論文レビュー]
- Snippert, H. J., van der Flier, L. G., Sato, T. et al.: Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell, 143, 134-144 (2010)[PubMed]
- Tian, H., Biehs, B., Warming, S. et al.: A reserve stem cell population in small intestine renders Lgr5-positive cells dispensable. Nature, 478, 255-259 (2011)[PubMed]
- Sangiorgi, E. & Capecchi, M. R.: Bmi1 is expressed in vivo in intestinal stem cells. Nat. Genet., 40, 915-920 (2008)[PubMed]
- Itzkovitz, S., Lyubimova, A., Blat, I. C. et al.: Single-molecule transcript counting of stem-cell markers in the mouse intestine. Nat. Cell Biol., 14, 106-114 (2012)[PubMed]
- Munoz, J., Stange, D. E., Schepers, A. G. et al.: The Lgr5 intestinal stem cell signature: robust expression of proposed quiescent '+4' cell markers. EMBO J., 31, 3079-3091 (2012)[PubMed]
- Farin, H. F., Van Es, J. H. & Clevers, H.: Redundant sources of Wnt regulate intestinal stem cells and promote formation of Paneth cells. Gastroenterology, 143, 1518-1529 (2012)[PubMed]
- Nakagawa, T., Sharma, M., Nabeshima, Y. et al.: Functional hierarchy and reversibility within the murine spermatogenic stem cell compartment. Science, 328, 62-67 (2010)[PubMed]
- Potten, C. S. & Loeffler, M.: Stem cells: attributes, cycles, spirals, pitfalls and uncertainties. Lessons for and from the crypt. Development, 110, 1001-1020 (1990)[PubMed]
著者プロフィール
略歴:2004年 慶應義塾大学大学院医学研究科にて博士号取得,同年 慶應義塾大学医学部 研究員,2006年 米国Stowers Institute for Medical Research研究員,2007年 オランダHubrecht Institute研究員を経て,2011年4月より慶應義塾大学医学部 特任講師.
研究テーマ:腸管上皮幹細胞,炎症性腸疾患,大腸がん.
Hans Clevers
オランダHubrecht InstituteにてGroup Leader.
研究室URL:http://www.hubrecht.eu/research/clevers/index.html
© 2012 佐藤俊朗・Hans Clevers Licensed under CC 表示 2.1 日本
