Admp遺伝子はPinhead遺伝子によりタンパク質レベルおよび転写レベルで二重の抑制をうけ動物胚の腹側の領域を決定する
今井 (佐藤) 薫・佐藤ゆたか
(京都大学大学院理学研究科 生物科学専攻発生ゲノム科学研究室)
email:今井 (佐藤) 薫,佐藤ゆたか
DOI: 10.7875/first.author.2012.114
Cis-acting transcriptional repression establishes a sharp boundary in chordate embryos.
Kaoru S. Imai, Yutaka Daido, Takehiro G. Kusakabe, Yutaka Satou
Science, 337, 964-967 (2012)
動物胚における背側と腹側の領域の決定については,両生類の胚など脊椎動物の胚での研究によりBMPシグナル伝達系が大きな役割をはたしていることが明らかになっている.つまり,BMPシグナル伝達系は腹側の領域を誘導し,背側ではBMPシグナル伝達系は阻害される.ほかのBMPリガンドとは異なり,Admpは背側の領域で発現する.しかし,そののち腹側に輸送されて,そこでほかのBMPリガンドと同様に腹側の運命決定を行う.今回,PinheadはAdmpに特異的なアンタゴニストであり,脊索動物尾索類のカタユウレイボヤにおける表皮の腹側の領域の決定に必須であることが示された.Pinheadはこのタンパク質レベルでのAdmpの抑制にくわえ,Pinhead遺伝子の転写によりAdmp遺伝子の転写が直接的に抑制されるという二重の抑制機構をもっていた.この転写抑制はこの2つの遺伝子がゲノムにおいて隣接してコードされていることを利用していたが,この遺伝子の並びは動物のゲノムにおいて広く保存されており,この制御機構は幅広い動物において利用されているものと思われた.
動物の発生は,受精卵というひとつの細胞にはじまり,それが細胞分裂をへてさまざまな組織や器官をつくりあげていく現象である.発生において組織や器官の分化は漸進的に進んでいく.背腹軸の決定はそのなかでも初期において必要な過程であり,のちの胚発生はそこで決定された軸を利用している.この背腹軸の決定の分子機構の研究は多くの発生学者を魅了してきたが,近年,背腹軸の決定に関与するタンパク質の作用機序がつぎつぎに明らかになってきている.非常に簡単にいえば,脊椎動物では腹側においてBMPシグナル伝達系がはたらき,背側においてBMPシグナル伝達系が抑制されることが必要である.興味深いことに,BMP様のタンパク質であるAdmp(anti-dorsalising morphogenetic protein)は作用すべき腹側ではなく背側において発現する.そののち,Admpは腹側に輸送され,そこでBMP4などと協調し腹側における発生プログラムを始動する1).
筆者らは,この分子機構をさらにくわしく解析するため,カタユウレイボヤ(Ciona intestinalis)を用いて実験を行った.ホヤはわれわれ脊椎動物と同じ脊索動物であり,基本的な体制を共有している.つまり,ホヤは幼生時において脊索動物に特徴的な“オタマジャクシ型”の形態をとる.このことは,この基本的な体制をつくるための発生の機構が脊索動物において共有されていることを強く示唆する.一方で,筆者らがこれまでに明らかにしてきた遺伝子ネットワークの研究などをもとにして2),ホヤの初期胚ではBMPシグナル伝達系に依存的に背腹軸を決定する機構が失われている可能性が示唆されてきた3).しかしながら今回,筆者らは,ホヤにおいても神経胚期ののちBMPシグナルに依存的に背腹軸が決定される機構が存在することを見い出し,その実験系を使い研究を進めた.ホヤはゲノムがコンパクトなうえ4)(ゲノムサイズは160 Mb,遺伝子数は約16,000),幼生も単純なので(幼生は少数の組織から構成され,その細胞数は約2600),実験的な解析が容易という利点がある5).
BMPシグナル伝達系は動物胚における背腹の決定に重要な役割をはたす.BMPシグナル伝達系の特徴として,多くのアンタゴニストにより制御されていることがあげられる.興味深いことに,BMPリガンドとChordinなどBMPアンタゴニストの一部は,システインノットとよばれる構造を共通にもつ.そこで,BMPシグナル伝達系を調べる過程でこの構造に注目し,このシステインノット構造をもつタンパク質をコードする遺伝子をホヤのゲノムにおいてすべて調べた.このなかにはもちろん,BMPリガンドを含むTGFβスーパーファミリータンパク質の遺伝子やChordinなどのBMPアンタゴニストをコードする遺伝子など既知のものが含まれていたが,そのほか機能未知の遺伝子がいくつか同定された.なかでも,アフリカツメガエルにおいてすでに同定されていたPinhead遺伝子6) のホモログは,つぎの2つの点で興味深かった.第1に,同定されたほかすべてはシステインノット構造を1つしかもたなかったのに対し,Pinheadは3つのシステインノットをもっていた.第2に,Pinhead遺伝子はゲノムにおいてBMPリガンドのひとつをコードするAdmp遺伝子に隣接してコードされており,のちに述べるように,その遺伝子の並びは進化的に保存されていた(図1a).そこで,このPinhead遺伝子の機能をさらにくわしく調べることにした.
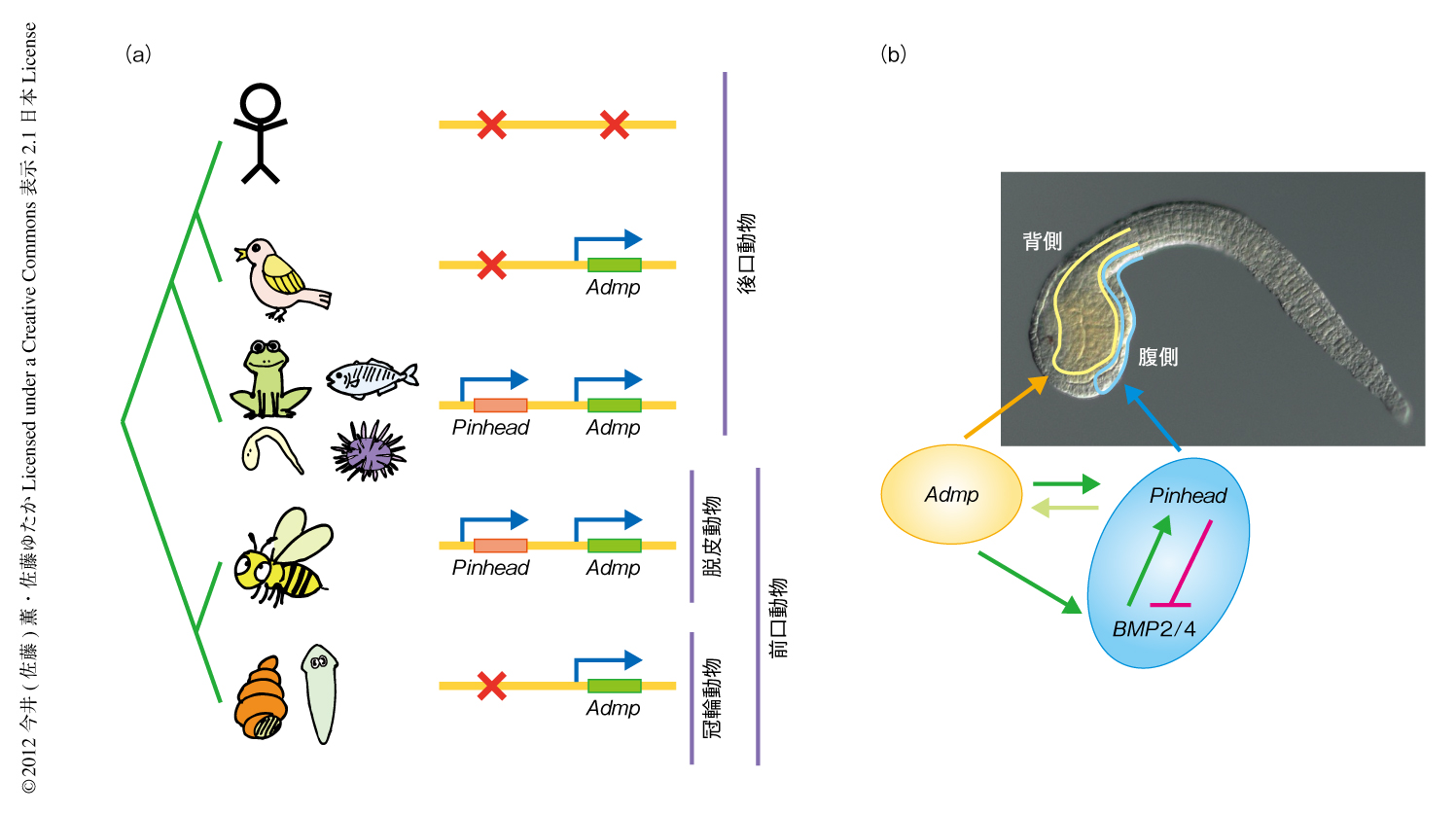
in situハイブリダイゼーションにより胚発生におけるPinhead遺伝子の発現を調べたところ,初期胚における一過的な発現にくわえ,神経胚期ののちに腹側の表皮細胞において発現していることが明らかになった.これまでの制御遺伝子の網羅的な発現解析により7),神経胚期ののちにはBMPアンタゴニストをコードするChordin遺伝子がもっとも背側の表皮細胞で発現し,それに隣接する背側から側方にかけての表皮細胞でAdmp遺伝子が発現し,さらにもっとも腹側の領域でBmp2/4遺伝子(脊椎動物のBMP2遺伝子とBMP4遺伝子のオーソログ)が発現することがわかっていた.この発現パターンは,たとえばカエルの原腸胚での背腹軸の決定におけるホモログ遺伝子の発現パターンを思わせるものであった.また,Bmp2/4遺伝子の発現領域はPinhead遺伝子の発現領域とオーバーラップするが,Admp遺伝子の発現領域とPinhead遺伝子の発現領域はオーバーラップしないこともわかった(図1b).
モルフォリノアンチセンスオリゴヌクレオチドを用いて遺伝子の機能を阻害したところ,Admp遺伝子およびBmp2/4遺伝子の機能を阻害した胚では腹側の領域の形成が阻害された(腹側マーカーであるNk4遺伝子の発現が消失または減少した)のに対し,Pinhead遺伝子の機能を阻害した胚では腹側の領域が拡大した(腹側マーカーであるNk4遺伝子の発現領域が拡大した).このことは,Admp遺伝子とBmp2/4遺伝子は腹側の形成に対し正の制御を行い,Pinhead遺伝子は負の制御を行なっていることを意味していた.さらに,これら3つの遺伝子は複雑な制御関係にあって互いの制御を行なっていた(図1b).
Pinhead遺伝子がAdmp遺伝子およびBmp2/4遺伝子に対し相反する機能をもつこと,Pinheadはシステインノット構造をもつことから,Pinheadが新規のBMPアンタゴニストである可能性が考えられた.そこで,そのことを免疫沈降により確認したところ,興味深いことに,in vivoにおいてPinheadはAdmpと強く結合するのに対し,Bmp2/4とはほとんど結合しなかった.したがって,PinheadはAdmpに特異的なアンタゴニストであると考えられた.
余談になるが,免疫沈降実験にはMycタグを結合したAdmp(または,Bmp2/4)とFlagタグを結合したPinheadとを利用した.これらのタグタンパク質は,その遺伝子をDll-b遺伝子の上流につないだプラスミドをエレクトロポレーション法により受精卵に導入することにより表皮細胞の全体において強制発現させた.ホヤではこうした手法により,培養細胞を用いなくとも比較的容易にin vivoにおける免疫沈降実験が可能である.
Admp遺伝子とPinhead遺伝子は互いに隣接してゲノムに位置し,それが進化的にも保存されていた.その理由を探り,また,Pinhead遺伝子による背腹軸の制御機構をさらにくわしく理解するため,Admp遺伝子およびPinhead遺伝子による遺伝子発現の制御機構を解析した(図2a).Pinhead遺伝子の上流からAdmp遺伝子まで約10 kbの領域を遺伝子クローニングし,Pinhead遺伝子をRFP遺伝子に,Admp遺伝子をGFP遺伝子に置換した.このDNAコンストラクトをエレクトロポレーション法により受精卵に導入すると,そこから発生する胚では尾芽胚期において,GFPが背側から側方にかけての領域,RFPが腹側の領域で発現し,本来のAdmp遺伝子およびPinhead遺伝子の発現パターンが再現された.
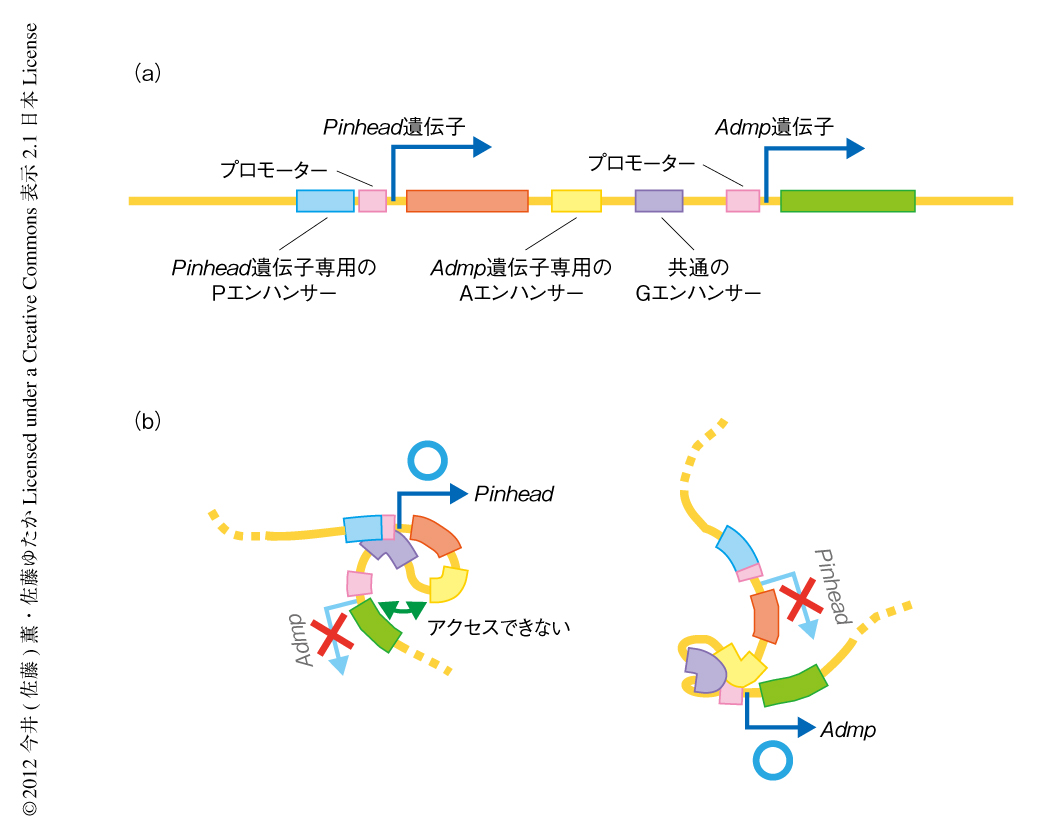
Pinhead遺伝子の上流には複数のSmad結合部位が存在した.SmadはBMPシグナル伝達系をうけ活性化する転写因子であり,また,さきに述べた解析によりPinhead遺伝子はAdmpおよびBMP2/4の制御下にあることがわかっていた.そこで,Pinhead遺伝子の上流のSmad結合部位に変異を導入したところ,Pinhead遺伝子の発現を模倣するはずのRFP遺伝子は発現しなかった.つまり,このSmad結合部位を含む領域はエンハンサーとして機能していた(Pエンハンサー).
Admp遺伝子の上流には未知のbHLH型転写因子が結合すると考えられる領域が存在し,Admp遺伝子のエンハンサーとして機能した(Aエンハンサー).このAエンハンサーとAdmp遺伝子とのあいだには,Pinhead遺伝子とAdmp遺伝子の両方を正に制御するエンハンサーが存在していた(Gエンハンサー).
興味深いことに,最初のPinhead遺伝子上流+ RFP遺伝子+ Admp遺伝子上流+ GFP遺伝子というDNAコンストラクトを,Pinhead遺伝子上流+ RFP遺伝子とAdmp遺伝子上流+ GFP遺伝子という2つのコンストラクトに分割してホヤの胚に導入すると,RFPは本来の腹側にのみ発現するものの,GFPは背側から側方にかけての本来の発現領域にくわえ腹側でも異所的に発現した.このことは,Pinhead遺伝子上流+ RFP遺伝子の領域がなんらかのかたちでGFP遺伝子の発現を制御し,通常の状態でのGFPの腹側における発現を抑制していることを意味した.しかし,RFP遺伝子を除きPinhead遺伝子上流を直接にAdmp遺伝子上流に結合したコンストラクトを導入したときにもGFPは腹側に発現したので,Pinhead遺伝子上流がAdmp遺伝子のエンハンサーとして直接に機能するとは考えにくかった.
以上の実験結果から,RFP遺伝子の転写そのものが下流にあるGFP遺伝子の転写を阻害していると考えられた.例は多くないが既知の分子機構からの推測として,RFP遺伝子のリードスルー転写産物(通常のポリA付加部位をこえて転写されたRNA)がAdmp遺伝子のエンハンサーの機能を阻害している可能性が考えられたが,この領域にはほとんどリードスルー転写産物は検出されなかった.そこで,この領域の三次元構造を3C(chromosome conformation capture,染色体コンホメーション捕捉)法8) により調べた.解析には内在性のPinhead遺伝子またはAdmp遺伝子の転写を実験的に亢進させた胚を用いた.その結果,Pinhead遺伝子の発現が亢進している状況では,Pinhead遺伝子のプロモーター領域およびPエンハンサーを含む領域が,AエンハンサーおよびGエンハンサーを含む領域と強く相互作用していた(図2b).つまり,Pinhead遺伝子が転写されているときにはPinhead遺伝子の上流とGエンハンサーとが相互作用しており,結果としてできるDNAのループに,Admp遺伝子の発現に必要なAエンハンサーが隔離されてしまっていると考えられた.AエンハンサーとGエンハンサーの位置を入れ替えた場合には,上流の遺伝子の転写が下流の遺伝子の転写を阻害することはなかった.
Pinhead遺伝子とAdmp遺伝子とがゲノムに並んで存在するという遺伝子の配置は,動物のゲノムにおいて広範囲に保存されていた.今回,公開されている種々の動物のゲノムを調べたところ,Admp遺伝子はこれまで考えられていたより広範囲の動物においてみつかった(図1a).また,Pinhead遺伝子は前口動物のうち冠輪動物にはみつからなかったが,脱皮動物にはみつかった(ただし,ショウジョウバエなど一部の動物ではAdmp遺伝子もPinhead遺伝子も存在しなかった).後口動物では,ヒトを含む一部の哺乳類でAdmp遺伝子が存在せず,Pinhead遺伝子は哺乳類と鳥類に存在しなかったものの,そのほかの後口動物にはAdmp遺伝子もPinhead遺伝子も存在していた.大事なのは,Pinhead遺伝子の存在するゲノムではつねにそのすぐ下流にAdmp遺伝子がコードされていたことである.
また,メダカでもホヤと同様の分子機構のはたらいていることが確認された.この遺伝子の並びはこれら遺伝子の機能に必要不可欠な役割をはたしており,それが進化において遺伝子の並びが高度に保存されてきた理由であろうと考えられた.通常,ゲノムにおける遺伝子の並びは進化の過程で比較的自由に変化すると考えられている.しかしながら,このPinhead-Admp遺伝子クラスターのように高度に保存された遺伝子クラスターは少数ながら存在している.今回,発見された分子機構が進化的な制約となりゲノムの進化に影響したものと考えられた.
今回,筆者らの解析したPinhead遺伝子は,タンパク質レベルおよび転写レベルの両方でAdmp遺伝子を負に制御していた.この二重の抑制機構は腹側の領域の境界をシャープに決める役割を担っているものと考えられた.この機構は広く動物界において保存されていると考えられることから,動物の発生において非常に基本的な役割をはたしているに違いない.哺乳類と鳥類ではPinhead-Admp遺伝子クラスターは存在していないが,これらの動物ではPinhead-Admp遺伝子クラスターの機能を代替するなんらかの新しい機構が生み出されたのかもしれない.Spemannの有名な結紮実験(イモリの胚を背腹で半分になるよう結紮すると,背側の半分にほぼ完全な胚が生じる)にみられるような両生類の胚の制御能力あるいは再生能力にAdmp遺伝子が重要な役割をはたすことが明らかになるなど9,10),Admp遺伝子の役割はこれまで考えられていたよりずっと重要なのかもしれない.その背景には,今回,発見された分子機構の存在があるだろう.逆に,われわれヒトは進化の過程において,なぜこのPinhead-Admp遺伝子クラスターを失うことができたのだろうか.この問いは,今回,提起されたひとつの課題である.
略歴:2003年 京都大学大学院理学研究科 修了,同年 同 研究員,2006年 米国California大学Berkeley校 研究員,2007年 京都大学大学院理学研究科 研究員,2008年 同 特任助教を経て,2012年より日本学術振興会 特別研究員(RPD).
研究テーマ:ホヤの発生生物学.
関心事:ゲノムレベルにおける発生の制御.
佐藤 ゆたか(Yutaka Satou)
京都大学大学院理学研究科 准教授.
研究室URL:http://ghost.zool.kyoto-u.ac.jp/bunshi/opening.htm
© 2012 今井 (佐藤) 薫・佐藤ゆたか Licensed under CC 表示 2.1 日本
(京都大学大学院理学研究科 生物科学専攻発生ゲノム科学研究室)
email:今井 (佐藤) 薫,佐藤ゆたか
DOI: 10.7875/first.author.2012.114
Cis-acting transcriptional repression establishes a sharp boundary in chordate embryos.
Kaoru S. Imai, Yutaka Daido, Takehiro G. Kusakabe, Yutaka Satou
Science, 337, 964-967 (2012)
要 約
動物胚における背側と腹側の領域の決定については,両生類の胚など脊椎動物の胚での研究によりBMPシグナル伝達系が大きな役割をはたしていることが明らかになっている.つまり,BMPシグナル伝達系は腹側の領域を誘導し,背側ではBMPシグナル伝達系は阻害される.ほかのBMPリガンドとは異なり,Admpは背側の領域で発現する.しかし,そののち腹側に輸送されて,そこでほかのBMPリガンドと同様に腹側の運命決定を行う.今回,PinheadはAdmpに特異的なアンタゴニストであり,脊索動物尾索類のカタユウレイボヤにおける表皮の腹側の領域の決定に必須であることが示された.Pinheadはこのタンパク質レベルでのAdmpの抑制にくわえ,Pinhead遺伝子の転写によりAdmp遺伝子の転写が直接的に抑制されるという二重の抑制機構をもっていた.この転写抑制はこの2つの遺伝子がゲノムにおいて隣接してコードされていることを利用していたが,この遺伝子の並びは動物のゲノムにおいて広く保存されており,この制御機構は幅広い動物において利用されているものと思われた.
はじめに
動物の発生は,受精卵というひとつの細胞にはじまり,それが細胞分裂をへてさまざまな組織や器官をつくりあげていく現象である.発生において組織や器官の分化は漸進的に進んでいく.背腹軸の決定はそのなかでも初期において必要な過程であり,のちの胚発生はそこで決定された軸を利用している.この背腹軸の決定の分子機構の研究は多くの発生学者を魅了してきたが,近年,背腹軸の決定に関与するタンパク質の作用機序がつぎつぎに明らかになってきている.非常に簡単にいえば,脊椎動物では腹側においてBMPシグナル伝達系がはたらき,背側においてBMPシグナル伝達系が抑制されることが必要である.興味深いことに,BMP様のタンパク質であるAdmp(anti-dorsalising morphogenetic protein)は作用すべき腹側ではなく背側において発現する.そののち,Admpは腹側に輸送され,そこでBMP4などと協調し腹側における発生プログラムを始動する1).
筆者らは,この分子機構をさらにくわしく解析するため,カタユウレイボヤ(Ciona intestinalis)を用いて実験を行った.ホヤはわれわれ脊椎動物と同じ脊索動物であり,基本的な体制を共有している.つまり,ホヤは幼生時において脊索動物に特徴的な“オタマジャクシ型”の形態をとる.このことは,この基本的な体制をつくるための発生の機構が脊索動物において共有されていることを強く示唆する.一方で,筆者らがこれまでに明らかにしてきた遺伝子ネットワークの研究などをもとにして2),ホヤの初期胚ではBMPシグナル伝達系に依存的に背腹軸を決定する機構が失われている可能性が示唆されてきた3).しかしながら今回,筆者らは,ホヤにおいても神経胚期ののちBMPシグナルに依存的に背腹軸が決定される機構が存在することを見い出し,その実験系を使い研究を進めた.ホヤはゲノムがコンパクトなうえ4)(ゲノムサイズは160 Mb,遺伝子数は約16,000),幼生も単純なので(幼生は少数の組織から構成され,その細胞数は約2600),実験的な解析が容易という利点がある5).
1.Pinhead遺伝子の同定
BMPシグナル伝達系は動物胚における背腹の決定に重要な役割をはたす.BMPシグナル伝達系の特徴として,多くのアンタゴニストにより制御されていることがあげられる.興味深いことに,BMPリガンドとChordinなどBMPアンタゴニストの一部は,システインノットとよばれる構造を共通にもつ.そこで,BMPシグナル伝達系を調べる過程でこの構造に注目し,このシステインノット構造をもつタンパク質をコードする遺伝子をホヤのゲノムにおいてすべて調べた.このなかにはもちろん,BMPリガンドを含むTGFβスーパーファミリータンパク質の遺伝子やChordinなどのBMPアンタゴニストをコードする遺伝子など既知のものが含まれていたが,そのほか機能未知の遺伝子がいくつか同定された.なかでも,アフリカツメガエルにおいてすでに同定されていたPinhead遺伝子6) のホモログは,つぎの2つの点で興味深かった.第1に,同定されたほかすべてはシステインノット構造を1つしかもたなかったのに対し,Pinheadは3つのシステインノットをもっていた.第2に,Pinhead遺伝子はゲノムにおいてBMPリガンドのひとつをコードするAdmp遺伝子に隣接してコードされており,のちに述べるように,その遺伝子の並びは進化的に保存されていた(図1a).そこで,このPinhead遺伝子の機能をさらにくわしく調べることにした.
2.Pinhead遺伝子の機能
in situハイブリダイゼーションにより胚発生におけるPinhead遺伝子の発現を調べたところ,初期胚における一過的な発現にくわえ,神経胚期ののちに腹側の表皮細胞において発現していることが明らかになった.これまでの制御遺伝子の網羅的な発現解析により7),神経胚期ののちにはBMPアンタゴニストをコードするChordin遺伝子がもっとも背側の表皮細胞で発現し,それに隣接する背側から側方にかけての表皮細胞でAdmp遺伝子が発現し,さらにもっとも腹側の領域でBmp2/4遺伝子(脊椎動物のBMP2遺伝子とBMP4遺伝子のオーソログ)が発現することがわかっていた.この発現パターンは,たとえばカエルの原腸胚での背腹軸の決定におけるホモログ遺伝子の発現パターンを思わせるものであった.また,Bmp2/4遺伝子の発現領域はPinhead遺伝子の発現領域とオーバーラップするが,Admp遺伝子の発現領域とPinhead遺伝子の発現領域はオーバーラップしないこともわかった(図1b).
モルフォリノアンチセンスオリゴヌクレオチドを用いて遺伝子の機能を阻害したところ,Admp遺伝子およびBmp2/4遺伝子の機能を阻害した胚では腹側の領域の形成が阻害された(腹側マーカーであるNk4遺伝子の発現が消失または減少した)のに対し,Pinhead遺伝子の機能を阻害した胚では腹側の領域が拡大した(腹側マーカーであるNk4遺伝子の発現領域が拡大した).このことは,Admp遺伝子とBmp2/4遺伝子は腹側の形成に対し正の制御を行い,Pinhead遺伝子は負の制御を行なっていることを意味していた.さらに,これら3つの遺伝子は複雑な制御関係にあって互いの制御を行なっていた(図1b).
Pinhead遺伝子がAdmp遺伝子およびBmp2/4遺伝子に対し相反する機能をもつこと,Pinheadはシステインノット構造をもつことから,Pinheadが新規のBMPアンタゴニストである可能性が考えられた.そこで,そのことを免疫沈降により確認したところ,興味深いことに,in vivoにおいてPinheadはAdmpと強く結合するのに対し,Bmp2/4とはほとんど結合しなかった.したがって,PinheadはAdmpに特異的なアンタゴニストであると考えられた.
余談になるが,免疫沈降実験にはMycタグを結合したAdmp(または,Bmp2/4)とFlagタグを結合したPinheadとを利用した.これらのタグタンパク質は,その遺伝子をDll-b遺伝子の上流につないだプラスミドをエレクトロポレーション法により受精卵に導入することにより表皮細胞の全体において強制発現させた.ホヤではこうした手法により,培養細胞を用いなくとも比較的容易にin vivoにおける免疫沈降実験が可能である.
3.Admp遺伝子およびPinhead遺伝子による遺伝子発現の制御機構
Admp遺伝子とPinhead遺伝子は互いに隣接してゲノムに位置し,それが進化的にも保存されていた.その理由を探り,また,Pinhead遺伝子による背腹軸の制御機構をさらにくわしく理解するため,Admp遺伝子およびPinhead遺伝子による遺伝子発現の制御機構を解析した(図2a).Pinhead遺伝子の上流からAdmp遺伝子まで約10 kbの領域を遺伝子クローニングし,Pinhead遺伝子をRFP遺伝子に,Admp遺伝子をGFP遺伝子に置換した.このDNAコンストラクトをエレクトロポレーション法により受精卵に導入すると,そこから発生する胚では尾芽胚期において,GFPが背側から側方にかけての領域,RFPが腹側の領域で発現し,本来のAdmp遺伝子およびPinhead遺伝子の発現パターンが再現された.
Pinhead遺伝子の上流には複数のSmad結合部位が存在した.SmadはBMPシグナル伝達系をうけ活性化する転写因子であり,また,さきに述べた解析によりPinhead遺伝子はAdmpおよびBMP2/4の制御下にあることがわかっていた.そこで,Pinhead遺伝子の上流のSmad結合部位に変異を導入したところ,Pinhead遺伝子の発現を模倣するはずのRFP遺伝子は発現しなかった.つまり,このSmad結合部位を含む領域はエンハンサーとして機能していた(Pエンハンサー).
Admp遺伝子の上流には未知のbHLH型転写因子が結合すると考えられる領域が存在し,Admp遺伝子のエンハンサーとして機能した(Aエンハンサー).このAエンハンサーとAdmp遺伝子とのあいだには,Pinhead遺伝子とAdmp遺伝子の両方を正に制御するエンハンサーが存在していた(Gエンハンサー).
興味深いことに,最初のPinhead遺伝子上流+ RFP遺伝子+ Admp遺伝子上流+ GFP遺伝子というDNAコンストラクトを,Pinhead遺伝子上流+ RFP遺伝子とAdmp遺伝子上流+ GFP遺伝子という2つのコンストラクトに分割してホヤの胚に導入すると,RFPは本来の腹側にのみ発現するものの,GFPは背側から側方にかけての本来の発現領域にくわえ腹側でも異所的に発現した.このことは,Pinhead遺伝子上流+ RFP遺伝子の領域がなんらかのかたちでGFP遺伝子の発現を制御し,通常の状態でのGFPの腹側における発現を抑制していることを意味した.しかし,RFP遺伝子を除きPinhead遺伝子上流を直接にAdmp遺伝子上流に結合したコンストラクトを導入したときにもGFPは腹側に発現したので,Pinhead遺伝子上流がAdmp遺伝子のエンハンサーとして直接に機能するとは考えにくかった.
以上の実験結果から,RFP遺伝子の転写そのものが下流にあるGFP遺伝子の転写を阻害していると考えられた.例は多くないが既知の分子機構からの推測として,RFP遺伝子のリードスルー転写産物(通常のポリA付加部位をこえて転写されたRNA)がAdmp遺伝子のエンハンサーの機能を阻害している可能性が考えられたが,この領域にはほとんどリードスルー転写産物は検出されなかった.そこで,この領域の三次元構造を3C(chromosome conformation capture,染色体コンホメーション捕捉)法8) により調べた.解析には内在性のPinhead遺伝子またはAdmp遺伝子の転写を実験的に亢進させた胚を用いた.その結果,Pinhead遺伝子の発現が亢進している状況では,Pinhead遺伝子のプロモーター領域およびPエンハンサーを含む領域が,AエンハンサーおよびGエンハンサーを含む領域と強く相互作用していた(図2b).つまり,Pinhead遺伝子が転写されているときにはPinhead遺伝子の上流とGエンハンサーとが相互作用しており,結果としてできるDNAのループに,Admp遺伝子の発現に必要なAエンハンサーが隔離されてしまっていると考えられた.AエンハンサーとGエンハンサーの位置を入れ替えた場合には,上流の遺伝子の転写が下流の遺伝子の転写を阻害することはなかった.
4.Pinhead-Admp遺伝子クラスター
Pinhead遺伝子とAdmp遺伝子とがゲノムに並んで存在するという遺伝子の配置は,動物のゲノムにおいて広範囲に保存されていた.今回,公開されている種々の動物のゲノムを調べたところ,Admp遺伝子はこれまで考えられていたより広範囲の動物においてみつかった(図1a).また,Pinhead遺伝子は前口動物のうち冠輪動物にはみつからなかったが,脱皮動物にはみつかった(ただし,ショウジョウバエなど一部の動物ではAdmp遺伝子もPinhead遺伝子も存在しなかった).後口動物では,ヒトを含む一部の哺乳類でAdmp遺伝子が存在せず,Pinhead遺伝子は哺乳類と鳥類に存在しなかったものの,そのほかの後口動物にはAdmp遺伝子もPinhead遺伝子も存在していた.大事なのは,Pinhead遺伝子の存在するゲノムではつねにそのすぐ下流にAdmp遺伝子がコードされていたことである.
また,メダカでもホヤと同様の分子機構のはたらいていることが確認された.この遺伝子の並びはこれら遺伝子の機能に必要不可欠な役割をはたしており,それが進化において遺伝子の並びが高度に保存されてきた理由であろうと考えられた.通常,ゲノムにおける遺伝子の並びは進化の過程で比較的自由に変化すると考えられている.しかしながら,このPinhead-Admp遺伝子クラスターのように高度に保存された遺伝子クラスターは少数ながら存在している.今回,発見された分子機構が進化的な制約となりゲノムの進化に影響したものと考えられた.
おわりに
今回,筆者らの解析したPinhead遺伝子は,タンパク質レベルおよび転写レベルの両方でAdmp遺伝子を負に制御していた.この二重の抑制機構は腹側の領域の境界をシャープに決める役割を担っているものと考えられた.この機構は広く動物界において保存されていると考えられることから,動物の発生において非常に基本的な役割をはたしているに違いない.哺乳類と鳥類ではPinhead-Admp遺伝子クラスターは存在していないが,これらの動物ではPinhead-Admp遺伝子クラスターの機能を代替するなんらかの新しい機構が生み出されたのかもしれない.Spemannの有名な結紮実験(イモリの胚を背腹で半分になるよう結紮すると,背側の半分にほぼ完全な胚が生じる)にみられるような両生類の胚の制御能力あるいは再生能力にAdmp遺伝子が重要な役割をはたすことが明らかになるなど9,10),Admp遺伝子の役割はこれまで考えられていたよりずっと重要なのかもしれない.その背景には,今回,発見された分子機構の存在があるだろう.逆に,われわれヒトは進化の過程において,なぜこのPinhead-Admp遺伝子クラスターを失うことができたのだろうか.この問いは,今回,提起されたひとつの課題である.
文 献
- De Robertis, E. M.: Spemann's organizer and the self-regulation of embryonic fields. Mech. Dev., 126, 925-941 (2009)[PubMed]
- Imai, K. S., Levine, M., Satoh, N. et al.: Regulatory blueprint for a chordate embryo. Science, 312, 1183-1187 (2006)[PubMed]
- Lemaire, P.: Developmental biology. How many ways to make a chordate? Science, 312, 1145-1146 (2006)[PubMed]
- Dehal, P., Satou, Y., Campbell, R. K. et al.: The draft genome of Ciona intestinalis: insights into chordate and vertebrate origins. Science, 298, 2157-2167 (2002)[PubMed]
- Satoh, N.: The ascidian tadpole larva: comparative molecular development and genomics. Nat. Rev. Genet., 4, 285-295 (2003)[PubMed]
- Kenwrick, S., Amaya, E. & Papalopulu, N.: Pilot morpholino screen in Xenopus tropicalis identifies a novel gene involved in head development. Dev. Dyn., 229, 289-299 (2004)[PubMed]
- Imai, K. S., Hino, K., Yagi, K. et al.: Gene expression profiles of transcription factors and signaling molecules in the ascidian embryo: towards a comprehensive understanding of gene networks. Development, 131, 4047-4058 (2004)[PubMed]
- Dekker, J., Rippe, K., Dekker, M. et al.: Capturing chromosome conformation. Science, 295, 1306-1311 (2002)[PubMed]
- Reversade, B. & De Robertis, E. M.: Regulation of ADMP and BMP2/4/7 at opposite embryonic poles generates a self-regulating morphogenetic field. Cell, 123, 1147-1160 (2005)[PubMed]
- Ben-Zvi, D., Shilo, B. Z., Fainsod, A. et al.: Scaling of the BMP activation gradient in Xenopus embryos. Nature, 453, 1205-1211 (2008)[PubMed]
著者プロフィール
略歴:2003年 京都大学大学院理学研究科 修了,同年 同 研究員,2006年 米国California大学Berkeley校 研究員,2007年 京都大学大学院理学研究科 研究員,2008年 同 特任助教を経て,2012年より日本学術振興会 特別研究員(RPD).
研究テーマ:ホヤの発生生物学.
関心事:ゲノムレベルにおける発生の制御.
佐藤 ゆたか(Yutaka Satou)
京都大学大学院理学研究科 准教授.
研究室URL:http://ghost.zool.kyoto-u.ac.jp/bunshi/opening.htm
© 2012 今井 (佐藤) 薫・佐藤ゆたか Licensed under CC 表示 2.1 日本
