熱性けいれんののちの興奮性GABAA受容体シグナル伝達により異所性の顆粒細胞の出現とてんかんの発症が誘導される
小山 隆太
(東京大学大学院薬学系研究科 薬品作用学教室)
email:小山隆太
DOI: 10.7875/first.author.2012.101
GABAergic excitation after febrile seizures induces ectopic granule cells and adult epilepsy.
Ryuta Koyama, Kentaro Tao, Takuya Sasaki, Junya Ichikawa, Daisuke Miyamoto, Rieko Muramatsu, Norio Matsuki, Yuji Ikegaya
Nature Medicine, 18, 1271-1278 (2012)
側頭葉てんかんの患者の海馬歯状回では,しばしば顆粒細胞の異常な局在が観察される.この論文では,側頭葉てんかんの発症への関与が示唆される複雑型熱性けいれんのラットモデルを用いて,新生仔期に生じた顆粒細胞の異常な移動が,成体期まで持続する異所性の顆粒細胞の出現につながることを報告した.熱性けいれんは新生仔期に生じた顆粒細胞においてGABAA受容体の発現の上昇を誘導した.また,このことによる興奮性GABA入力の増強は顆粒細胞の移動方向の逆転をひき起こした.顆粒細胞の移動に対する興奮性GABA入力の影響は,GABAの興奮作用を制御するNa+K+2Cl-共輸送体NKCC1をRNAi法によりノックダウンすることにより阻止された.また,熱性けいれんののちNKCC1をその阻害薬により抑制すると,成体期における異所性の顆粒細胞の出現,辺縁系けいれん発作の感受性の上昇,および,てんかんの発症が阻止された.この研究は,興奮性GABAA受容体シグナル伝達のこれまで明らかにされていなかった病因的な役割を明らかにした.さらに,NKCC1の抑制が熱性けいれんののちの異所性の顆粒細胞の出現,および,てんかんの発症を防御する新規の治療法となる可能性を示した.
皮質の発達期におけるニューロンの移動の制御不全は,ニューロンの異常な局在につながり,てんかんの発症の可能性を上昇させる1).ニューロンの異常な局在は各種のてんかん動物モデルにおいて自発発作や発作閾値の低下と関連することが示唆されている.なかでも,皮質下帯状異所性灰白質のモデルラットにおいてニューロンの異常な局在の防止がけいれん発作の感受性を低下させたという報告は,ニューロンの異常な局在とてんかんの発症との関連を検証するうえで重要なものである2).
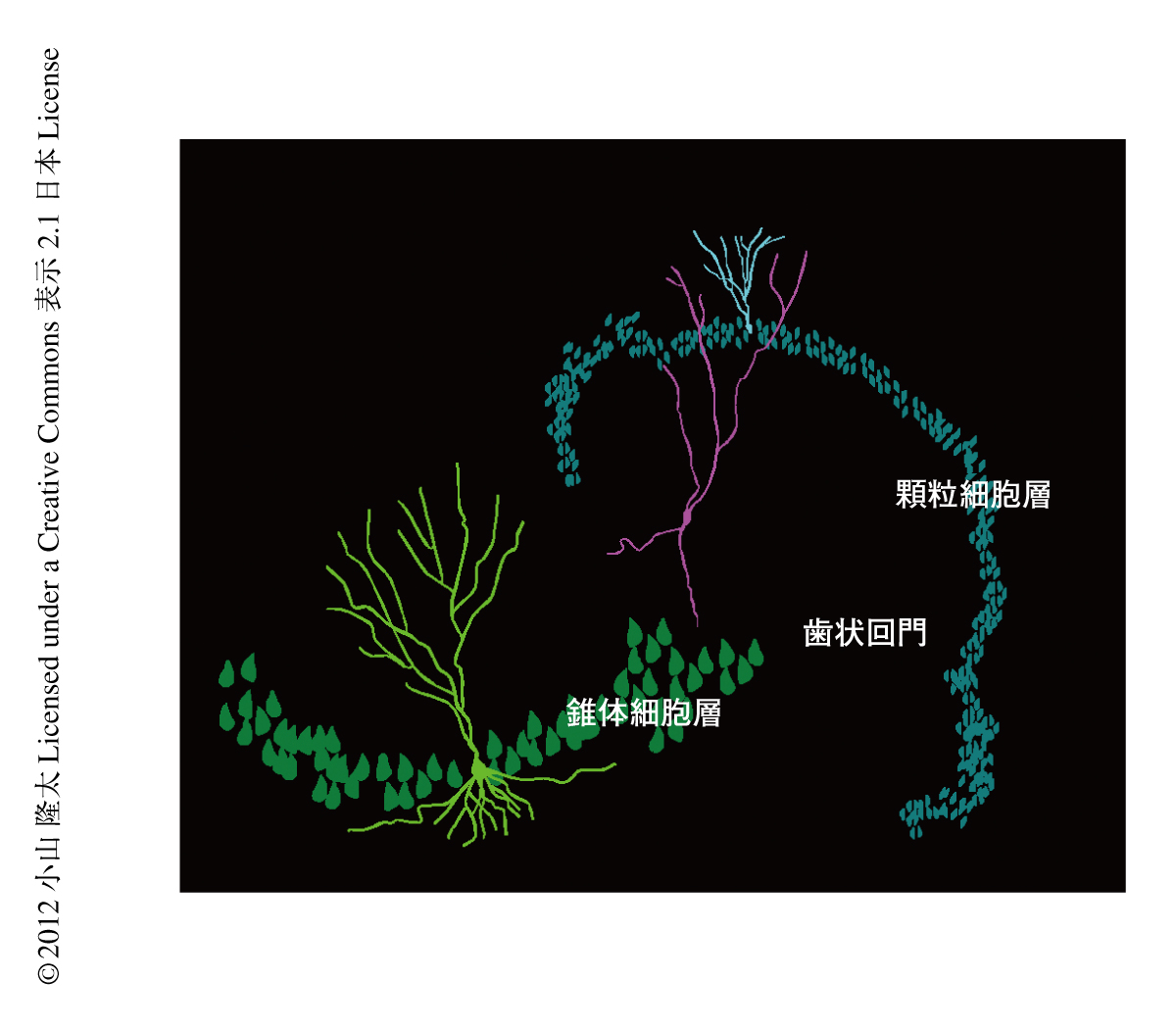
側頭葉てんかんの患者およびその動物モデルでは,海馬歯状回に存在する顆粒細胞について2種類の異常な局在が確認されている.ひとつは,顆粒細胞が細胞層の付近に分散して存在する“顆粒細胞の分散”であり3),もうひとつは,顆粒細胞が歯状回門に異所的に存在する“異所性の顆粒細胞の出現”である4).とくに,動物モデルにおいて頻繁に確認される異所性の顆粒細胞は,歯状回に興奮回路を付加することにより海馬に存在するほかのニューロンと同期した発射を行うため,てんかん原生の獲得に関与する可能性が示唆されている4)(図1).
熱性けいれんは生後6カ月から5歳までの小児にもっとも頻繁に起こるけいれんである.その発症率には地理的な差異があるが,世界中のこの年齢の小児の約2~14%に生じている5).多くの場合,熱性けいれんは良性だが,全体の30~40%は発作時間が15分をこえる複雑型熱性けいれんであり,成人の側頭葉てんかんの患者の30~70%が幼少期に複雑型熱性けいれんを経験しているという報告がある6,7).
筆者らは,複雑型熱性けいれんのモデルラット8) を利用し,熱性けいれんが異所性の顆粒細胞の出現を誘導する可能性およびその分子機構,そして,てんかんの発症への関与を検証した.
熱性けいれんが新生児期に産生された顆粒細胞の局在に影響する可能性を,熱性けいれんモデルラットを利用して検証した.熱性けいれんは生後11日齢のラットを高温の環境に曝露することにより誘導した8).まず,生後5日齢のラットに細胞周期S期のマーカーであるBrdUを皮下投与し,生後11日齢において熱性けいれんを誘導した.そののち,生後18日齢または生後60日齢において還流固定し,免疫染色法により顆粒細胞のマーカーであるProx1とBrdUの両方のマーカーについて陽性な細胞,すなわち,新生した顆粒細胞の局在を調べた.その結果,熱性けいれんを経験したラットには顆粒細胞層までの移動を完了せず歯状回門に異所的に存在する新生した顆粒細胞が,対照のラットと比較して有意に多く観察された.さらに,これらの顆粒細胞の形態的な特徴を検証するため,GFPの遺伝子をコードしたレトロウイルスを生後5日齢のラットの歯状回に注入し,そののち生後11日齢において熱性けいれんを誘導した.その結果,生後60日齢において双極性の樹状突起と顆粒細胞層にむけて投射する軸索をもつ異所性の顆粒細胞の形態が観察された.以上の結果は,熱性けいれんにより正常な移動の阻害された新生した顆粒細胞が,将来的に異所性の顆粒細胞となる可能性を示唆した.
神経伝達物質であるGABA(γ-aminobutyric acid,γ-アミノ酪酸)は皮質の形成期におけるニューロンの移動の制御に必須である.そこで,新生した顆粒細胞の移動においてGABAA受容体シグナル伝達が関与する可能性を,GABAA受容体の阻害薬ピクロトキシン,あるいは,作用薬フェノバルビタールを,熱性けいれんの誘導ののち腹腔内投与することにより検証した.その結果,ピクロトキシンは熱性けいれんによる異所性の顆粒細胞の出現を抑制した一方,フェノバルビタールは異所性の顆粒細胞の数を増加させた.GABAA受容体シグナル伝達が新生した顆粒細胞の移動を制御する可能性をより詳細に検証するため,海馬の切片の培養系を利用して薬理学的な検証を行った.海馬の切片は生後5日齢においてBrdUを投与したラットから生後6日齢にて作製し,5日間の培養を行った.培地に各種の薬物を添加して新生した顆粒細胞の局在への影響を調べたところ,GABAA受容体の阻害薬ビククリンは新生した顆粒細胞の顆粒細胞層への局在を促進させ,その作用薬ムシモールは異所性の顆粒細胞の出現を促進させた.また,同じく神経伝達物質であるグルタミン酸の受容体を阻害しても,対照と比較して有意な影響は観察されなかった.以上の結果は,GABAA受容体シグナル伝達の活性化は新生した顆粒細胞の顆粒細胞層への移動を抑制する可能性を示唆した.
さらに,全身にGFPを発現するGFPトランスジェニックラットに由来する歯状回門の切片を野生型ラットに由来する海馬の切片に挿入して培養する新規の共培養法を確立し,歯状回門から顆粒細胞層へと移動する新生した顆粒細胞をGFPにより可視化することに成功した.この系において,電気生理学的な手法を用いて移動している顆粒細胞の機能的な特徴を調べたところ,これらの顆粒細胞は移動しているとき興奮性のGABAA受容体シグナル伝達をうける一方,グルタミン酸受容体シグナル伝達はうけていないことが明らかになった.
以上の結果から,熱性けいれんを経験したラットの歯状回では興奮性GABAA受容体シグナル伝達の過剰な活性化による新生した顆粒細胞の移動の異常が生じ,最終的に異所性の顆粒細胞が出現している可能性が示唆された.まず,新生した顆粒細胞が移動する環境である歯状回門においてGABAの量が上昇している可能性を免疫染色法により検証したが,対照ラットおよび熱性けいれんラットとのあいだに有意な差は確認されなかった.この結果は,熱性けいれんが移動する環境(歯状回門)の側ではなく,移動する顆粒細胞の側になんらかの影響を及ぼした可能性を示唆した.この可能性を検証するため,さきに述べた培養切片の共培養法により,熱性けいれんを誘導したGFPトランスジェニックラットより作製した歯状回門の切片を野生型ラットの海馬の切片に挿入して培養し,タイムラプスイメージング法により観察した.その結果,GFP陽性の顆粒細胞は本来の顆粒細胞層の方向とは逆の方向に移動するようすが頻繁にみられた.さらに,このような異常な移動はGABAA受容体の阻害薬ビククリンの処理により阻止されたため,移動している顆粒細胞において興奮性GABAA受容体シグナル伝達の関与していることが明らかになった.
これらの現象をより詳細に解析するため,さらに,熱性けいれんラットに由来する歯状回門の移植片の培養法を確立した.これは,熱性けいれん誘導の1日後(生後12日齢)に作製した歯状回門の切片をリジンおよびラミニンでコーティングしたガラスのうえで培養するものである.この時期の歯状回門には多数の幼若な顆粒細胞が存在するため,移植片より外部へと遊走する顆粒細胞を観察することが可能となった.この系において,免疫染色法およびイムノブロット法により熱性けいれんを経験したラットに由来する顆粒細胞の細胞膜に存在するGABAA受容体(のβサブユニット)の発現量が,対照のラットと比較して有意に上昇していることが明らかになった.また,このGABAA受容体の発現の上昇はニューロンの移動に必須な構造である先導突起の成長円錐において確認されたため,遊走した顆粒細胞の成長円錐にて局所的にGABAA受容体の作用薬ムシモールを処理したところ,対照のラットに由来する顆粒細胞では観察されない移動の停止や逆走が誘導された.以上の結果は,熱性けいれんののちの興奮性GABAA受容体の入力による顆粒細胞の移動の異常の原因は,移動する環境の側ではなく,移動する顆粒細胞の側における変化であることを明らかにした.
幼若なニューロンにおいてGABAが興奮性にはたらく理由のひとつは,Na+K+2Cl-共輸送体NKCC1の発現が成熟したニューロンよりも高く,ニューロンにおけるCl-濃度が高く保たれるためである9).そこで,新生した顆粒細胞の移動における興奮性GABAA受容体シグナル伝達の影響がNKCC1により担われている可能性を,熱性けいれんラットに由来する歯状回門の移植片の培養系を利用して検証した.この系において,GABAA受容体の作用薬ムシモールの処理により顆粒細胞の移動距離は減少するが,この現象は培地にNKCC1の阻害薬ブメタニドを処理すること,または,siRNA法を用いて顆粒細胞においてNKCC1をノックダウンすることにより阻止された.
最後に,熱性けいれんによる異所性の顆粒細胞の出現と将来のてんかんの発症との関連を検証した.その結果,熱性けいれんを経験したラットは成体期(11~13週齢)においてピロカルピン誘導性のけいれん発作への感受性の高いことが明らかになった.さらに重要なことに,熱性けいれんを経験したラット16匹のうちの半数では辺縁系に由来する自発発作が確認され,その発作頻度と異所性の顆粒細胞の数とのあいだに正の相関性が確認された.また,熱性けいれんを経験したラットの海馬ではニューロンの過剰な発射の起こっていることをin vivoにおける電気生理学的な手法により明らかにした.一連のin vitroにおける実験により,興奮性GABAA受容体シグナル伝達による異所性の顆粒細胞の出現の根本的な原因はNKCC1であることがつきとめられたため,NKCC1の阻害薬ブメタニドを熱性けいれんの誘導ののち1週間にわたり処理した.その結果,成体期における異所性の顆粒細胞の出現,発作閾値の低下,自発発作の発症,そして,海馬におけるニューロンの過剰な発射が抑制された.
複雑型熱性けいれんの発達期の神経回路の形成にあたえる影響,および,将来的なてんかんの発症への関与は明らかではなかった10).この研究では,ラットにおいて複雑型熱性けいれんが興奮性GABAA受容体シグナル伝達を介し異所性の顆粒細胞の出現を誘導することを明らかにした(図2).さらに,異所性の顆粒細胞とてんかんの発症とのあいだには正の相関性が確認された.また,複雑型熱性けいれんののちのブメタニドの投与(NKCC1の阻害)が,将来のてんかんの発症を防止するための新規の治療法となりうる可能性を示した.

略歴:2006年 東京大学大学院薬学系研究科 修了,同年 同 寄付講座教員を経て,2007年より同 助教.2010年より米国Harvard Medical Schoolに留学中.
研究テーマ:発達期および病態の脳における神経回路の形成機構.
抱負:各種の脳疾患における神経回路の形成の変化をオリジナリティーあふれる培養系に再現し,その細胞生物学的および分子生物学的な機構を明らかにするとともに,将来的な創薬をめざしたい.
© 2012 小山 隆太 Licensed under CC 表示 2.1 日本
(東京大学大学院薬学系研究科 薬品作用学教室)
email:小山隆太
DOI: 10.7875/first.author.2012.101
GABAergic excitation after febrile seizures induces ectopic granule cells and adult epilepsy.
Ryuta Koyama, Kentaro Tao, Takuya Sasaki, Junya Ichikawa, Daisuke Miyamoto, Rieko Muramatsu, Norio Matsuki, Yuji Ikegaya
Nature Medicine, 18, 1271-1278 (2012)
要 約
側頭葉てんかんの患者の海馬歯状回では,しばしば顆粒細胞の異常な局在が観察される.この論文では,側頭葉てんかんの発症への関与が示唆される複雑型熱性けいれんのラットモデルを用いて,新生仔期に生じた顆粒細胞の異常な移動が,成体期まで持続する異所性の顆粒細胞の出現につながることを報告した.熱性けいれんは新生仔期に生じた顆粒細胞においてGABAA受容体の発現の上昇を誘導した.また,このことによる興奮性GABA入力の増強は顆粒細胞の移動方向の逆転をひき起こした.顆粒細胞の移動に対する興奮性GABA入力の影響は,GABAの興奮作用を制御するNa+K+2Cl-共輸送体NKCC1をRNAi法によりノックダウンすることにより阻止された.また,熱性けいれんののちNKCC1をその阻害薬により抑制すると,成体期における異所性の顆粒細胞の出現,辺縁系けいれん発作の感受性の上昇,および,てんかんの発症が阻止された.この研究は,興奮性GABAA受容体シグナル伝達のこれまで明らかにされていなかった病因的な役割を明らかにした.さらに,NKCC1の抑制が熱性けいれんののちの異所性の顆粒細胞の出現,および,てんかんの発症を防御する新規の治療法となる可能性を示した.
はじめに
皮質の発達期におけるニューロンの移動の制御不全は,ニューロンの異常な局在につながり,てんかんの発症の可能性を上昇させる1).ニューロンの異常な局在は各種のてんかん動物モデルにおいて自発発作や発作閾値の低下と関連することが示唆されている.なかでも,皮質下帯状異所性灰白質のモデルラットにおいてニューロンの異常な局在の防止がけいれん発作の感受性を低下させたという報告は,ニューロンの異常な局在とてんかんの発症との関連を検証するうえで重要なものである2).
側頭葉てんかんの患者およびその動物モデルでは,海馬歯状回に存在する顆粒細胞について2種類の異常な局在が確認されている.ひとつは,顆粒細胞が細胞層の付近に分散して存在する“顆粒細胞の分散”であり3),もうひとつは,顆粒細胞が歯状回門に異所的に存在する“異所性の顆粒細胞の出現”である4).とくに,動物モデルにおいて頻繁に確認される異所性の顆粒細胞は,歯状回に興奮回路を付加することにより海馬に存在するほかのニューロンと同期した発射を行うため,てんかん原生の獲得に関与する可能性が示唆されている4)(図1).
熱性けいれんは生後6カ月から5歳までの小児にもっとも頻繁に起こるけいれんである.その発症率には地理的な差異があるが,世界中のこの年齢の小児の約2~14%に生じている5).多くの場合,熱性けいれんは良性だが,全体の30~40%は発作時間が15分をこえる複雑型熱性けいれんであり,成人の側頭葉てんかんの患者の30~70%が幼少期に複雑型熱性けいれんを経験しているという報告がある6,7).
筆者らは,複雑型熱性けいれんのモデルラット8) を利用し,熱性けいれんが異所性の顆粒細胞の出現を誘導する可能性およびその分子機構,そして,てんかんの発症への関与を検証した.
1.熱性けいれんにより異所性の顆粒細胞の出現が誘導される
熱性けいれんが新生児期に産生された顆粒細胞の局在に影響する可能性を,熱性けいれんモデルラットを利用して検証した.熱性けいれんは生後11日齢のラットを高温の環境に曝露することにより誘導した8).まず,生後5日齢のラットに細胞周期S期のマーカーであるBrdUを皮下投与し,生後11日齢において熱性けいれんを誘導した.そののち,生後18日齢または生後60日齢において還流固定し,免疫染色法により顆粒細胞のマーカーであるProx1とBrdUの両方のマーカーについて陽性な細胞,すなわち,新生した顆粒細胞の局在を調べた.その結果,熱性けいれんを経験したラットには顆粒細胞層までの移動を完了せず歯状回門に異所的に存在する新生した顆粒細胞が,対照のラットと比較して有意に多く観察された.さらに,これらの顆粒細胞の形態的な特徴を検証するため,GFPの遺伝子をコードしたレトロウイルスを生後5日齢のラットの歯状回に注入し,そののち生後11日齢において熱性けいれんを誘導した.その結果,生後60日齢において双極性の樹状突起と顆粒細胞層にむけて投射する軸索をもつ異所性の顆粒細胞の形態が観察された.以上の結果は,熱性けいれんにより正常な移動の阻害された新生した顆粒細胞が,将来的に異所性の顆粒細胞となる可能性を示唆した.
2.GABAA受容体シグナル伝達は新生した顆粒細胞の移動および局在を制御する
神経伝達物質であるGABA(γ-aminobutyric acid,γ-アミノ酪酸)は皮質の形成期におけるニューロンの移動の制御に必須である.そこで,新生した顆粒細胞の移動においてGABAA受容体シグナル伝達が関与する可能性を,GABAA受容体の阻害薬ピクロトキシン,あるいは,作用薬フェノバルビタールを,熱性けいれんの誘導ののち腹腔内投与することにより検証した.その結果,ピクロトキシンは熱性けいれんによる異所性の顆粒細胞の出現を抑制した一方,フェノバルビタールは異所性の顆粒細胞の数を増加させた.GABAA受容体シグナル伝達が新生した顆粒細胞の移動を制御する可能性をより詳細に検証するため,海馬の切片の培養系を利用して薬理学的な検証を行った.海馬の切片は生後5日齢においてBrdUを投与したラットから生後6日齢にて作製し,5日間の培養を行った.培地に各種の薬物を添加して新生した顆粒細胞の局在への影響を調べたところ,GABAA受容体の阻害薬ビククリンは新生した顆粒細胞の顆粒細胞層への局在を促進させ,その作用薬ムシモールは異所性の顆粒細胞の出現を促進させた.また,同じく神経伝達物質であるグルタミン酸の受容体を阻害しても,対照と比較して有意な影響は観察されなかった.以上の結果は,GABAA受容体シグナル伝達の活性化は新生した顆粒細胞の顆粒細胞層への移動を抑制する可能性を示唆した.
さらに,全身にGFPを発現するGFPトランスジェニックラットに由来する歯状回門の切片を野生型ラットに由来する海馬の切片に挿入して培養する新規の共培養法を確立し,歯状回門から顆粒細胞層へと移動する新生した顆粒細胞をGFPにより可視化することに成功した.この系において,電気生理学的な手法を用いて移動している顆粒細胞の機能的な特徴を調べたところ,これらの顆粒細胞は移動しているとき興奮性のGABAA受容体シグナル伝達をうける一方,グルタミン酸受容体シグナル伝達はうけていないことが明らかになった.
3.熱性けいれんの影響をうけた顆粒細胞は異常な移動を行う
以上の結果から,熱性けいれんを経験したラットの歯状回では興奮性GABAA受容体シグナル伝達の過剰な活性化による新生した顆粒細胞の移動の異常が生じ,最終的に異所性の顆粒細胞が出現している可能性が示唆された.まず,新生した顆粒細胞が移動する環境である歯状回門においてGABAの量が上昇している可能性を免疫染色法により検証したが,対照ラットおよび熱性けいれんラットとのあいだに有意な差は確認されなかった.この結果は,熱性けいれんが移動する環境(歯状回門)の側ではなく,移動する顆粒細胞の側になんらかの影響を及ぼした可能性を示唆した.この可能性を検証するため,さきに述べた培養切片の共培養法により,熱性けいれんを誘導したGFPトランスジェニックラットより作製した歯状回門の切片を野生型ラットの海馬の切片に挿入して培養し,タイムラプスイメージング法により観察した.その結果,GFP陽性の顆粒細胞は本来の顆粒細胞層の方向とは逆の方向に移動するようすが頻繁にみられた.さらに,このような異常な移動はGABAA受容体の阻害薬ビククリンの処理により阻止されたため,移動している顆粒細胞において興奮性GABAA受容体シグナル伝達の関与していることが明らかになった.
これらの現象をより詳細に解析するため,さらに,熱性けいれんラットに由来する歯状回門の移植片の培養法を確立した.これは,熱性けいれん誘導の1日後(生後12日齢)に作製した歯状回門の切片をリジンおよびラミニンでコーティングしたガラスのうえで培養するものである.この時期の歯状回門には多数の幼若な顆粒細胞が存在するため,移植片より外部へと遊走する顆粒細胞を観察することが可能となった.この系において,免疫染色法およびイムノブロット法により熱性けいれんを経験したラットに由来する顆粒細胞の細胞膜に存在するGABAA受容体(のβサブユニット)の発現量が,対照のラットと比較して有意に上昇していることが明らかになった.また,このGABAA受容体の発現の上昇はニューロンの移動に必須な構造である先導突起の成長円錐において確認されたため,遊走した顆粒細胞の成長円錐にて局所的にGABAA受容体の作用薬ムシモールを処理したところ,対照のラットに由来する顆粒細胞では観察されない移動の停止や逆走が誘導された.以上の結果は,熱性けいれんののちの興奮性GABAA受容体の入力による顆粒細胞の移動の異常の原因は,移動する環境の側ではなく,移動する顆粒細胞の側における変化であることを明らかにした.
幼若なニューロンにおいてGABAが興奮性にはたらく理由のひとつは,Na+K+2Cl-共輸送体NKCC1の発現が成熟したニューロンよりも高く,ニューロンにおけるCl-濃度が高く保たれるためである9).そこで,新生した顆粒細胞の移動における興奮性GABAA受容体シグナル伝達の影響がNKCC1により担われている可能性を,熱性けいれんラットに由来する歯状回門の移植片の培養系を利用して検証した.この系において,GABAA受容体の作用薬ムシモールの処理により顆粒細胞の移動距離は減少するが,この現象は培地にNKCC1の阻害薬ブメタニドを処理すること,または,siRNA法を用いて顆粒細胞においてNKCC1をノックダウンすることにより阻止された.
4.NKCC1の阻害により異所性の顆粒細胞の出現と将来のてんかんの発症が抑制される
最後に,熱性けいれんによる異所性の顆粒細胞の出現と将来のてんかんの発症との関連を検証した.その結果,熱性けいれんを経験したラットは成体期(11~13週齢)においてピロカルピン誘導性のけいれん発作への感受性の高いことが明らかになった.さらに重要なことに,熱性けいれんを経験したラット16匹のうちの半数では辺縁系に由来する自発発作が確認され,その発作頻度と異所性の顆粒細胞の数とのあいだに正の相関性が確認された.また,熱性けいれんを経験したラットの海馬ではニューロンの過剰な発射の起こっていることをin vivoにおける電気生理学的な手法により明らかにした.一連のin vitroにおける実験により,興奮性GABAA受容体シグナル伝達による異所性の顆粒細胞の出現の根本的な原因はNKCC1であることがつきとめられたため,NKCC1の阻害薬ブメタニドを熱性けいれんの誘導ののち1週間にわたり処理した.その結果,成体期における異所性の顆粒細胞の出現,発作閾値の低下,自発発作の発症,そして,海馬におけるニューロンの過剰な発射が抑制された.
おわりに
複雑型熱性けいれんの発達期の神経回路の形成にあたえる影響,および,将来的なてんかんの発症への関与は明らかではなかった10).この研究では,ラットにおいて複雑型熱性けいれんが興奮性GABAA受容体シグナル伝達を介し異所性の顆粒細胞の出現を誘導することを明らかにした(図2).さらに,異所性の顆粒細胞とてんかんの発症とのあいだには正の相関性が確認された.また,複雑型熱性けいれんののちのブメタニドの投与(NKCC1の阻害)が,将来のてんかんの発症を防止するための新規の治療法となりうる可能性を示した.
文 献
- Chevassus-au-Louis, N., Baraban, S. C., Gaiarsa, J. L. et al.: Cortical malformations and epilepsy: new insights from animal models. Epilepsia, 40, 811-821 (1999)[PubMed]
- Manent, J. B., Wang, Y., Chang, Y. et al.: Dcx reexpression reduces subcortical band heterotopia and seizure threshold in an animal model of neuronal migration disorder. Nat. Med., 15, 84-90 (2009)[PubMed]
- Houser, C. R.: Granule cell dispersion in the dentate gyrus of humans with temporal lobe epilepsy. Brain Res., 535, 195-204 (1990)[PubMed]
- Scharfman, H., Goodman, J. & McCloskey, D.: Ectopic granule cells of the rat dentate gyrus. Dev. Neurosci., 29, 14-27 (2007)[PubMed]
- Hauser, W. A.: The prevalence and incidence of convulsive disorders in children. Epilepsia, 35(Suppl.2), S1-S6 (1994)[PubMed]
- Cendes, F., Andermann, F., Dubeau, F. et al.: Early childhood prolonged febrile convulsions, atrophy and sclerosis of mesial structures, and temporal lobe epilepsy: an MRI volumetric study. Neurology, 43, 1083-1087 (1993)[PubMed]
- French, J. A., Williamson, P. D., Thadani, V. M. et al.: Characteristics of medial temporal lobe epilepsy: I. Results of history and physical examination. Ann. Neurol., 34, 774-780 (1993)[PubMed]
- Bender, R. A., Dube, C. & Baram, T. Z.: Febrile seizures and mechanisms of epileptogenesis: insights from an animal model. Adv. Exp. Med. Biol., 548, 213-225 (2004)[PubMed]
- Fukuda, A., Muramatsu, K., Okabe, A. et al.: Changes in intracellular Ca2+ induced by GABAA receptor activation and reduction in Cl- gradient in neonatal rat neocortex. J. Neurophysiol., 79, 439-446 (1998)[PubMed]
- Koyama, R. & Matsuki, N.: Novel etiological and therapeutic strategies for neurodiseases: mechanisms and consequences of febrile seizures: lessons from animal models. J. Pharmacol. Sci., 113, 14-22 (2010)[PubMed]
著者プロフィール
略歴:2006年 東京大学大学院薬学系研究科 修了,同年 同 寄付講座教員を経て,2007年より同 助教.2010年より米国Harvard Medical Schoolに留学中.
研究テーマ:発達期および病態の脳における神経回路の形成機構.
抱負:各種の脳疾患における神経回路の形成の変化をオリジナリティーあふれる培養系に再現し,その細胞生物学的および分子生物学的な機構を明らかにするとともに,将来的な創薬をめざしたい.
© 2012 小山 隆太 Licensed under CC 表示 2.1 日本
