マカクザルの前肢の巧緻な運動に必要な神経回路の新しい遺伝子導入法による選択的な遮断
木下正治・伊佐 正
(生理学研究所 認知行動発達研究部門)
email:木下正治,伊佐 正
DOI: 10.7875/first.author.2012.089
Genetic dissection of the circuit for hand dexterity in primates.
Masaharu Kinoshita, Ryosuke Matsui, Shigeki Kato, Taku Hasegawa, Hironori Kasahara, Kaoru Isa, Akiya Watakabe, Tetsuo Yamamori, Yukio Nishimura, Bror Alstermark, Dai Watanabe, Kazuto Kobayashi, Tadashi Isa
Nature, 487, 235-238 (2012)
前肢の到達運動と巧緻な把持運動は霊長類の運動系のなかでももっとも高度に発達しているものである.従来,大脳皮質の運動野から脊髄の運動ニューロンへの直接経路の発達がこのような巧緻な運動の基盤であると考えられてきた.一方,系統発生的に古い脊髄固有ニューロンを介する間接経路が霊長類の手の運動制御においてはたす役割について見解は分かれていた.この特定の経路の機能を明らかにするため,筆者らは,ウイルスベクターを用いた二重感染法とTetシステムとを組み合わせた新しい手法を開発し,マカクザルの脊髄固有ニューロンに選択的かつ可逆的に破傷風毒素を発現させそのシナプス伝達を遮断することに成功した.そして,行動実験と電気生理学的および組織学的な検討により,脊髄固有ニューロンが霊長類の精密な把持運動において重要な役割をはたしていることを明らかにした.この手法は,マカクザルなど細胞種に特異的な遺伝子プロモーターの明らかでない動物種においても,経路選択的に導入遺伝子の発現制御を行うことにより機能解明を行うことができるという点で画期的である.
ヒトやマカクザルなどの霊長類は,それぞれの指を独立に動かして対象物の精密な把持や操作を行うことができる.これらの動物種では大脳皮質の運動野から脊髄の運動ニューロンへの単シナプス性の結合(直接経路)が発達している.一方,ネコなどでは指を独立に動かすことはできないが,これらの動物種には直接経路は存在せず,大脳皮質から脊髄の運動ニューロンへは脊髄固有ニューロンなどの介在ニューロンを介した,最低でも2シナプス性の結合(間接経路)しか存在していない1).直接経路は進化的に新しく,また,サルにおける直接経路の切断実験などにもとづき,この直接経路が手の巧緻な運動に本質的な役割をはたしているものと考えられてきた2).一方,筆者らの研究グループは,マカクザルにおいても脊髄固有ニューロンを介する間接経路の存在していることを示し3,4),直接経路の物理的な切断ののち,頸髄の中部に存在する脊髄固有ニューロンのはたらきにより1~2カ月以内に手指の精密な把持運動の機能が回復することを明らかにした5,6).しかし,これらの結果は物理的な損傷ののちの可塑的な変化により脊髄固有ニューロンが機能の代償にかかわることを示していても,健常な状態での脊髄固有ニューロンの役割については明らかにしていない.むしろ,脊髄の介在ニューロンの役割については,1個1個のニューロンが直接経路のニューロンに比べ多数の種類の筋の運動ニューロンを支配しており,多数の指をまとめたような運動の制御には役立つが,1本1本の指を個別に動かすような精緻な運動の制御には不向きであるという主張もあった7,8).
このような特定の経路の機能を明らかにするためには物理的な損傷実験では限界があり,特定の経路を選択的かつ可逆的に操作することのできる分子遺伝学的なツールが有効である.これまでも,マウスなどでは細胞種に特異的な遺伝子プロモーターを用いた遺伝子改変動物を作製することによりこのような操作が可能であった.しかし,マウスにおいても細胞種に特異的な遺伝子プロモーターを探索することは容易ではなく,まして,遺伝子改変動物の作製が困難な霊長類などではこのような研究はこれまで事実上不可能であった.
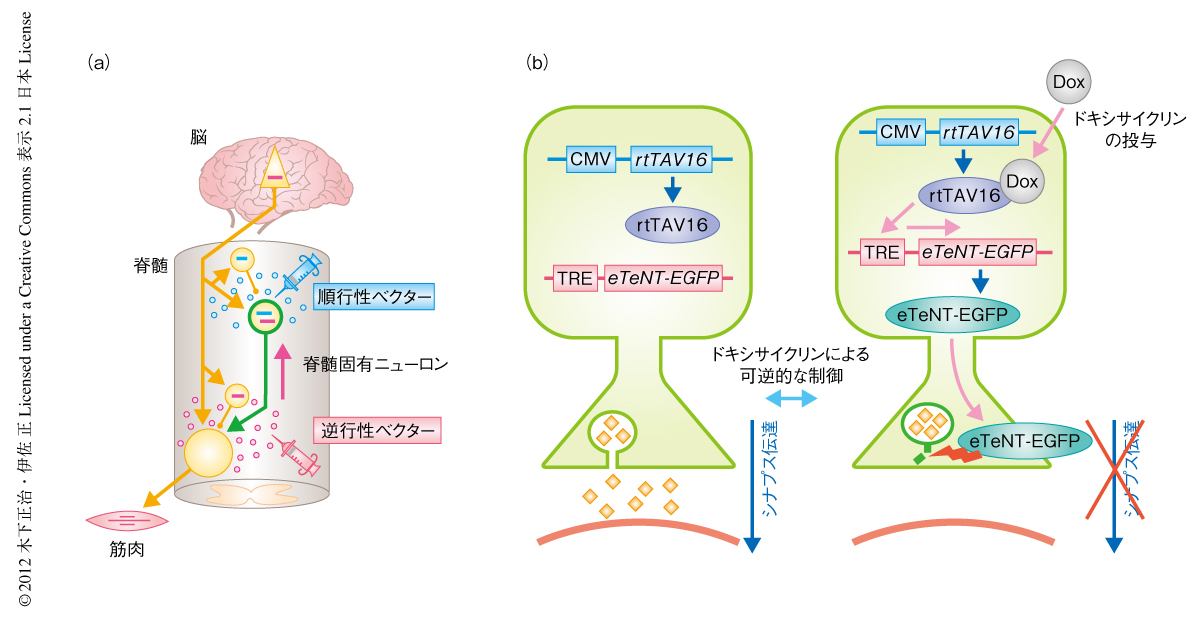
特定の神経経路に選択的な機能操作は,逆行性に輸送されるウイルスベクターと順行性に輸送されるウイルスベクターの2つを組み合わせることで理論上は可能になるとされてきた(図1).しかし実際には,レンチウイルスの膜の表面の糖タンパク質を狂犬病ウイルスの糖タンパク質と置き換えることで作製したRV-Gベクターなどの逆行性ベクターの輸送効率は決して高くなかった.ところが近年,新規に開発されたHiRetベクターはRV-Gベクターに比べ30倍以上も逆行性輸送の効率が高く9),これにより経路選択的な機能操作が現実のものとなった.

二重感染法の概念を説明する(図1).まず,逆行性ベクターを標的とするニューロンの投射先に注入する.つぎに,順行性ベクターを標的とするニューロンの細胞体の領域に注入する.それぞれのベクターは標的とするニューロン以外にも感染するが,2つのベクターによる二重感染は標的とするニューロンでのみ起こる.注入された2つのベクターによりそれぞれコードされた2種類の遺伝子が同じ細胞に存在するときだけ遺伝子発現の起こるしくみにしておくことで,標的とするニューロンに選択的に導入遺伝子を発現させることができる.
この研究では,このウイルスベクター二重感染法と,Tetシステム,破傷風毒素を用いることで,脊髄固有ニューロンのシナプス伝達を選択的かつ可逆的に遮断した(図2).まず,逆行性ベクターであるHiRetベクターを,脊髄固有ニューロンの投射している頚髄第6髄節から胸髄第1髄節の前角(前肢の筋肉を支配する脊髄の運動ニューロンの存在する部位)に注入した.つぎに,逆行性ベクターの注入から7~10日のち,順行性ベクターとしてアデノ随伴ウイルス2型ベクターを,脊髄固有ニューロンの細胞体の存在する頚髄第2髄節から頚髄第5髄節の中間帯に注入した.これにより,脊髄固有ニューロンに選択的に二重感染させた(図2a).逆行性ベクターにはテトラサイクリン応答配列,および,その下流に増強型の破傷風毒素と蛍光タンパク質EGFPとの融合タンパク質の遺伝子を組み込み,順行性ベクターにはテトラサイクリンと結合して高感度でテトラサイクリン応答配列における遺伝子発現を誘導するタンパク質rtTAV16の遺伝子を組み込んだ.ベクターの注入ののちテトラサイクリンの誘導体であるドキシサイクリンを投与すると,rtTAV16と複合体を形成してテトラサイクリン応答配列に作用し,その結果,二重感染した脊髄固有ニューロンにおいてのみ増強型の破傷風毒素が発現する.破傷風毒素が神経伝達物質の放出に必要なVAMP2タンパク質を破壊することにより,シナプス伝達は遮断される.このプロセスは可逆的で,ドキシサイクリンの投与を停止するとシナプス伝達はもとにもどる(図2b).
今回の実験では,4頭のマカクザルに対しこの逆行性ベクターと順行性ベクターを注入した.さらに1頭を対照として,逆行性ベクターから増強型の破傷風毒素の遺伝子をはずしたもの,および,順行性ベクターを注入した.
ベクターの注入から1~2か月後にドキシサイクリンの経口投与を開始した.すると,4頭のマカクザルのいずれにおいても投与の開始から2~5日のちに前肢の巧緻な到達-把持運動の失敗率が増加し運動は遅くなった.一方,対照の個体ではドキシサイクリンの投与によっても障害は観察されなかった.ドキシサイクリンの投与,つまり,脊髄固有ニューロンにおけるシナプス伝達の遮断により運動の障害が起こったことから,脊髄固有ニューロンを介する間接経路が巧緻な到達-把持運動の制御に関与していることが明らかになった.
この機能障害はドキシサイクリンの投与期間において回復した.また,いったんドキシサイクリンの投与を停止し約1か月後に再投与すると,同様な障害がふたたび観察された.これらの結果は,ドキシサイクリンの投与期間において,今回,標的とした脊髄固有ニューロン以外の経路(直接経路など)が機能の代償をしたこと,さらに,ドキシサイクリンの投与の停止により脊髄固有ニューロン経路がシナプス伝達能を回復し,ふたたびその経路が運動制御に使われるようになったことを示唆した.
実際に,脊髄固有ニューロンの何%が遮断されているかを調べるため,ドキシサイクリンの投与の最終日にマカクザルを麻酔し非動化した状態で急性の電気生理実験を行った.なんの操作もしていないマカクザルの場合,延髄の錐体に刺激電極を刺入して大脳皮質から下降する軸索を電気刺激し,刺激と反対側の脊髄の頚髄第7髄節のレベルにおいて手指の運動神経核からフィールド電位を記録すると,直接経路による単シナプス性のフィールド電位が記録され,さらに,その1ミリ秒後くらいに2シナプス性のシナプス伝達を反映するフィールド電位が記録される.無傷な脊髄においてこの2シナプス性のフィールド電位は,頚髄第3髄節から頚髄第5髄節レベルにある脊髄固有ニューロンを介する成分と,運動ニューロンと同じ頚髄第7髄節に存在する髄節内の介在ニューロンを介する成分とが混在しているが,頚髄第5髄節レベルにおいて大脳皮質から下降する軸索を物理的に損傷すると,この2シナプス性のフィールド電位はほとんどすべてが頚髄第3髄節から頚髄第5髄節の脊髄固有ニューロンを介する成分のみとなる.この成分を,二重感染法による遺伝子導入を行った側の脊髄と,ベクターを注入していない健常な側の脊髄とで比較したところ,二重感染により脊髄固有ニューロンを介する成分の約90%が抑制されていることが明らかになった.つまり,二重感染法によるシナプス伝達の選択的な遮断の効果を,行動実験のみならず,電気生理学的にも証明することができた.
これまで用いられてきた電気刺激法や薬物注入法による神経回路の操作では,その効果の範囲を正確に知ることはほぼ不可能であった.一方,遺伝子導入による操作では導入遺伝子にマーカーとなる遺伝子を付加しておくことで操作の対象となったニューロンを可視化することができる.この研究では,破傷風毒素を蛍光タンパク質EGFPとの融合タンパク質として発現させることでシナプス伝達の遮断されたニューロンの可視化を行った.
組織学的な検査を行うと,順行性ベクターを注入した髄節のうち,頚髄第3髄節から頚髄第5髄節にかけて,VII層を中心とする中間帯の外側に多数のEGFP陽性細胞が見い出された.これらの分布は,これまでネコやマカクザルで知られていた脊髄固有ニューロンの分布と一致していた.さらに,免疫組織染色を行うと,細胞体だけでなく,胸髄の運動神経核でも軸索の走行を確認することができた.運動神経核においては,実際に運動ニューロンと考えられる大型の細胞に終末をもつ軸索の像が多く確認され,今回,シナプス伝達の遮断されたのは運動ニューロンに伝達する脊髄固有ニューロンであることが確認できた.また,脊髄固有ニューロンの一部は脳幹の外側網様核にも軸索の分枝を延ばしていることが知られているが,この研究では,運動神経核だけでなく,脳幹の外側網様核に終末をもつ軸索も可視化することができた.このように,シナプス伝達の遮断を行ったニューロンを組織学的に明確にすることは,その機能を考察するうえで大きな利点となるだろう.
以上の結果により,霊長類において系統発生的に古い脊髄固有ニューロンを介する間接経路は,前肢の巧緻な到達-把持運動の制御に関与していることが明らかになった.
さらに今回の研究により,細胞種に特異的な遺伝子プロモーターを用いなくても,解剖学的な結合関係をもとに2種類のウイルスベクターを組み合わせることで,経路選択的かつ可逆的なシナプス伝達の遮断が可能になった.このような手法は,遺伝子改変動物を作製できない霊長類などの大型動物における高次の脳機能の解明,また,遺伝子改変動物の作製の可能な種の研究においても細胞種に特異的な遺伝子プロモーターのみつかっていない多くの経路の機能の解明において,たいへん有効なツールとなることが期待される.
略歴:1998年 筑波大学大学院医学研究科 修了,同年 生理学研究所 リサーチアソシエイト,2001年 米国Rockefeller大学 博士研究員,2009年 生理学研究所 専門研究職員を経て,2010年より同 特任助教.
研究テーマ:高等動物の認知および行動制御の基礎的な神経機構.
抱負:個人的に,これまでの研究はマカクザルなどを用いた電気生理学的な手法がメインであったが,遺伝子操作をはじめとした新しい手法を取り入れ,これまでの手法では解決できなかった課題に挑戦したい.また,今回,開発した手法は広く応用が可能で有用な方法なので積極的に広めていきたい.
伊佐 正(Tadashi Isa)
生理学研究所 教授.
研究室URL:http://www.nips.ac.jp/hbfp/
© 2012 木下正治・伊佐 正 Licensed under CC 表示 2.1 日本
(生理学研究所 認知行動発達研究部門)
email:木下正治,伊佐 正
DOI: 10.7875/first.author.2012.089
Genetic dissection of the circuit for hand dexterity in primates.
Masaharu Kinoshita, Ryosuke Matsui, Shigeki Kato, Taku Hasegawa, Hironori Kasahara, Kaoru Isa, Akiya Watakabe, Tetsuo Yamamori, Yukio Nishimura, Bror Alstermark, Dai Watanabe, Kazuto Kobayashi, Tadashi Isa
Nature, 487, 235-238 (2012)
要 約
前肢の到達運動と巧緻な把持運動は霊長類の運動系のなかでももっとも高度に発達しているものである.従来,大脳皮質の運動野から脊髄の運動ニューロンへの直接経路の発達がこのような巧緻な運動の基盤であると考えられてきた.一方,系統発生的に古い脊髄固有ニューロンを介する間接経路が霊長類の手の運動制御においてはたす役割について見解は分かれていた.この特定の経路の機能を明らかにするため,筆者らは,ウイルスベクターを用いた二重感染法とTetシステムとを組み合わせた新しい手法を開発し,マカクザルの脊髄固有ニューロンに選択的かつ可逆的に破傷風毒素を発現させそのシナプス伝達を遮断することに成功した.そして,行動実験と電気生理学的および組織学的な検討により,脊髄固有ニューロンが霊長類の精密な把持運動において重要な役割をはたしていることを明らかにした.この手法は,マカクザルなど細胞種に特異的な遺伝子プロモーターの明らかでない動物種においても,経路選択的に導入遺伝子の発現制御を行うことにより機能解明を行うことができるという点で画期的である.
はじめに
ヒトやマカクザルなどの霊長類は,それぞれの指を独立に動かして対象物の精密な把持や操作を行うことができる.これらの動物種では大脳皮質の運動野から脊髄の運動ニューロンへの単シナプス性の結合(直接経路)が発達している.一方,ネコなどでは指を独立に動かすことはできないが,これらの動物種には直接経路は存在せず,大脳皮質から脊髄の運動ニューロンへは脊髄固有ニューロンなどの介在ニューロンを介した,最低でも2シナプス性の結合(間接経路)しか存在していない1).直接経路は進化的に新しく,また,サルにおける直接経路の切断実験などにもとづき,この直接経路が手の巧緻な運動に本質的な役割をはたしているものと考えられてきた2).一方,筆者らの研究グループは,マカクザルにおいても脊髄固有ニューロンを介する間接経路の存在していることを示し3,4),直接経路の物理的な切断ののち,頸髄の中部に存在する脊髄固有ニューロンのはたらきにより1~2カ月以内に手指の精密な把持運動の機能が回復することを明らかにした5,6).しかし,これらの結果は物理的な損傷ののちの可塑的な変化により脊髄固有ニューロンが機能の代償にかかわることを示していても,健常な状態での脊髄固有ニューロンの役割については明らかにしていない.むしろ,脊髄の介在ニューロンの役割については,1個1個のニューロンが直接経路のニューロンに比べ多数の種類の筋の運動ニューロンを支配しており,多数の指をまとめたような運動の制御には役立つが,1本1本の指を個別に動かすような精緻な運動の制御には不向きであるという主張もあった7,8).
このような特定の経路の機能を明らかにするためには物理的な損傷実験では限界があり,特定の経路を選択的かつ可逆的に操作することのできる分子遺伝学的なツールが有効である.これまでも,マウスなどでは細胞種に特異的な遺伝子プロモーターを用いた遺伝子改変動物を作製することによりこのような操作が可能であった.しかし,マウスにおいても細胞種に特異的な遺伝子プロモーターを探索することは容易ではなく,まして,遺伝子改変動物の作製が困難な霊長類などではこのような研究はこれまで事実上不可能であった.
特定の神経経路に選択的な機能操作は,逆行性に輸送されるウイルスベクターと順行性に輸送されるウイルスベクターの2つを組み合わせることで理論上は可能になるとされてきた(図1).しかし実際には,レンチウイルスの膜の表面の糖タンパク質を狂犬病ウイルスの糖タンパク質と置き換えることで作製したRV-Gベクターなどの逆行性ベクターの輸送効率は決して高くなかった.ところが近年,新規に開発されたHiRetベクターはRV-Gベクターに比べ30倍以上も逆行性輸送の効率が高く9),これにより経路選択的な機能操作が現実のものとなった.
1.ウイルスベクター二重感染法による経路選択的なシナプス伝達の遮断
二重感染法の概念を説明する(図1).まず,逆行性ベクターを標的とするニューロンの投射先に注入する.つぎに,順行性ベクターを標的とするニューロンの細胞体の領域に注入する.それぞれのベクターは標的とするニューロン以外にも感染するが,2つのベクターによる二重感染は標的とするニューロンでのみ起こる.注入された2つのベクターによりそれぞれコードされた2種類の遺伝子が同じ細胞に存在するときだけ遺伝子発現の起こるしくみにしておくことで,標的とするニューロンに選択的に導入遺伝子を発現させることができる.
この研究では,このウイルスベクター二重感染法と,Tetシステム,破傷風毒素を用いることで,脊髄固有ニューロンのシナプス伝達を選択的かつ可逆的に遮断した(図2).まず,逆行性ベクターであるHiRetベクターを,脊髄固有ニューロンの投射している頚髄第6髄節から胸髄第1髄節の前角(前肢の筋肉を支配する脊髄の運動ニューロンの存在する部位)に注入した.つぎに,逆行性ベクターの注入から7~10日のち,順行性ベクターとしてアデノ随伴ウイルス2型ベクターを,脊髄固有ニューロンの細胞体の存在する頚髄第2髄節から頚髄第5髄節の中間帯に注入した.これにより,脊髄固有ニューロンに選択的に二重感染させた(図2a).逆行性ベクターにはテトラサイクリン応答配列,および,その下流に増強型の破傷風毒素と蛍光タンパク質EGFPとの融合タンパク質の遺伝子を組み込み,順行性ベクターにはテトラサイクリンと結合して高感度でテトラサイクリン応答配列における遺伝子発現を誘導するタンパク質rtTAV16の遺伝子を組み込んだ.ベクターの注入ののちテトラサイクリンの誘導体であるドキシサイクリンを投与すると,rtTAV16と複合体を形成してテトラサイクリン応答配列に作用し,その結果,二重感染した脊髄固有ニューロンにおいてのみ増強型の破傷風毒素が発現する.破傷風毒素が神経伝達物質の放出に必要なVAMP2タンパク質を破壊することにより,シナプス伝達は遮断される.このプロセスは可逆的で,ドキシサイクリンの投与を停止するとシナプス伝達はもとにもどる(図2b).
今回の実験では,4頭のマカクザルに対しこの逆行性ベクターと順行性ベクターを注入した.さらに1頭を対照として,逆行性ベクターから増強型の破傷風毒素の遺伝子をはずしたもの,および,順行性ベクターを注入した.
2.脊髄固有ニューロンにおける選択的なシナプス伝達の遮断により前肢の巧緻な運動は阻害された
ベクターの注入から1~2か月後にドキシサイクリンの経口投与を開始した.すると,4頭のマカクザルのいずれにおいても投与の開始から2~5日のちに前肢の巧緻な到達-把持運動の失敗率が増加し運動は遅くなった.一方,対照の個体ではドキシサイクリンの投与によっても障害は観察されなかった.ドキシサイクリンの投与,つまり,脊髄固有ニューロンにおけるシナプス伝達の遮断により運動の障害が起こったことから,脊髄固有ニューロンを介する間接経路が巧緻な到達-把持運動の制御に関与していることが明らかになった.
この機能障害はドキシサイクリンの投与期間において回復した.また,いったんドキシサイクリンの投与を停止し約1か月後に再投与すると,同様な障害がふたたび観察された.これらの結果は,ドキシサイクリンの投与期間において,今回,標的とした脊髄固有ニューロン以外の経路(直接経路など)が機能の代償をしたこと,さらに,ドキシサイクリンの投与の停止により脊髄固有ニューロン経路がシナプス伝達能を回復し,ふたたびその経路が運動制御に使われるようになったことを示唆した.
3.シナプス伝達の遮断の電気生理学的な検証
実際に,脊髄固有ニューロンの何%が遮断されているかを調べるため,ドキシサイクリンの投与の最終日にマカクザルを麻酔し非動化した状態で急性の電気生理実験を行った.なんの操作もしていないマカクザルの場合,延髄の錐体に刺激電極を刺入して大脳皮質から下降する軸索を電気刺激し,刺激と反対側の脊髄の頚髄第7髄節のレベルにおいて手指の運動神経核からフィールド電位を記録すると,直接経路による単シナプス性のフィールド電位が記録され,さらに,その1ミリ秒後くらいに2シナプス性のシナプス伝達を反映するフィールド電位が記録される.無傷な脊髄においてこの2シナプス性のフィールド電位は,頚髄第3髄節から頚髄第5髄節レベルにある脊髄固有ニューロンを介する成分と,運動ニューロンと同じ頚髄第7髄節に存在する髄節内の介在ニューロンを介する成分とが混在しているが,頚髄第5髄節レベルにおいて大脳皮質から下降する軸索を物理的に損傷すると,この2シナプス性のフィールド電位はほとんどすべてが頚髄第3髄節から頚髄第5髄節の脊髄固有ニューロンを介する成分のみとなる.この成分を,二重感染法による遺伝子導入を行った側の脊髄と,ベクターを注入していない健常な側の脊髄とで比較したところ,二重感染により脊髄固有ニューロンを介する成分の約90%が抑制されていることが明らかになった.つまり,二重感染法によるシナプス伝達の選択的な遮断の効果を,行動実験のみならず,電気生理学的にも証明することができた.
4.シナプス伝達の遮断された脊髄固有ニューロンの可視化
これまで用いられてきた電気刺激法や薬物注入法による神経回路の操作では,その効果の範囲を正確に知ることはほぼ不可能であった.一方,遺伝子導入による操作では導入遺伝子にマーカーとなる遺伝子を付加しておくことで操作の対象となったニューロンを可視化することができる.この研究では,破傷風毒素を蛍光タンパク質EGFPとの融合タンパク質として発現させることでシナプス伝達の遮断されたニューロンの可視化を行った.
組織学的な検査を行うと,順行性ベクターを注入した髄節のうち,頚髄第3髄節から頚髄第5髄節にかけて,VII層を中心とする中間帯の外側に多数のEGFP陽性細胞が見い出された.これらの分布は,これまでネコやマカクザルで知られていた脊髄固有ニューロンの分布と一致していた.さらに,免疫組織染色を行うと,細胞体だけでなく,胸髄の運動神経核でも軸索の走行を確認することができた.運動神経核においては,実際に運動ニューロンと考えられる大型の細胞に終末をもつ軸索の像が多く確認され,今回,シナプス伝達の遮断されたのは運動ニューロンに伝達する脊髄固有ニューロンであることが確認できた.また,脊髄固有ニューロンの一部は脳幹の外側網様核にも軸索の分枝を延ばしていることが知られているが,この研究では,運動神経核だけでなく,脳幹の外側網様核に終末をもつ軸索も可視化することができた.このように,シナプス伝達の遮断を行ったニューロンを組織学的に明確にすることは,その機能を考察するうえで大きな利点となるだろう.
おわりに
以上の結果により,霊長類において系統発生的に古い脊髄固有ニューロンを介する間接経路は,前肢の巧緻な到達-把持運動の制御に関与していることが明らかになった.
さらに今回の研究により,細胞種に特異的な遺伝子プロモーターを用いなくても,解剖学的な結合関係をもとに2種類のウイルスベクターを組み合わせることで,経路選択的かつ可逆的なシナプス伝達の遮断が可能になった.このような手法は,遺伝子改変動物を作製できない霊長類などの大型動物における高次の脳機能の解明,また,遺伝子改変動物の作製の可能な種の研究においても細胞種に特異的な遺伝子プロモーターのみつかっていない多くの経路の機能の解明において,たいへん有効なツールとなることが期待される.
文 献
- Alstermark, B. & Isa, T.: Circuits for skilled reaching and grasping. Annu. Rev. Neurosci., 35, 559-578 (2012)[PubMed]
- Lawrence, D. G. & Kuypers, H. G.: The functional organization of the motor system in the monkey. I. The effect of bilateral pyramidal lesions. Brain, 91, 1-14 (1968)[PubMed]
- Alstermark, B., Isa, T., Ohki, Y. et al.: Disynaptic pyramidal excitation in forelimb motoneurons mediated via C3-C4 propriospinal neurons in the Macaca fuscata. J. Neurophysiol., 82, 3580-3585 (1999)[PubMed]
- Isa, T., Ohki, Y., Seki, K. et al.: Properties of propriospinal neurons in the C3-C4 segments mediating disynaptic pyramidal excitation to forelimb motoneurons in the macaque monkey. J. Neurophysiol., 95, 3674-3685 (2006)[PubMed]
- Sasaki, S., Isa, T., Pettersson, L. G. et al.: Dexterous finger movements in primate without monosynaptic corticomotoneuronal excitation. J. Neurophysiol., 92, 3142-3147 (2004)[PubMed]
- Alstermark, B., Pettersson, L. G., Nishimura, Y. et al.: Motor command for precision grip in the macaque monkey can be mediated by spinal interneurons. J. Neurophysiol., 106, 122-126 (2011)[PubMed]
- Kuypers, H. G.: A new look at the organization of the motor system. Prog. Brain Res., 57, 381-403 (1982)[PubMed]
- Lemon, R. N.: Descending pathways in motor control. Annu. Rev. Neurosci., 31, 195-218 (2008)[PubMed]
- Kato, S., Kobayashi, K., Inoue, K. et al.: A lentiviral strategy for highly efficient retrograde gene transfer by pseudotyping with fusion envelope glycoprotein. Hum. Gene Ther., 22, 197-206 (2011)[PubMed]
著者プロフィール
略歴:1998年 筑波大学大学院医学研究科 修了,同年 生理学研究所 リサーチアソシエイト,2001年 米国Rockefeller大学 博士研究員,2009年 生理学研究所 専門研究職員を経て,2010年より同 特任助教.
研究テーマ:高等動物の認知および行動制御の基礎的な神経機構.
抱負:個人的に,これまでの研究はマカクザルなどを用いた電気生理学的な手法がメインであったが,遺伝子操作をはじめとした新しい手法を取り入れ,これまでの手法では解決できなかった課題に挑戦したい.また,今回,開発した手法は広く応用が可能で有用な方法なので積極的に広めていきたい.
伊佐 正(Tadashi Isa)
生理学研究所 教授.
研究室URL:http://www.nips.ac.jp/hbfp/
© 2012 木下正治・伊佐 正 Licensed under CC 表示 2.1 日本
