RIG-I様受容体経路とToll様受容体経路との干渉による細菌に対するT細胞の応答の抑制
根岸英雄・柳井秀元・中島 啓・谷口維紹
(東京大学医学系研究科 免疫学教室)
email:谷口維紹
DOI: 10.7875/first.author.2012.066
Cross-interference of RLR and TLR signaling pathways modulates antibacterial T cell responses.
Hideo Negishi, Hideyuki Yanai, Akira Nakajima, Ryuji Koshiba, Koji Atarashi, Atsushi Matsuda, Kosuke Matsuki, Shoji Miki, Takahiro Doi, Alan Aderem, Junko Nishio, Stephen T Smale, Kenya Honda, Tadatsugu Taniguchi
Nature Immunology, 13, 659-666 (2012)
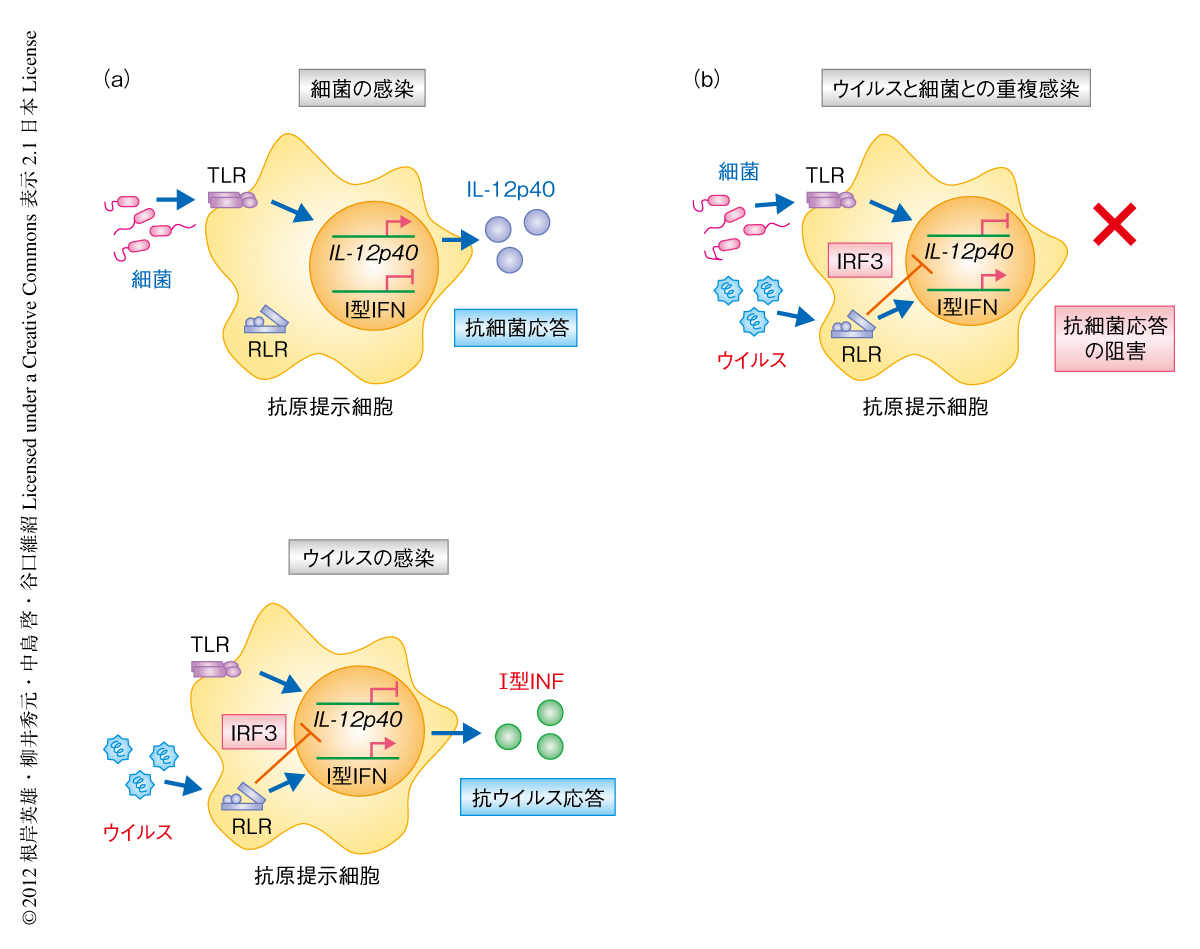
ウイルスに感染した患者がさらなる細菌の感染により非常に重篤な症状を起こすことは広く知られているが,その背後にある分子機構については不明な点が多い.筆者らは,生体がウイルスに対し活性化する防御応答が,同時に,細菌に対する防御応答を抑制する性質をもつことを見い出した.さらに,その分子制御の機構について,ウイルスに由来する核酸により強力に活性化される転写因子IRF3は,抗ウイルス応答に必須であるI型インターフェロンの遺伝子発現を活性化する一方,抗細菌応答に重要なインターロイキン12のp40サブユニットの遺伝子発現を抑制する,という二面性をもつことを発見した.この機構により,ウイルスに感染したマウスでは細菌の感染に対するTh1細胞およびTh17細胞の応答が強く抑制され,微量の細菌の感染に対しても高い感受性を示すようになることが明らかになった.IRF3による抗細菌応答の抑制機構は免疫応答における弱点ともいえ,さまざまなウイルスあるいは細菌による重複感染症に関与しているものと考えられる.一方で,インターロイキン12のp40サブユニットはT細胞の応答を制御する重要なタンパク質であるため,今回,発見された機構はT細胞の応答によりひき起こされるさまざまなアレルギーや自己免疫疾患,抗腫瘍応答に関係する可能性がある.今後,この機構をさらに詳細に解析することにより,ウイルスの感染と関連するこれら疾患の理解や,さまざまな重複感染症の治療法の確立へとつながる可能性がある.
ウイルスと細菌との重複感染がそれぞれ単独の感染と比較して重篤な症状をひき起こすことは広く知られ,重複感染症として臨床的にも重要であるが,なぜそのような重複感染が症状を重篤化するかについてはよくわかっていなかった1,2).このような重複感染症は呼吸器系へのウイルスの感染の際に起こるケースがよく知られていたことから,インフルエンザなどいくつかのウイルスについては,そのウイルスのもつ特性として抗細菌応答を抑制する機構が存在するという報告もあったが,このような機構は特定のウイルスにかぎられたものであり,それ以外のさまざまなウイルスと細菌との組合せによりひき起こされる多様な重複感染症の背後にある分子機構を説明できるものではなかった.また,いままでの重複感染症の解析の焦点はおもに病原体の性質にあり,宿主の免疫応答,とくに,自然免疫応答のシグナル伝達経路に主眼をおいた解析はほとんどなされていなかった1).
病原体の感染に対する生体の防御応答はさまざまな病原体を認識する受容体により担われている.ウイルスの感染を感知する受容体としてはRIG-I様受容体が知られており,RIG-I様受容体経路の下流では転写因子IRF3が活性化され,I型インターフェロンの発現を介して抗ウイルス応答の惹起されることがわかっている3-5).一方で,細菌の感染を感知する受容体としてToll様受容体が知られ,Toll様受容体経路が生体防御に重要な役割をはたしていることがわかっている3,6-9).しかしながら,RIG-I様受容体経路とToll様受容体経路とにおいてひき起こされる免疫応答の違いやその詳細な制御機構,さらには,重複感染の際に互いの経路がどう影響しあっているかについては知られていなかった.
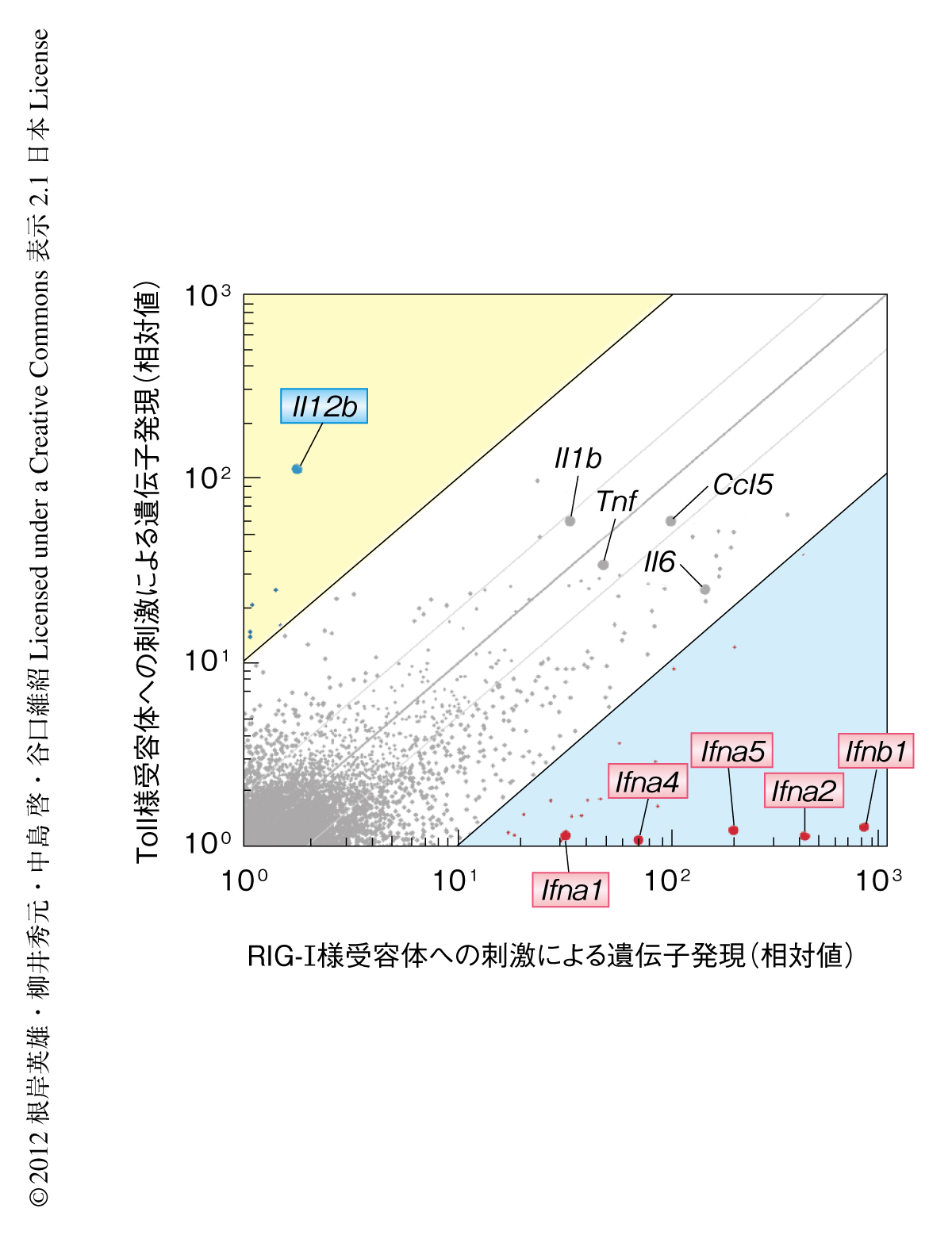
まず,ウイルスの感染あるいは細菌の感染により抗原提示細胞において発現誘導される遺伝子を網羅的に比較解析した.その結果,ウイルスの感染ではRIG-I様受容体経路を介しI型インターフェロンの遺伝子が,細菌の感染ではToll様受容体経路を介しインターロイキン12のp40サブユニット(IL-12p40)の遺伝子(Il12b)が,強力かつ特異的に発現誘導されることがわかった(図1).これらの遺伝子はそれぞれ抗ウイルス応答および抗細菌応答に必須であり,とくに,IL-12p40はTh1細胞およびTh17細胞の応答を活性化し獲得免疫応答の方向性を制御することが知られている.そのため,ウイルスの感染あるいは細菌の感染により誘導される獲得免疫応答を比較したところ,ウイルスの感染はTh2細胞の応答をより強く誘導する一方で,細菌の感染はTh1細胞およびTh17細胞の応答を著明に誘導することが判明した.
ウイルスあるいは細菌それぞれの感染により特異的に誘導される免疫応答が明らかになったので,ウイルスの感染により活性化する応答が,細菌の感染により活性化される応答にあたえる影響を解析した.その結果,ウイルスを認識するRIG-I様受容体経路が活性化した状態では,細菌の感染により活性化するToll様受容体経路により誘導されるIL-12p40遺伝子の発現が強く抑制されることを発見した(図2).一方で,IRF3を欠損するとRIG-I様受容体経路が活性化した状態でもToll様受容体経路によるIL-12p40遺伝子の発現誘導が抑制されないことから(図2),さらに詳細にこの抑制機構を解析した結果,ウイルスに由来する核酸により強力に活性化されたIRF3がIL-12p40遺伝子のプロモーターに結合し転写を抑制していることを発見した.

これらの結果から,ウイルスの感染がIL-12p40遺伝子の発現を抑制することで抗細菌応答を抑制していると考えられた.この仮説を生体レベルにおいて検証するため,マウスに水泡性口内炎ウイルスおよびリステリア菌を重複感染させ,ウイルスの感染が抗細菌応答にあたえる影響を解析した.その結果,水泡性口内炎ウイルスを感染したマウスでは,リステリア菌の感染により誘導されるIL-12p40遺伝子の発現が低下し,Th1細胞およびTh17細胞の応答が抑制されることが明らかになった.この抗細菌応答の抑制により,水泡性口内炎ウイルスとリステリア菌とを重複感染したマウスでは,リステリア菌が単独で感染したマウスと比較してリステリア菌の量が1000倍以上に増加し,著明な臓器の障害および早期の死亡が起こった.一方で,IRF3ノックアウトマウスでは水泡性口内炎ウイルスとリステリア菌との重複感染の際にもIL-12p40遺伝子の発現誘導は抑制されず,Th1細胞およびTh17細胞の応答も正常に誘導され,野生型マウスよりも著明な耐性を示し生存率の上昇および臓器の障害の低減が認められた.
この研究において発見された抗ウイルス応答による抗細菌応答の抑制機構(図3)は,宿主の免疫応答の性質からもたらされる免疫応答の弱点ともいえる機構であり,少なくとも検討した範囲ではウイルスおよび細菌の種類に依存していなかった.そのため,さまざまなウイルスと細菌とによる重複感染症においてこの機構が同じように関与している可能性がある.今後,この機構を標的とすることで,さまざまな重複感染症の治療法の確立が期待できると考えている.
一方で,IL-12p40により活性化されるTh1細胞およびTh17細胞の応答は,炎症反応を増強し自己免疫疾患を増悪することが知られている.ウイルスの感染が自己免疫疾患の病態に関与するケースも報告されているため,この研究において見い出されたIRF3によるIL-12p40の発現の抑制機構は,ウイルスの感染による過剰な炎症応答や自己免疫疾患の発症のリスクを抑えるブレーキ役をはたしている可能性がある.また,IL-12p40にはアレルギー疾患の増悪に関与することの報告されているTh2細胞の応答を抑制する性質もあることから,今回,見い出された機構がウイルスの感染によるアレルギー疾患の増悪に関与していることも考えられる.さらに,Th1細胞は抗腫瘍応答にかかわることも知られていることから,ウイルスの感染の際のTh1細胞の応答の抑制は腫瘍ウイルスによる発がんの促進機構のひとつとして機能している可能性もある.今回の発見を礎として,ウイルスの感染が関連するこれらの疾患についての理解がさらに深まるものと期待している.
略歴:2006年 東京大学大学院医学系研究科 修了,同年 同 研究員,2008年 同 特任助教,2012年より東京大学生産技術研究所 特任助教.
研究テーマ:自然免疫応答の制御機構.
抱負:新しく解明した自然免疫応答の分子制御の機構を疾患の予防や治療に役立てたい.
柳井 秀元(Hideyuki Yanai)
東京大学生産技術研究所 特任助教.
中島 啓(Akira Nakajima)
京都大学医学系研究科 研究員.
谷口 維紹(Tadatsugu Taniguchi)
東京大学生産技術研究所 特任教授.
研究室URL:http://www.immunol.m.u-tokyo.ac.jp/page%20files/ja/top/top-ja.html
© 2012 根岸英雄・柳井秀元・中島 啓・谷口維紹 Licensed under CC 表示 2.1 日本
(東京大学医学系研究科 免疫学教室)
email:谷口維紹
DOI: 10.7875/first.author.2012.066
Cross-interference of RLR and TLR signaling pathways modulates antibacterial T cell responses.
Hideo Negishi, Hideyuki Yanai, Akira Nakajima, Ryuji Koshiba, Koji Atarashi, Atsushi Matsuda, Kosuke Matsuki, Shoji Miki, Takahiro Doi, Alan Aderem, Junko Nishio, Stephen T Smale, Kenya Honda, Tadatsugu Taniguchi
Nature Immunology, 13, 659-666 (2012)
要 約
ウイルスに感染した患者がさらなる細菌の感染により非常に重篤な症状を起こすことは広く知られているが,その背後にある分子機構については不明な点が多い.筆者らは,生体がウイルスに対し活性化する防御応答が,同時に,細菌に対する防御応答を抑制する性質をもつことを見い出した.さらに,その分子制御の機構について,ウイルスに由来する核酸により強力に活性化される転写因子IRF3は,抗ウイルス応答に必須であるI型インターフェロンの遺伝子発現を活性化する一方,抗細菌応答に重要なインターロイキン12のp40サブユニットの遺伝子発現を抑制する,という二面性をもつことを発見した.この機構により,ウイルスに感染したマウスでは細菌の感染に対するTh1細胞およびTh17細胞の応答が強く抑制され,微量の細菌の感染に対しても高い感受性を示すようになることが明らかになった.IRF3による抗細菌応答の抑制機構は免疫応答における弱点ともいえ,さまざまなウイルスあるいは細菌による重複感染症に関与しているものと考えられる.一方で,インターロイキン12のp40サブユニットはT細胞の応答を制御する重要なタンパク質であるため,今回,発見された機構はT細胞の応答によりひき起こされるさまざまなアレルギーや自己免疫疾患,抗腫瘍応答に関係する可能性がある.今後,この機構をさらに詳細に解析することにより,ウイルスの感染と関連するこれら疾患の理解や,さまざまな重複感染症の治療法の確立へとつながる可能性がある.
はじめに
ウイルスと細菌との重複感染がそれぞれ単独の感染と比較して重篤な症状をひき起こすことは広く知られ,重複感染症として臨床的にも重要であるが,なぜそのような重複感染が症状を重篤化するかについてはよくわかっていなかった1,2).このような重複感染症は呼吸器系へのウイルスの感染の際に起こるケースがよく知られていたことから,インフルエンザなどいくつかのウイルスについては,そのウイルスのもつ特性として抗細菌応答を抑制する機構が存在するという報告もあったが,このような機構は特定のウイルスにかぎられたものであり,それ以外のさまざまなウイルスと細菌との組合せによりひき起こされる多様な重複感染症の背後にある分子機構を説明できるものではなかった.また,いままでの重複感染症の解析の焦点はおもに病原体の性質にあり,宿主の免疫応答,とくに,自然免疫応答のシグナル伝達経路に主眼をおいた解析はほとんどなされていなかった1).
病原体の感染に対する生体の防御応答はさまざまな病原体を認識する受容体により担われている.ウイルスの感染を感知する受容体としてはRIG-I様受容体が知られており,RIG-I様受容体経路の下流では転写因子IRF3が活性化され,I型インターフェロンの発現を介して抗ウイルス応答の惹起されることがわかっている3-5).一方で,細菌の感染を感知する受容体としてToll様受容体が知られ,Toll様受容体経路が生体防御に重要な役割をはたしていることがわかっている3,6-9).しかしながら,RIG-I様受容体経路とToll様受容体経路とにおいてひき起こされる免疫応答の違いやその詳細な制御機構,さらには,重複感染の際に互いの経路がどう影響しあっているかについては知られていなかった.
1.ウイルスあるいは細菌の感染により抗原提示細胞において発現誘導される遺伝子
まず,ウイルスの感染あるいは細菌の感染により抗原提示細胞において発現誘導される遺伝子を網羅的に比較解析した.その結果,ウイルスの感染ではRIG-I様受容体経路を介しI型インターフェロンの遺伝子が,細菌の感染ではToll様受容体経路を介しインターロイキン12のp40サブユニット(IL-12p40)の遺伝子(Il12b)が,強力かつ特異的に発現誘導されることがわかった(図1).これらの遺伝子はそれぞれ抗ウイルス応答および抗細菌応答に必須であり,とくに,IL-12p40はTh1細胞およびTh17細胞の応答を活性化し獲得免疫応答の方向性を制御することが知られている.そのため,ウイルスの感染あるいは細菌の感染により誘導される獲得免疫応答を比較したところ,ウイルスの感染はTh2細胞の応答をより強く誘導する一方で,細菌の感染はTh1細胞およびTh17細胞の応答を著明に誘導することが判明した.
2.ウイルスの感染によるRIG-I様受容体経路を介したToll様受容体経路の抑制
ウイルスあるいは細菌それぞれの感染により特異的に誘導される免疫応答が明らかになったので,ウイルスの感染により活性化する応答が,細菌の感染により活性化される応答にあたえる影響を解析した.その結果,ウイルスを認識するRIG-I様受容体経路が活性化した状態では,細菌の感染により活性化するToll様受容体経路により誘導されるIL-12p40遺伝子の発現が強く抑制されることを発見した(図2).一方で,IRF3を欠損するとRIG-I様受容体経路が活性化した状態でもToll様受容体経路によるIL-12p40遺伝子の発現誘導が抑制されないことから(図2),さらに詳細にこの抑制機構を解析した結果,ウイルスに由来する核酸により強力に活性化されたIRF3がIL-12p40遺伝子のプロモーターに結合し転写を抑制していることを発見した.
3.マウス個体におけるウイルスの感染による抗細菌応答の抑制
これらの結果から,ウイルスの感染がIL-12p40遺伝子の発現を抑制することで抗細菌応答を抑制していると考えられた.この仮説を生体レベルにおいて検証するため,マウスに水泡性口内炎ウイルスおよびリステリア菌を重複感染させ,ウイルスの感染が抗細菌応答にあたえる影響を解析した.その結果,水泡性口内炎ウイルスを感染したマウスでは,リステリア菌の感染により誘導されるIL-12p40遺伝子の発現が低下し,Th1細胞およびTh17細胞の応答が抑制されることが明らかになった.この抗細菌応答の抑制により,水泡性口内炎ウイルスとリステリア菌とを重複感染したマウスでは,リステリア菌が単独で感染したマウスと比較してリステリア菌の量が1000倍以上に増加し,著明な臓器の障害および早期の死亡が起こった.一方で,IRF3ノックアウトマウスでは水泡性口内炎ウイルスとリステリア菌との重複感染の際にもIL-12p40遺伝子の発現誘導は抑制されず,Th1細胞およびTh17細胞の応答も正常に誘導され,野生型マウスよりも著明な耐性を示し生存率の上昇および臓器の障害の低減が認められた.
おわりに
この研究において発見された抗ウイルス応答による抗細菌応答の抑制機構(図3)は,宿主の免疫応答の性質からもたらされる免疫応答の弱点ともいえる機構であり,少なくとも検討した範囲ではウイルスおよび細菌の種類に依存していなかった.そのため,さまざまなウイルスと細菌とによる重複感染症においてこの機構が同じように関与している可能性がある.今後,この機構を標的とすることで,さまざまな重複感染症の治療法の確立が期待できると考えている.
一方で,IL-12p40により活性化されるTh1細胞およびTh17細胞の応答は,炎症反応を増強し自己免疫疾患を増悪することが知られている.ウイルスの感染が自己免疫疾患の病態に関与するケースも報告されているため,この研究において見い出されたIRF3によるIL-12p40の発現の抑制機構は,ウイルスの感染による過剰な炎症応答や自己免疫疾患の発症のリスクを抑えるブレーキ役をはたしている可能性がある.また,IL-12p40にはアレルギー疾患の増悪に関与することの報告されているTh2細胞の応答を抑制する性質もあることから,今回,見い出された機構がウイルスの感染によるアレルギー疾患の増悪に関与していることも考えられる.さらに,Th1細胞は抗腫瘍応答にかかわることも知られていることから,ウイルスの感染の際のTh1細胞の応答の抑制は腫瘍ウイルスによる発がんの促進機構のひとつとして機能している可能性もある.今回の発見を礎として,ウイルスの感染が関連するこれらの疾患についての理解がさらに深まるものと期待している.
文 献
- Bakaletz, L. O.: Developing animal models for polymicrobial diseases. Nat. Rev. Microbiol., 2, 552-568 (2004)[PubMed]
- Trinchieri, G.: Type I interferon: friend or foe? J. Exp. Med., 207, 2053-2063 (2010)[PubMed]
- Iwasaki, A. & Medzhitov, R.: Regulation of adaptive immunity by the innate immune system. Science, 327, 291-295 (2010)[PubMed]
- Takeuchi, O. & Akira, S.: Pattern recognition receptors and inflammation. Cell, 140, 805-820 (2010)[PubMed]
- Honda, K. & Taniguchi, T.: IRFs: master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nat. Rev. Immunol., 6, 644-658 (2006)[PubMed]
- Janeway, C. A., Jr. & Medzhitov, R.: Innate immune recognition. Annu. Rev. Immunol., 20, 197-216 (2002)[PubMed]
- Steinman, R. M. & Hemmi, H.: Dendritic cells: translating innate to adaptive immunity. Curr. Top. Microbiol. Immunol., 311, 17-58 (2006)[PubMed]
- Blasius, A. L. & Beutler, B.: Intracellular toll-like receptors. Immunity, 32, 305-315 (2010)[PubMed]
- Yoneyama, M. & Fujita, T.: RNA recognition and signal transduction by RIG-I-like receptors. Immunol. Rev., 227, 54-65 (2009)[PubMed]
著者プロフィール
略歴:2006年 東京大学大学院医学系研究科 修了,同年 同 研究員,2008年 同 特任助教,2012年より東京大学生産技術研究所 特任助教.
研究テーマ:自然免疫応答の制御機構.
抱負:新しく解明した自然免疫応答の分子制御の機構を疾患の予防や治療に役立てたい.
柳井 秀元(Hideyuki Yanai)
東京大学生産技術研究所 特任助教.
中島 啓(Akira Nakajima)
京都大学医学系研究科 研究員.
谷口 維紹(Tadatsugu Taniguchi)
東京大学生産技術研究所 特任教授.
研究室URL:http://www.immunol.m.u-tokyo.ac.jp/page%20files/ja/top/top-ja.html
© 2012 根岸英雄・柳井秀元・中島 啓・谷口維紹 Licensed under CC 表示 2.1 日本
