抑制性の神経伝達物質GABAは小脳においてシナプスの刈り込みを制御する
中山寿子・狩野方伸
(東京大学大学院医学系研究科 神経生理学分野)
email:中山寿子,狩野方伸
DOI: 10.7875/first.author.2012.056
GABAergic inhibition regulates developmental synapse elimination in the cerebellum.
Hisako Nakayama, Taisuke Miyazaki, Kazuo Kitamura, Kouichi Hashimoto, Yuchio Yanagawa, Kunihiko Obata, Kenji Sakimura, Masahiko Watanabe, Masanobu Kano
Neuron, 74, 384-396 (2012)
生後まもない脳には過剰なシナプスが存在するが,やがて,必要なシナプスは強められ不要なシナプスは除去されて,機能的な神経回路が完成する.この過程は“シナプス刈り込み”とよばれ,神経回路の発達における普遍的な現象である.今回,筆者らは,小脳におけるシナプス刈り込みにおいて抑制性の神経伝達物質であるGABAのはたらきが必要であることを明らかにした.GABAの合成が低下するよう遺伝子改変されたマウスを用い,脳幹から小脳へ興奮性の信号を伝達する登上線維と小脳のプルキンエ細胞とのあいだのシナプスを調べた.その結果,このマウスでは生後10日ごろから生後16日ごろまでのあいだシナプス刈り込みが障害されており,このとき,GABAのはたらきを増強する薬を投与するとシナプス刈り込みは正常となることを確認した.さらに,この遺伝子改変マウスでは登上線維の入力に対してプルキンエ細胞の細胞体において,正常なマウスよりも大きなCa2+濃度の上昇の起こることが認められた.今回の結果から,発達期の小脳ではGABAによる抑制が細胞内のCa2+濃度を制御することにより,シナプス刈り込みに重要なはたらきをしていることが示唆された.
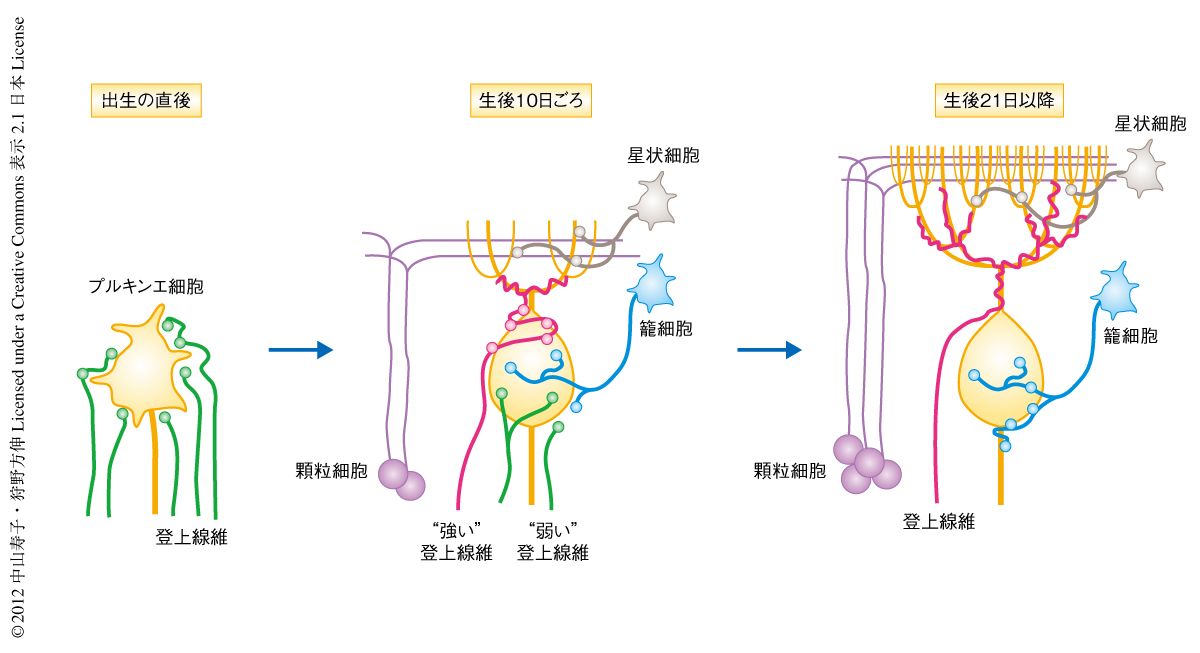
発達初期の動物の脳には過剰なシナプスが存在するが,生後の発達の過程において必要なシナプスだけが強められ不要なシナプスは除去されて,成熟した機能的な神経回路が完成する.この過程は“シナプス刈り込み”とよばれており,生後の発達期の神経回路にみられる普遍的な現象であると考えられている.中枢神経系においてシナプス刈り込みを定量的に評価できるのは,小脳の登上線維とプルキンエ細胞とのあいだのシナプスだけであり,この研究では,このシナプスの生後の変化を解析の対象とした.生まれたばかりの動物のプルキンエ細胞では,同じ程度の強さの信号を伝達する5本以上の登上線維がプルキンエ細胞の根元に相当する細胞体にシナプスを形成しているが,成熟した動物では強力な信号を伝達するわずか1本の登上線維が細胞体から大木の枝のように張り出した樹状突起にシナプスを形成している.このような単一の登上線維による支配は,つぎのような過程をへて獲得される.まず生後7日までに,細胞体にシナプスを形成していた複数の登上線維のうち1本だけが強化される(機能分化).つづく生後2~3週目には,選択的に強化された登上線維はプルキンエ細胞の樹状突起に侵入してシナプスを形成し(樹状突起移行),一方,生後1週目に強化されなかった弱い登上線維のシナプスはプルキンエ細胞の細胞体から除去される(シナプス刈り込み).その結果,1本の登上線維に由来するシナプスのみがプルキンエ細胞の樹状突起に残る1,2)(図1).
これまでの研究から,正常なシナプス刈り込みにはニューロンがまわりの環境などの刺激をうけて電気的に活動することが必要であることが知られていた3,4).グルタミン酸はニューロンの活動を上昇させる興奮性の神経伝達物質であるが,グルタミン酸を受け取る受容体のうちある種のものが欠落するとシナプス刈り込みはうまく起こらない5,6).一方,γ-アミノ酪酸(GABA)はニューロンの活動を抑える抑制性の神経伝達物質でありニューロンの活動の制御に重要なはたらきをしている.しかし,GABAが神経活動に依存的なシナプス刈り込みに影響するかどうかについてはこれまで不明であった.今回の研究では,GABAを合成する酵素のひとつであるGAD67の遺伝子をヘテロ欠損する遺伝子改変マウス(GAD67-GFPノックインマウス)を解析することにより,GABAによる抑制性のシナプス伝達がシナプス刈り込みに重要なはたらきをすることを発見した.
まず,GAD67遺伝子のヘテロ欠損によりGABA作動性のシナプス伝達が減弱するかどうかを調べた.さまざまな日齢のマウスの小脳から急性切片標本を作製し,ホールセルパッチクランプ法を用いて,プルキンエ細胞から膜電位を固定した状態でGABA作動性の微小シナプス後電流を測定した.その結果,GAD67ヘテロ欠損マウスでは,生後1週目においてはGABA作動性の電流の振幅について野生型マウスに対し有意な差は認められなかったが,生後2週目においてはこの電流の振幅は有意に小さいという結果が得られた.GABAは成熟動物のニューロンに対しては神経活動を抑えるように(抑制性に)作用するが,プルキンエ細胞を含め発達の初期のニューロンに対してはむしろ神経活動を亢進させるように(興奮性に)作用する7).そこで,GABA作動性のシナプス伝達に異常が認められた生後2週目において,GABAの作用がプルキンエ細胞に対し興奮性か抑制性かを調べた.その結果,GABA受容体の活性化によりプルキンエ細胞の興奮は抑制されることを確認した.すなわち,GAD67ヘテロ欠損マウスでは生後2週目においてプルキンエ細胞へのGABAによる抑制性のシナプス伝達が減弱していることがわかった.
個々のプルキンエ細胞からホールセル記録を行い,登上線維を1本ずつ別々に電気刺激したときにひき起こされるシナプス後電流を測定することにより,プルキンエ細胞にどの程度の強さの,そして,何本の登上線維が結合しているかを調べた.その結果,GAD67ヘテロ欠損マウスでは,生後9日ごろまでのシナプス刈り込みは正常に起こるが,生後10日目以降のシナプス刈り込みの過程は障害されており,野生型マウスでは登上線維の単一支配が完成している時期においても,約半数のプルキンエ細胞が2本以上の登上線維からのシナプス伝達をうけていることが明らかになった.すなわち,GAD67ヘテロ欠損マウスでは生後2週目においてプルキンエ細胞へのGABA作動性のシナプス伝達が減弱しており,同じ時期の登上線維におけるシナプス刈り込みも障害されていた.
GAD67ヘテロ欠損マウスは小脳以外の脳領域においてもGAD67遺伝子がヘテロ欠損しているため,さきに述べた異常が小脳皮質におけるGAD67遺伝子の改変に起因するのか,あるいは,小脳以外の脳領域におけるGAD67遺伝子の改変に起因するのかは明らかでなかった.そこで,遺伝子改変をしていない野生型マウスの小脳皮質に対し,GAD67のはたらきを阻害する3-MPという薬物を生後10日から慢性投与してGABAの合成を抑制した場合に,シナプス刈り込みがどのような影響をうけるのかを調べたところ,GAD67ヘテロ欠損マウスと同様にシナプス刈り込みに異常が起こった.一方,GAD67ヘテロ欠損マウスの小脳皮質にGABAのはたらきを増強するDiazepamという薬物を生後10日から投与したところシナプス刈り込みは正常化した.しかしながら,Diazepamを生後17日から投与した場合にはGAD67ヘテロ欠損マウスのシナプス刈り込みの異常は正常化しなかった.したがって,生後10日ごろから生後16日ごろまでのあいだ,小脳においてGABAが適正にはたらくことがシナプス刈り込みに必須であることが明らかになった.
小脳皮質の神経回路のなかで,どのGABA作動性のシナプス伝達が登上線維におけるシナプス刈り込みに必要なのかを調べた.プルキンエ細胞は籠細胞および星状細胞からGABA作動性のシナプス伝達を受け取っているが,籠細胞はプルキンエ細胞の細胞体に,星状細胞はプルキンエ細胞の樹状突起に,というように,両者はプルキンエ細胞の異なる部位にシナプスを形成している.シナプスの部位の違いはシナプス電流の波形の差として現われる.これを利用し,プルキンエ細胞で記録されるGABA作動性の電流の波形を詳細に解析することにより,GAD67ヘテロ欠損マウスで減弱しているGABA作動性のシナプス伝達はプルキンエ細胞の細胞体に形成されたシナプスに由来する,すなわち,籠細胞に由来する可能性の高いことを見い出した.この可能性を検証するため,籠細胞とプルキンエ細胞から同時にホールセル記録を行い,単一の籠細胞を刺激したときにプルキンエ細胞に誘発されるGABA作動性のシナプス後電流を測定して,GAD67ヘテロ欠損マウスと野生型マウスとのあいだで比較した.その結果,単一の籠細胞に由来するシナプス後電流はGAD67ヘテロ欠損マウスにおいて有意に小さく,籠細胞がプルキンエ細胞の細胞体に形成するGABA作動性の抑制性シナプスが重要であることがわかった.
GABA作動性のシナプス伝達がどのような機構で登上線維におけるシナプス刈り込みに関与しているのかを検討した.登上線維とプルキンエ細胞とのあいだのシナプスの正常な発達にはプルキンエ細胞に発現する電位依存性Ca2+チャネルが必要であるという先行研究がある8).そこで,登上線維の活性化によりプルキンエ細胞の細胞体に発生するCa2+流入の大きさがGABA作動性のシナプス伝達の強さにより制御されるという可能性を考え検討した.Ca2+濃度が上昇すると蛍光強度の上昇する色素をプルキンエ細胞に注入し,登上線維を電気刺激したときに細胞体において生じる蛍光強度の変化,すなわち,Ca2+濃度の変化をGABA作動性のシナプス伝達が減弱しているGAD67ヘテロ欠損マウスと野生型マウスとで比較した.その結果,強い登上線維を刺激したときのCa2+濃度の上昇には有意な差は認められなかったが,弱い登上線維を刺激したときのプルキンエ細胞の細胞体におけるCa2+濃度の上昇はGAD67ヘテロ欠損マウスのほうが有意に大きいことがわかった.また,GAD67ヘテロ欠損マウスにGABAのはたらきを増強するDiazepamを投与してGABA作動性のシナプス伝達を増強するとCa2+濃度の上昇が抑制された.これらの結果から,弱い登上線維の入力に対するプルキンエ細胞の細胞体におけるCa2+濃度の上昇の大きさはGABA作動性のシナプス伝達により制御されており,GAD67ヘテロ欠損マウスではGABA作動性のシナプス伝達が減弱しているため野生型マウスよりも大きなCa2+濃度の上昇の起こることが明らかになった.
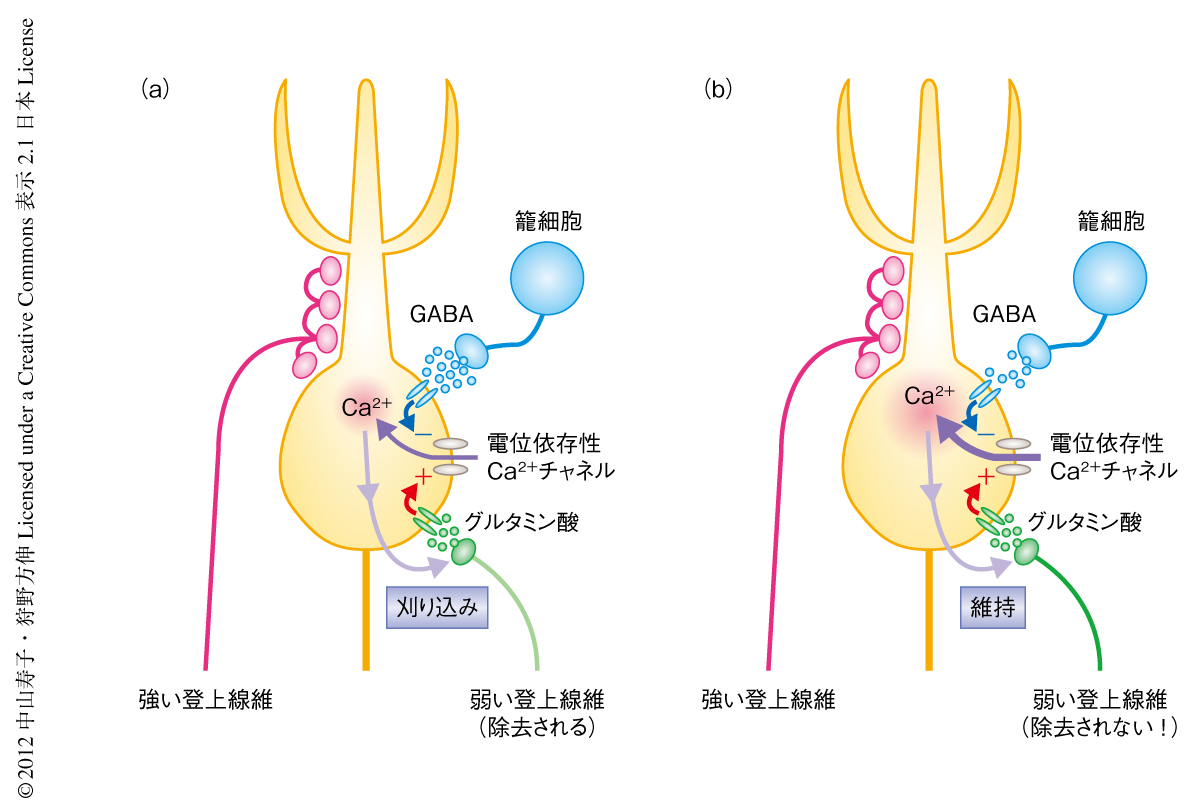
今回の研究により,小脳の登上線維とプルキンエ細胞とのあいだのシナプス刈り込みにおいて,籠細胞からプルキンエ細胞の細胞体へのGABA作動性のシナプス伝達が重要であることがはじめて明らかになった.今回の結果をうけて,GABAが登上線維におけるシナプス刈り込みを制御する機構に関して,以下のようなモデルを提唱する(図2).生後10日ごろから生後16日ごろは,プルキンエ細胞の細胞体では登上線維に由来する興奮性シナプスが減少し,反対に,籠細胞に由来するGABA作動性の抑制性シナプスが増加する時期である9).この時期には,籠細胞から放出されるGABAによりプルキンエ細胞の細胞体の活動が抑制されるため,登上線維から興奮性の入力が伝達されても細胞体の活動は高くなることはできない.すると,電位依存性Ca2+チャネルが十分に開口することができず,細胞体に流入するCa2+は少なくなる.十分な量のCa2+がないとシナプスを維持することができないため,やがて,プルキンエ細胞の細胞体にシナプスをもつ弱い登上線維は除去されてしまうのに対し,プルキンエ細胞の樹状突起にシナプスをもつ登上線維は維持され,成熟型の神経回路が獲得される.GAD67ヘテロ欠損マウスのようになんらかの原因でGABA作動性のシナプス伝達が減弱してしまうと,弱い登上線維においてもプルキンエ細胞の細胞体にシナプスを維持するのに十分な量のCa2+が流入してしまうため,本来はシナプス刈り込みをうけるべきシナプスが維持されてしまうと考えられる.プルキンエ細胞の細胞体へ流入するCa2+の量に依存してシナプス刈り込みが制御されているとすれば,Ca2+とシナプス刈り込みあるいはシナプス維持とを結ぶ機構とは何かを解明することが今後の重要な研究課題のひとつである.
これまで,登上線維におけるシナプス刈り込みに関する研究のほとんどは興奮性のシナプス伝達とその下流のシグナルタンパク質に着目して行われてきたが,生体における情報の伝達は興奮と抑制との絶妙なバランスにより成立しているはずであるから,GABA作動性の抑制性シナプスと興奮性シナプスの刈り込みとに着目したこの研究は,生体における神経回路の形成についての理解を格段に深めることが期待される.また,自閉症や統合失調症などの疾患でみられる社会性障害や情緒障害の原因となる細胞レベルの障害は,特定の神経系での興奮性活動と抑制性活動とのバランスの乱れにより起こるというモデルもある.また,統合失調症や自閉症の病因として神経回路の発達異常があり,発達の特定の時期に起こるシナプス刈り込みの異常の関与も考えられている.この研究で見い出されたGABAとシナプス刈り込みとの関連はあくまでマウスにおけるものではあるが,今後,モデル動物やヒトにおける研究がより進むことにより統合失調症や自閉症の病因の理解にもつながることが期待される.
2003年 大阪大学大学院基礎工学研究科博士後期課程 単位修得退学,2004年 大阪大学大学院基礎工学研究科にて博士号(理学)取得,同年 金沢大学大学院医学系研究科 研究員,2005年 大阪大学大学院医学系研究科 研究員,2007年 東京大学大学院医学系研究科 研究員を経て,2011年より広島大学大学院医歯薬保健学研究科 助教.
狩野 方伸(Masanobu Kano)
東京大学大学院医学系研究科 教授.
研究室URL:http://plaza.umin.ac.jp/~neurophy/Kano_Lab_j/Top_j.html
© 2012 中山寿子・狩野方伸 Licensed under CC 表示 2.1 日本
(東京大学大学院医学系研究科 神経生理学分野)
email:中山寿子,狩野方伸
DOI: 10.7875/first.author.2012.056
GABAergic inhibition regulates developmental synapse elimination in the cerebellum.
Hisako Nakayama, Taisuke Miyazaki, Kazuo Kitamura, Kouichi Hashimoto, Yuchio Yanagawa, Kunihiko Obata, Kenji Sakimura, Masahiko Watanabe, Masanobu Kano
Neuron, 74, 384-396 (2012)
この論文に出現する遺伝子・タンパク質のUniprot ID
GAD67(P48318), GABA受容体
要 約
生後まもない脳には過剰なシナプスが存在するが,やがて,必要なシナプスは強められ不要なシナプスは除去されて,機能的な神経回路が完成する.この過程は“シナプス刈り込み”とよばれ,神経回路の発達における普遍的な現象である.今回,筆者らは,小脳におけるシナプス刈り込みにおいて抑制性の神経伝達物質であるGABAのはたらきが必要であることを明らかにした.GABAの合成が低下するよう遺伝子改変されたマウスを用い,脳幹から小脳へ興奮性の信号を伝達する登上線維と小脳のプルキンエ細胞とのあいだのシナプスを調べた.その結果,このマウスでは生後10日ごろから生後16日ごろまでのあいだシナプス刈り込みが障害されており,このとき,GABAのはたらきを増強する薬を投与するとシナプス刈り込みは正常となることを確認した.さらに,この遺伝子改変マウスでは登上線維の入力に対してプルキンエ細胞の細胞体において,正常なマウスよりも大きなCa2+濃度の上昇の起こることが認められた.今回の結果から,発達期の小脳ではGABAによる抑制が細胞内のCa2+濃度を制御することにより,シナプス刈り込みに重要なはたらきをしていることが示唆された.
はじめに
発達初期の動物の脳には過剰なシナプスが存在するが,生後の発達の過程において必要なシナプスだけが強められ不要なシナプスは除去されて,成熟した機能的な神経回路が完成する.この過程は“シナプス刈り込み”とよばれており,生後の発達期の神経回路にみられる普遍的な現象であると考えられている.中枢神経系においてシナプス刈り込みを定量的に評価できるのは,小脳の登上線維とプルキンエ細胞とのあいだのシナプスだけであり,この研究では,このシナプスの生後の変化を解析の対象とした.生まれたばかりの動物のプルキンエ細胞では,同じ程度の強さの信号を伝達する5本以上の登上線維がプルキンエ細胞の根元に相当する細胞体にシナプスを形成しているが,成熟した動物では強力な信号を伝達するわずか1本の登上線維が細胞体から大木の枝のように張り出した樹状突起にシナプスを形成している.このような単一の登上線維による支配は,つぎのような過程をへて獲得される.まず生後7日までに,細胞体にシナプスを形成していた複数の登上線維のうち1本だけが強化される(機能分化).つづく生後2~3週目には,選択的に強化された登上線維はプルキンエ細胞の樹状突起に侵入してシナプスを形成し(樹状突起移行),一方,生後1週目に強化されなかった弱い登上線維のシナプスはプルキンエ細胞の細胞体から除去される(シナプス刈り込み).その結果,1本の登上線維に由来するシナプスのみがプルキンエ細胞の樹状突起に残る1,2)(図1).
これまでの研究から,正常なシナプス刈り込みにはニューロンがまわりの環境などの刺激をうけて電気的に活動することが必要であることが知られていた3,4).グルタミン酸はニューロンの活動を上昇させる興奮性の神経伝達物質であるが,グルタミン酸を受け取る受容体のうちある種のものが欠落するとシナプス刈り込みはうまく起こらない5,6).一方,γ-アミノ酪酸(GABA)はニューロンの活動を抑える抑制性の神経伝達物質でありニューロンの活動の制御に重要なはたらきをしている.しかし,GABAが神経活動に依存的なシナプス刈り込みに影響するかどうかについてはこれまで不明であった.今回の研究では,GABAを合成する酵素のひとつであるGAD67の遺伝子をヘテロ欠損する遺伝子改変マウス(GAD67-GFPノックインマウス)を解析することにより,GABAによる抑制性のシナプス伝達がシナプス刈り込みに重要なはたらきをすることを発見した.
1.GAD67ヘテロ欠損マウスでは生後2週目においてGABA作動性のシナプス伝達が減弱している
まず,GAD67遺伝子のヘテロ欠損によりGABA作動性のシナプス伝達が減弱するかどうかを調べた.さまざまな日齢のマウスの小脳から急性切片標本を作製し,ホールセルパッチクランプ法を用いて,プルキンエ細胞から膜電位を固定した状態でGABA作動性の微小シナプス後電流を測定した.その結果,GAD67ヘテロ欠損マウスでは,生後1週目においてはGABA作動性の電流の振幅について野生型マウスに対し有意な差は認められなかったが,生後2週目においてはこの電流の振幅は有意に小さいという結果が得られた.GABAは成熟動物のニューロンに対しては神経活動を抑えるように(抑制性に)作用するが,プルキンエ細胞を含め発達の初期のニューロンに対してはむしろ神経活動を亢進させるように(興奮性に)作用する7).そこで,GABA作動性のシナプス伝達に異常が認められた生後2週目において,GABAの作用がプルキンエ細胞に対し興奮性か抑制性かを調べた.その結果,GABA受容体の活性化によりプルキンエ細胞の興奮は抑制されることを確認した.すなわち,GAD67ヘテロ欠損マウスでは生後2週目においてプルキンエ細胞へのGABAによる抑制性のシナプス伝達が減弱していることがわかった.
2.GAD67ヘテロ欠損マウスでは生後2週目においてシナプス刈り込みが障害されている
個々のプルキンエ細胞からホールセル記録を行い,登上線維を1本ずつ別々に電気刺激したときにひき起こされるシナプス後電流を測定することにより,プルキンエ細胞にどの程度の強さの,そして,何本の登上線維が結合しているかを調べた.その結果,GAD67ヘテロ欠損マウスでは,生後9日ごろまでのシナプス刈り込みは正常に起こるが,生後10日目以降のシナプス刈り込みの過程は障害されており,野生型マウスでは登上線維の単一支配が完成している時期においても,約半数のプルキンエ細胞が2本以上の登上線維からのシナプス伝達をうけていることが明らかになった.すなわち,GAD67ヘテロ欠損マウスでは生後2週目においてプルキンエ細胞へのGABA作動性のシナプス伝達が減弱しており,同じ時期の登上線維におけるシナプス刈り込みも障害されていた.
3.小脳皮質におけるGABA作動性のシナプス伝達はシナプス刈り込みを制御する
GAD67ヘテロ欠損マウスは小脳以外の脳領域においてもGAD67遺伝子がヘテロ欠損しているため,さきに述べた異常が小脳皮質におけるGAD67遺伝子の改変に起因するのか,あるいは,小脳以外の脳領域におけるGAD67遺伝子の改変に起因するのかは明らかでなかった.そこで,遺伝子改変をしていない野生型マウスの小脳皮質に対し,GAD67のはたらきを阻害する3-MPという薬物を生後10日から慢性投与してGABAの合成を抑制した場合に,シナプス刈り込みがどのような影響をうけるのかを調べたところ,GAD67ヘテロ欠損マウスと同様にシナプス刈り込みに異常が起こった.一方,GAD67ヘテロ欠損マウスの小脳皮質にGABAのはたらきを増強するDiazepamという薬物を生後10日から投与したところシナプス刈り込みは正常化した.しかしながら,Diazepamを生後17日から投与した場合にはGAD67ヘテロ欠損マウスのシナプス刈り込みの異常は正常化しなかった.したがって,生後10日ごろから生後16日ごろまでのあいだ,小脳においてGABAが適正にはたらくことがシナプス刈り込みに必須であることが明らかになった.
4.籠細胞からプルキンエ細胞の細胞体へのGABA作動性のシナプス伝達が登上線維におけるシナプス刈り込みに重要である
小脳皮質の神経回路のなかで,どのGABA作動性のシナプス伝達が登上線維におけるシナプス刈り込みに必要なのかを調べた.プルキンエ細胞は籠細胞および星状細胞からGABA作動性のシナプス伝達を受け取っているが,籠細胞はプルキンエ細胞の細胞体に,星状細胞はプルキンエ細胞の樹状突起に,というように,両者はプルキンエ細胞の異なる部位にシナプスを形成している.シナプスの部位の違いはシナプス電流の波形の差として現われる.これを利用し,プルキンエ細胞で記録されるGABA作動性の電流の波形を詳細に解析することにより,GAD67ヘテロ欠損マウスで減弱しているGABA作動性のシナプス伝達はプルキンエ細胞の細胞体に形成されたシナプスに由来する,すなわち,籠細胞に由来する可能性の高いことを見い出した.この可能性を検証するため,籠細胞とプルキンエ細胞から同時にホールセル記録を行い,単一の籠細胞を刺激したときにプルキンエ細胞に誘発されるGABA作動性のシナプス後電流を測定して,GAD67ヘテロ欠損マウスと野生型マウスとのあいだで比較した.その結果,単一の籠細胞に由来するシナプス後電流はGAD67ヘテロ欠損マウスにおいて有意に小さく,籠細胞がプルキンエ細胞の細胞体に形成するGABA作動性の抑制性シナプスが重要であることがわかった.
5.登上線維の入力に対し発生するプルキンエ細胞におけるCa2+濃度の上昇は籠細胞からのGABA作動性のシナプス伝達により制御される
GABA作動性のシナプス伝達がどのような機構で登上線維におけるシナプス刈り込みに関与しているのかを検討した.登上線維とプルキンエ細胞とのあいだのシナプスの正常な発達にはプルキンエ細胞に発現する電位依存性Ca2+チャネルが必要であるという先行研究がある8).そこで,登上線維の活性化によりプルキンエ細胞の細胞体に発生するCa2+流入の大きさがGABA作動性のシナプス伝達の強さにより制御されるという可能性を考え検討した.Ca2+濃度が上昇すると蛍光強度の上昇する色素をプルキンエ細胞に注入し,登上線維を電気刺激したときに細胞体において生じる蛍光強度の変化,すなわち,Ca2+濃度の変化をGABA作動性のシナプス伝達が減弱しているGAD67ヘテロ欠損マウスと野生型マウスとで比較した.その結果,強い登上線維を刺激したときのCa2+濃度の上昇には有意な差は認められなかったが,弱い登上線維を刺激したときのプルキンエ細胞の細胞体におけるCa2+濃度の上昇はGAD67ヘテロ欠損マウスのほうが有意に大きいことがわかった.また,GAD67ヘテロ欠損マウスにGABAのはたらきを増強するDiazepamを投与してGABA作動性のシナプス伝達を増強するとCa2+濃度の上昇が抑制された.これらの結果から,弱い登上線維の入力に対するプルキンエ細胞の細胞体におけるCa2+濃度の上昇の大きさはGABA作動性のシナプス伝達により制御されており,GAD67ヘテロ欠損マウスではGABA作動性のシナプス伝達が減弱しているため野生型マウスよりも大きなCa2+濃度の上昇の起こることが明らかになった.
6.GABAが登上線維におけるシナプス刈り込みを制御する機構
今回の研究により,小脳の登上線維とプルキンエ細胞とのあいだのシナプス刈り込みにおいて,籠細胞からプルキンエ細胞の細胞体へのGABA作動性のシナプス伝達が重要であることがはじめて明らかになった.今回の結果をうけて,GABAが登上線維におけるシナプス刈り込みを制御する機構に関して,以下のようなモデルを提唱する(図2).生後10日ごろから生後16日ごろは,プルキンエ細胞の細胞体では登上線維に由来する興奮性シナプスが減少し,反対に,籠細胞に由来するGABA作動性の抑制性シナプスが増加する時期である9).この時期には,籠細胞から放出されるGABAによりプルキンエ細胞の細胞体の活動が抑制されるため,登上線維から興奮性の入力が伝達されても細胞体の活動は高くなることはできない.すると,電位依存性Ca2+チャネルが十分に開口することができず,細胞体に流入するCa2+は少なくなる.十分な量のCa2+がないとシナプスを維持することができないため,やがて,プルキンエ細胞の細胞体にシナプスをもつ弱い登上線維は除去されてしまうのに対し,プルキンエ細胞の樹状突起にシナプスをもつ登上線維は維持され,成熟型の神経回路が獲得される.GAD67ヘテロ欠損マウスのようになんらかの原因でGABA作動性のシナプス伝達が減弱してしまうと,弱い登上線維においてもプルキンエ細胞の細胞体にシナプスを維持するのに十分な量のCa2+が流入してしまうため,本来はシナプス刈り込みをうけるべきシナプスが維持されてしまうと考えられる.プルキンエ細胞の細胞体へ流入するCa2+の量に依存してシナプス刈り込みが制御されているとすれば,Ca2+とシナプス刈り込みあるいはシナプス維持とを結ぶ機構とは何かを解明することが今後の重要な研究課題のひとつである.
おわりに
これまで,登上線維におけるシナプス刈り込みに関する研究のほとんどは興奮性のシナプス伝達とその下流のシグナルタンパク質に着目して行われてきたが,生体における情報の伝達は興奮と抑制との絶妙なバランスにより成立しているはずであるから,GABA作動性の抑制性シナプスと興奮性シナプスの刈り込みとに着目したこの研究は,生体における神経回路の形成についての理解を格段に深めることが期待される.また,自閉症や統合失調症などの疾患でみられる社会性障害や情緒障害の原因となる細胞レベルの障害は,特定の神経系での興奮性活動と抑制性活動とのバランスの乱れにより起こるというモデルもある.また,統合失調症や自閉症の病因として神経回路の発達異常があり,発達の特定の時期に起こるシナプス刈り込みの異常の関与も考えられている.この研究で見い出されたGABAとシナプス刈り込みとの関連はあくまでマウスにおけるものではあるが,今後,モデル動物やヒトにおける研究がより進むことにより統合失調症や自閉症の病因の理解にもつながることが期待される.
文 献
- Hashimoto, K. & Kano, M.: Functional differentiation of multiple climbing fiber inputs during synapse elimination in the developing cerebellum. Neuron, 38, 785-796 (2003)[PubMed]
- Hashimoto, K., Ichikawa, R., Kitamura, K. et al.: Translocation of a "winner" climbing fiber to the Purkinje cell dendrite and subsequent elimination of "losers" from the soma in developing cerebellum. Neuron, 63, 106-118 (2009)[PubMed]
- Andjus, P. R., Zhu, L., Cesa, R. et al.: A change in the pattern of activity affects the developmental regression of the Purkinje cell polyinnervation by climbing fibers in the rat cerebellum. Neuroscience, 121, 563-572 (2003)[PubMed]
- Lorenzetto, E., Caselli, L., Feng, G. et al.: Genetic perturbation of postsynaptic activity regulates synapse elimination in developing cerebellum. Proc. Natl. Acad. Sci. USA, 106, 16475-16480 (2009)[PubMed]
- Kano, M. & Hashimoto, K.: Synapse elimination in the central nervous system. Curr. Opin. Neurobiol., 19, 154-161 (2009)[PubMed]
- Watanabe, M. & Kano, M.: Climbing fiber synapse elimination in cerebellar Purkinje cells. Eur. J. Neurosci., 34, 1697-1710 (2011)[PubMed]
- Eilers, J., Plant, T. D., Marandi, N. et al.: GABA-mediated Ca2+ signalling in developing rat cerebellar Purkinje neurones. J. Physiol., 536, 429-437 (2001)[PubMed]
- Hashimoto, K., Tsujita, M., Miyazaki, T. et al.: Postsynaptic P/Q-type Ca2+ channel in Purkinje cell mediates synaptic competition and elimination in developing cerebellum. Proc. Natl. Acad. Sci. USA, 108, 9987-9992 (2011)[PubMed]
- Ichikawa, R., Yamasaki, M., Miyazaki, T. et al.: Developmental switching of perisomatic innervation from climbing fibers to basket cell fibers in cerebellar Purkinje cells. J. Neurosci., 31, 16916-16127 (2011)[PubMed]
著者プロフィール
2003年 大阪大学大学院基礎工学研究科博士後期課程 単位修得退学,2004年 大阪大学大学院基礎工学研究科にて博士号(理学)取得,同年 金沢大学大学院医学系研究科 研究員,2005年 大阪大学大学院医学系研究科 研究員,2007年 東京大学大学院医学系研究科 研究員を経て,2011年より広島大学大学院医歯薬保健学研究科 助教.
狩野 方伸(Masanobu Kano)
東京大学大学院医学系研究科 教授.
研究室URL:http://plaza.umin.ac.jp/~neurophy/Kano_Lab_j/Top_j.html
© 2012 中山寿子・狩野方伸 Licensed under CC 表示 2.1 日本
