“分子のものさし”がRNAの長さを測り仕分けする
大野 睦人
(京都大学ウイルス研究所 情報高分子化学研究分野)
email:大野睦人
DOI: 10.7875/first.author.2012.046
HnRNP C tetramer measures RNA length to classify RNA polymerase II transcripts for export.
Asako McCloskey, Ichiro Taniguchi, Kaori Shinmyozu, Mutsuhito Ohno
Science, 335, 1643-1646 (2012)
通常,RNAの特異的な認識はその特異的な塩基配列や構造を介し行われる.しかし,筆者らは,核において“RNAの長さ”が認識される分子機構を発見した.RNAポリメラーゼIIはmRNAを合成するが,ほかにもU snRNAなど短いRNAも合成する.しかし,転写開始の直後にはその転写産物が将来mRNAになるのかU snRNAになるのかはわからない.転写がさらに進み新生RNAの長さが200~300塩基長より長くなると,hnRNP Cという核内RNA結合タンパク質のヘテロ四量体が安定に結合できるようになることで,U snRNAの輸送タンパク質の結合が阻害されその転写産物はmRNA前駆体であると認定される.逆に,RNAの長さが200~300塩基長より短いまま転写が終了した場合,hnRNP Cヘテロ四量体が安定に結合できずその転写産物はU snRNA前駆体であると認定される.このように,hnRNP Cヘテロ四量体が“分子のものさし”となってRNAの長さを測り,RNAをその長さに応じて仕分けしていた.
真核生物ではほとんどのRNAは核において合成されるが,そののちの行き先はRNA種ごとに決まっている.多くのRNAは核から細胞質へと輸送されるが,そのようなRNAの主要なものとしては,リボソームRNA,tRNA,スプライソソームに含まれるウリジンに富む核内低分子RNA(U small nuclear RNA:U snRNA),mRNA,マイクロRNAなどの前駆体があげられる1).これらのRNAは核においてそれぞれのRNA種に固有の輸送タンパク質が結合したのち細胞質へと輸送される.核においてそれぞれのRNAに結合する輸送タンパク質は核外輸送をつかさどるだけでなく,そののちのRNAの運命(局在化,翻訳,安定性など)をも規定することが明らかになってきた.このことは,それぞれのRNA種は核においてすでに輸送タンパク質により識別されていることを意味する.しかし,それぞれのRNA種のいかなる特徴が核において識別されているのか,その識別を行う因子は何か,という点に関しては未知な部分が多い.
多くのRNA種のなかでもU snRNAとmRNAはいずれもRNAポリメラーゼIIにより合成され,5’末端にm7G型のキャップ構造が共通に形成されて,そこにはキャップ構造結合タンパク質複合体が結合する.しかし,このあとの核外輸送複合体の形成は両者でまったく違っている1).U snRNAの場合は,PHAX(phosphorylated adaptor for RNA export)というアダプタータンパク質がキャップ構造結合タンパク質複合体とその近傍のRNAに結合することによりRNAに呼び込まれ,そのPHAXが今度は核外輸送シグナル(nuclear export signal:NES)の輸送タンパク質CRM1-RanGTPを呼び込みU snRNA核外輸送複合体が完成する2).これに対して,mRNAの場合は,Aly/REFなどのRNA結合性のアダプタータンパク質がしばしばTREX(transcription/export)複合体やエキソンジャンクション複合体といった巨大複合体の一部としてmRNAに結合し,それがmRNAの主要な輸送タンパク質であるTAP/NXF1をRNAに呼び込む.このように,U snRNAとmRNAはある意味で似た者どうしではあるが,輸送タンパク質がきちんと区別できるような特徴をもっているはずである.
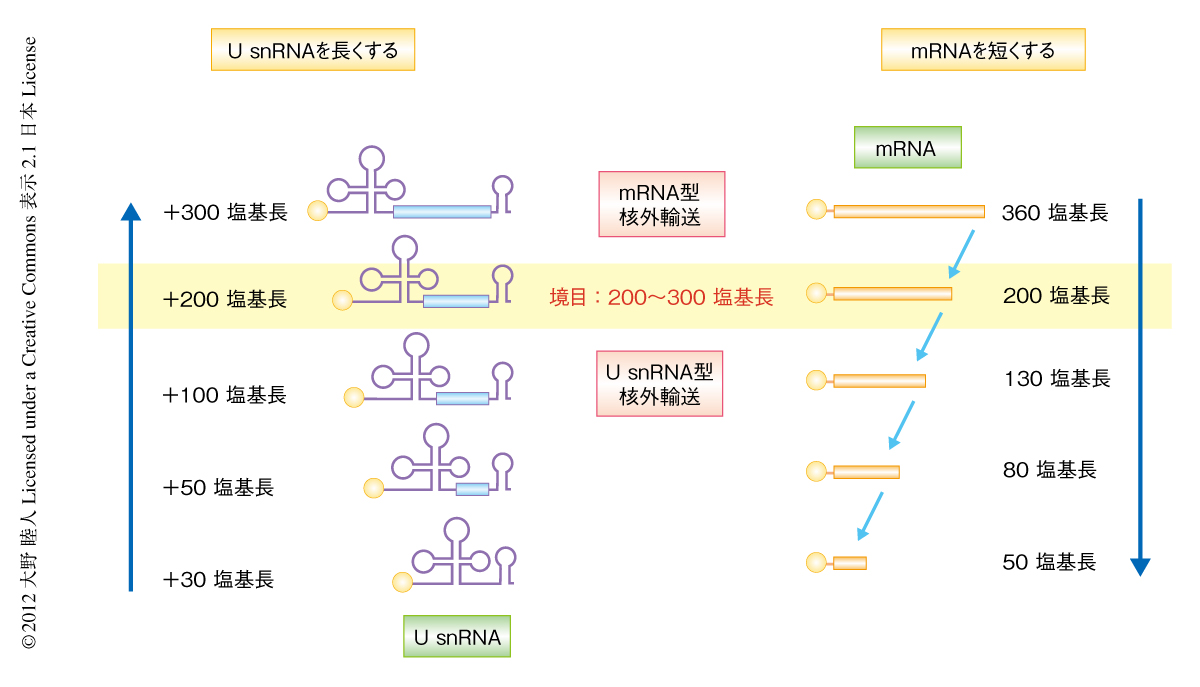
筆者らは,そのような特徴を探索した結果,“RNAの長さ”という概念にたどりついた.m7G型のキャップ構造をもつRNAは塩基配列ではなくその長さに応じてU snRNAの機構あるいはmRNAの機構により核外輸送されることを発見した.すなわち,人工的に長くしたU snRNAはmRNAの輸送タンパク質により,逆に,人工的に短くしたmRNAはU snRNAの輸送タンパク質より核から細胞質へと輸送されるようになったのである3,4).また,その切り替わりの長さの境目は約200~300塩基長であった5)(図1).ただし,強固な高次構造をもつRNA領域には長さの効果が表われないこともわかった3-5).このことは,細胞にはRNAの長さを測り輸送経路を決定する分子機構の存在することを意味していたが,その詳細については長いあいだ謎であった.
この問題に接近するため,RNAの長さに応じたRNA結合タンパク質の変化を試験管内で再現することを試みた.試験管内で合成したさまざまな長さのRNAと大腸菌により産生させ精製した組換えキャップ構造結合タンパク質複合体と組換えGST-PHAX融合タンパク質とを混合し,GSTのプルダウンを行うことによりU snRNA核外輸送複合体を部分的に模擬するRNA-キャップ構造結合タンパク質複合体-PHAX三者複合体の形成を調べた.その結果,この三者複合体はRNAの長さにかかわらずm7G型のキャップ構造をもつすべてのRNAにおいて形成された.この結果は,この精製系ではin vivoにおいてみられたRNAの長さに応じたRNA核外輸送複合体の形成がみられないことを意味した.しかし,HeLa細胞の核抽出液をこの精製系にくわえると,in vivoと同様に,長いRNAへのPHAXの結合が特異的に阻害された.このことは,核にはRNAの長さを測り長いRNAへのPHAXの結合を阻害する因子の存在することを示唆していた.
このPHAX呼び込み阻害活性をHeLa細胞の核抽出液からさまざまなカラムクロマトグラフィーを用いて精製した.最終精製画分にはこの活性と完全に相関する2本のメジャーなバンドがみられた.質量分析の結果,これらのバンドはヘテロ核内リボタンパク質(heterogeneous nuclear ribonucleoprotein:hnRNP)のC1とC2であることがわかった.hnRNP C1とhnRNP C2は3対1の割合でヘテロ四量体を形成する非常に量の多い核内RNA結合タンパク質であり,転写されつつあるmRNA前駆体につぎつぎと連続して結合しているものと考えられている6).
大腸菌で産生したhnRNP Cヘテロ四量体は試験管内において200~300塩基長以上の長さのRNAと特異的に結合しPHAXの結合を阻害した.これは,hnRNP Cヘテロ四量体がキャップ構造結合タンパク質複合体とRNAの両方に結合することによりPHAXの結合を競合的に阻害するためであることもわかった.RNAi法によりHeLa細胞においてhnRNP C1およびhnRNP C2をノックダウンすると,本来はmRNAに結合しないはずのPHAXがmRNAにも結合することが観察され,同時に,mRNAの核から細胞質への輸送が阻害された.これはおそらく,PHAXが結合したmRNAが異常なRNAであると認識され核外輸送が停止させられたのであろう.この考えと合致するように,hnRNP C1およびhnRNP C2とともにPHAXをノックダウンするとmRNAの核から細胞質への輸送が回復した.これらの結果から,細胞においてもこのhnRNP Cヘテロ四量体がPHAXとmRNAとの結合を防いでおり,そうすることでmRNAに適正な核外輸送複合体が形成されmRNAの核外輸送がとどこおりなく行われるよう管理していることが示された.
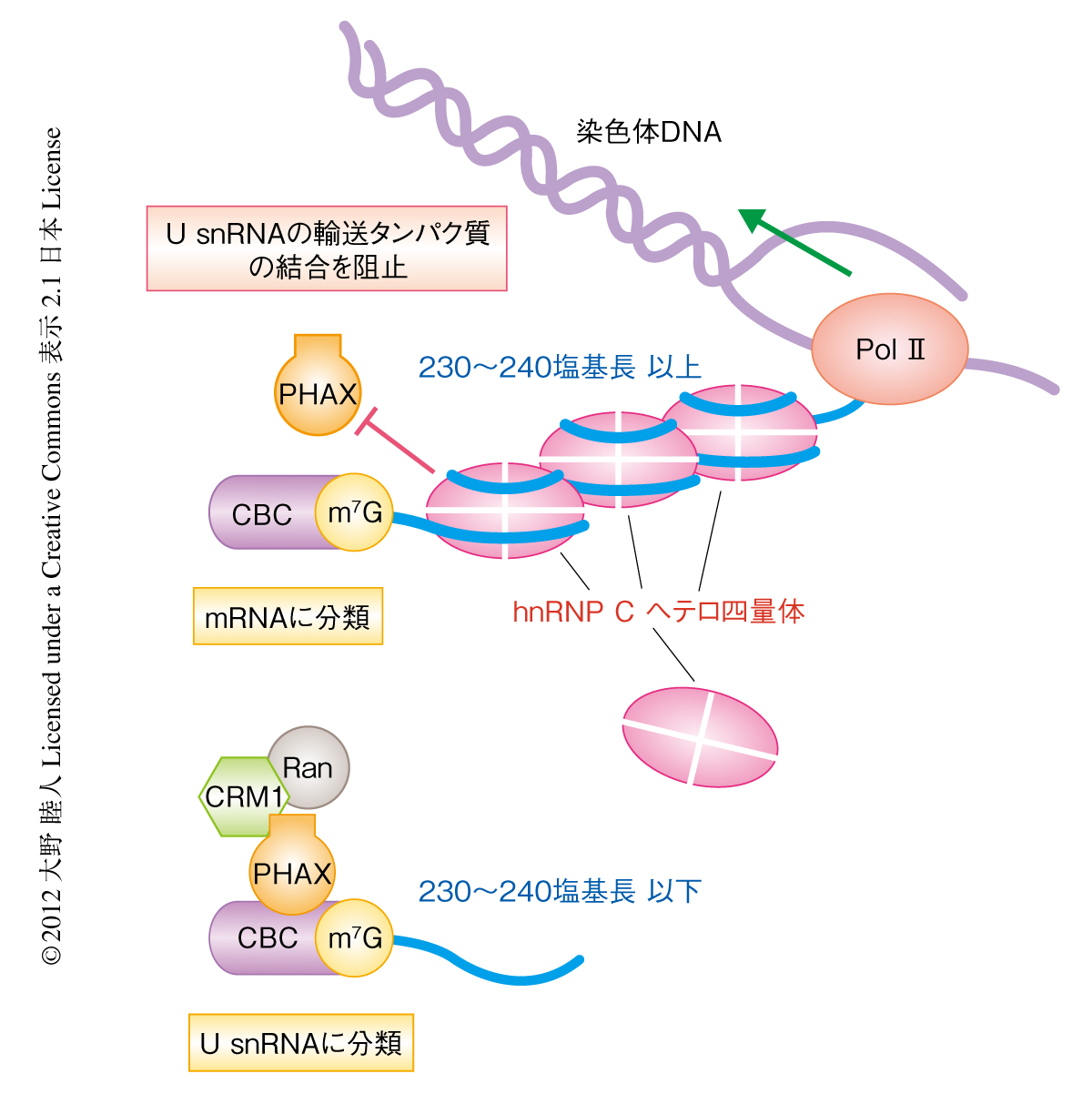
これらの結果にもとづき,細胞がRNAを長さに応じ分類する分子機構について,つぎのようなモデルを提唱した(図2).RNAポリメラーゼIIによる転写開始の直後,染色体DNAから新生RNAの5’末端が現われはじめると,そこにキャップ構造が付加されさらにキャップ構造結合タンパク質複合体が結合する.この短いRNAが合成された時点ではこのRNAが将来どの程度まで長くなるのか,つまり,mRNAになるのかU snRNAになるのかはわからない.転写がさらに進み新生RNAの長さが200~300塩基長より長くなると,hnRNP Cヘテロ四量体が少なくともひとつ安定に結合できるようになり,そのような転写産物はmRNA前駆体であると分類され同時にU snRNAの輸送タンパク質であるPHAXの結合が阻害される.逆に,RNAの長さが200~300塩基長より短いまま転写が終了した場合,hnRNP Cヘテロ四量体は安定に結合できず,そのような転写産物はU snRNA前駆体であると分類されPHAXをはじめとしたU snRNAの輸送タンパク質がRNAに呼び込まれる.
それでは,mRNAと分類されたRNAにmRNAの輸送タンパク質はどのように呼び込まれるのだろうか.mRNAの核外輸送におけるRNA結合アダプタータンパク質のひとつAly/REFは,PHAXやhnRNP Cヘテロ四量体と同じように,キャップ構造結合タンパク質複合体とその近傍のRNAの両方に結合するものと考えられている7,8).であるから,Aly/REFがmRNAに結合するためには,少なくともキャップ構造の近傍に結合するhnRNP Cヘテロ四量体と置き換わらなければならない.hnRNP Cヘテロ四量体はmRNAといっしょに核から細胞質へと輸送されることはないと考えられているので9),核あるいは核膜孔においてなんらかの機構によりmRNAから解離しmRNAの輸送タンパク質と置き換わるのであろう.
筆者らは,RNAポリメラーゼIIの転写産物が長さに応じ仕分けされる分子機構を明らかにした.しかし,RNAポリメラーゼIIの転写産物はmRNAとU snRNAだけではない.マイクロRNAの一次転写産物もRNAポリメラーゼIIにより合成される.マイクロRNAの一次転写産物は核においてマイクロプロセッサー複合体により約70塩基長のヘアピン型マイクロRNA前駆体へと切断されてから核外輸送されるが,この切断によりキャップ構造を失う.このマイクロプロセッサー複合体の呼び込みを介助するタンパク質としてArs2が同定されているが,Ars2もキャップ構造結合タンパク質複合体とRNAの両方に結合する10).このArs2がRNAポリメラーゼIIの転写産物がマイクロRNAへと仕分けされる際に活躍しているのであろう.また,U3やU8などの核小体内低分子RNA(small nucleolar RNA:snoRNA)もRNAポリメラーゼIIにより合成され,キャップ構造結合タンパク質複合体やPHAXが結合するところまでU snRNAと類似している.しかし,これらのRNAは核外輸送に対し優性にはたらく核小体局在シグナルをもっているため,細胞質へと輸送されることなく核小体へと輸送される11).
略歴:1989年 京都大学大学院理学研究科にて博士号取得,米国留学,京都大学理学部 助手,ドイツEuropean Molecular Biology Laboratory客員研究員などを経て,2001年より京都大学ウイルス研究所 教授.
研究テーマ:RNAと遺伝子発現の制御機構.RNAはDNAから転写されるやいなやタンパク質が結合しリボ核タンパク質(RNP)として誕生する.RNA/RNPは改変されてからそのはたらく場所まで輸送され機能する.RNA/RNPは古くなったり機能がなくなったりすると壊され死んでゆく.このような“RNA/RNPの一生”のさまざまな局面に興味をもっている.
関心事:やる気のある大学院生を常時募集中.研究室見学大歓迎.
研究室URL:http://www.virus.kyoto-u.ac.jp/Lab/ohnolab.html
© 2012 大野 睦人 Licensed under CC 表示 2.1 日本
(京都大学ウイルス研究所 情報高分子化学研究分野)
email:大野睦人
DOI: 10.7875/first.author.2012.046
HnRNP C tetramer measures RNA length to classify RNA polymerase II transcripts for export.
Asako McCloskey, Ichiro Taniguchi, Kaori Shinmyozu, Mutsuhito Ohno
Science, 335, 1643-1646 (2012)
要 約
通常,RNAの特異的な認識はその特異的な塩基配列や構造を介し行われる.しかし,筆者らは,核において“RNAの長さ”が認識される分子機構を発見した.RNAポリメラーゼIIはmRNAを合成するが,ほかにもU snRNAなど短いRNAも合成する.しかし,転写開始の直後にはその転写産物が将来mRNAになるのかU snRNAになるのかはわからない.転写がさらに進み新生RNAの長さが200~300塩基長より長くなると,hnRNP Cという核内RNA結合タンパク質のヘテロ四量体が安定に結合できるようになることで,U snRNAの輸送タンパク質の結合が阻害されその転写産物はmRNA前駆体であると認定される.逆に,RNAの長さが200~300塩基長より短いまま転写が終了した場合,hnRNP Cヘテロ四量体が安定に結合できずその転写産物はU snRNA前駆体であると認定される.このように,hnRNP Cヘテロ四量体が“分子のものさし”となってRNAの長さを測り,RNAをその長さに応じて仕分けしていた.
はじめに
真核生物ではほとんどのRNAは核において合成されるが,そののちの行き先はRNA種ごとに決まっている.多くのRNAは核から細胞質へと輸送されるが,そのようなRNAの主要なものとしては,リボソームRNA,tRNA,スプライソソームに含まれるウリジンに富む核内低分子RNA(U small nuclear RNA:U snRNA),mRNA,マイクロRNAなどの前駆体があげられる1).これらのRNAは核においてそれぞれのRNA種に固有の輸送タンパク質が結合したのち細胞質へと輸送される.核においてそれぞれのRNAに結合する輸送タンパク質は核外輸送をつかさどるだけでなく,そののちのRNAの運命(局在化,翻訳,安定性など)をも規定することが明らかになってきた.このことは,それぞれのRNA種は核においてすでに輸送タンパク質により識別されていることを意味する.しかし,それぞれのRNA種のいかなる特徴が核において識別されているのか,その識別を行う因子は何か,という点に関しては未知な部分が多い.
1.U snRNAとmRNAとを区別する特徴のひとつは“RNAの長さ”である
多くのRNA種のなかでもU snRNAとmRNAはいずれもRNAポリメラーゼIIにより合成され,5’末端にm7G型のキャップ構造が共通に形成されて,そこにはキャップ構造結合タンパク質複合体が結合する.しかし,このあとの核外輸送複合体の形成は両者でまったく違っている1).U snRNAの場合は,PHAX(phosphorylated adaptor for RNA export)というアダプタータンパク質がキャップ構造結合タンパク質複合体とその近傍のRNAに結合することによりRNAに呼び込まれ,そのPHAXが今度は核外輸送シグナル(nuclear export signal:NES)の輸送タンパク質CRM1-RanGTPを呼び込みU snRNA核外輸送複合体が完成する2).これに対して,mRNAの場合は,Aly/REFなどのRNA結合性のアダプタータンパク質がしばしばTREX(transcription/export)複合体やエキソンジャンクション複合体といった巨大複合体の一部としてmRNAに結合し,それがmRNAの主要な輸送タンパク質であるTAP/NXF1をRNAに呼び込む.このように,U snRNAとmRNAはある意味で似た者どうしではあるが,輸送タンパク質がきちんと区別できるような特徴をもっているはずである.
筆者らは,そのような特徴を探索した結果,“RNAの長さ”という概念にたどりついた.m7G型のキャップ構造をもつRNAは塩基配列ではなくその長さに応じてU snRNAの機構あるいはmRNAの機構により核外輸送されることを発見した.すなわち,人工的に長くしたU snRNAはmRNAの輸送タンパク質により,逆に,人工的に短くしたmRNAはU snRNAの輸送タンパク質より核から細胞質へと輸送されるようになったのである3,4).また,その切り替わりの長さの境目は約200~300塩基長であった5)(図1).ただし,強固な高次構造をもつRNA領域には長さの効果が表われないこともわかった3-5).このことは,細胞にはRNAの長さを測り輸送経路を決定する分子機構の存在することを意味していたが,その詳細については長いあいだ謎であった.
2.RNAの長さを測る分子機構
この問題に接近するため,RNAの長さに応じたRNA結合タンパク質の変化を試験管内で再現することを試みた.試験管内で合成したさまざまな長さのRNAと大腸菌により産生させ精製した組換えキャップ構造結合タンパク質複合体と組換えGST-PHAX融合タンパク質とを混合し,GSTのプルダウンを行うことによりU snRNA核外輸送複合体を部分的に模擬するRNA-キャップ構造結合タンパク質複合体-PHAX三者複合体の形成を調べた.その結果,この三者複合体はRNAの長さにかかわらずm7G型のキャップ構造をもつすべてのRNAにおいて形成された.この結果は,この精製系ではin vivoにおいてみられたRNAの長さに応じたRNA核外輸送複合体の形成がみられないことを意味した.しかし,HeLa細胞の核抽出液をこの精製系にくわえると,in vivoと同様に,長いRNAへのPHAXの結合が特異的に阻害された.このことは,核にはRNAの長さを測り長いRNAへのPHAXの結合を阻害する因子の存在することを示唆していた.
このPHAX呼び込み阻害活性をHeLa細胞の核抽出液からさまざまなカラムクロマトグラフィーを用いて精製した.最終精製画分にはこの活性と完全に相関する2本のメジャーなバンドがみられた.質量分析の結果,これらのバンドはヘテロ核内リボタンパク質(heterogeneous nuclear ribonucleoprotein:hnRNP)のC1とC2であることがわかった.hnRNP C1とhnRNP C2は3対1の割合でヘテロ四量体を形成する非常に量の多い核内RNA結合タンパク質であり,転写されつつあるmRNA前駆体につぎつぎと連続して結合しているものと考えられている6).
大腸菌で産生したhnRNP Cヘテロ四量体は試験管内において200~300塩基長以上の長さのRNAと特異的に結合しPHAXの結合を阻害した.これは,hnRNP Cヘテロ四量体がキャップ構造結合タンパク質複合体とRNAの両方に結合することによりPHAXの結合を競合的に阻害するためであることもわかった.RNAi法によりHeLa細胞においてhnRNP C1およびhnRNP C2をノックダウンすると,本来はmRNAに結合しないはずのPHAXがmRNAにも結合することが観察され,同時に,mRNAの核から細胞質への輸送が阻害された.これはおそらく,PHAXが結合したmRNAが異常なRNAであると認識され核外輸送が停止させられたのであろう.この考えと合致するように,hnRNP C1およびhnRNP C2とともにPHAXをノックダウンするとmRNAの核から細胞質への輸送が回復した.これらの結果から,細胞においてもこのhnRNP Cヘテロ四量体がPHAXとmRNAとの結合を防いでおり,そうすることでmRNAに適正な核外輸送複合体が形成されmRNAの核外輸送がとどこおりなく行われるよう管理していることが示された.
3.RNAの仕分けのモデル
これらの結果にもとづき,細胞がRNAを長さに応じ分類する分子機構について,つぎのようなモデルを提唱した(図2).RNAポリメラーゼIIによる転写開始の直後,染色体DNAから新生RNAの5’末端が現われはじめると,そこにキャップ構造が付加されさらにキャップ構造結合タンパク質複合体が結合する.この短いRNAが合成された時点ではこのRNAが将来どの程度まで長くなるのか,つまり,mRNAになるのかU snRNAになるのかはわからない.転写がさらに進み新生RNAの長さが200~300塩基長より長くなると,hnRNP Cヘテロ四量体が少なくともひとつ安定に結合できるようになり,そのような転写産物はmRNA前駆体であると分類され同時にU snRNAの輸送タンパク質であるPHAXの結合が阻害される.逆に,RNAの長さが200~300塩基長より短いまま転写が終了した場合,hnRNP Cヘテロ四量体は安定に結合できず,そのような転写産物はU snRNA前駆体であると分類されPHAXをはじめとしたU snRNAの輸送タンパク質がRNAに呼び込まれる.
それでは,mRNAと分類されたRNAにmRNAの輸送タンパク質はどのように呼び込まれるのだろうか.mRNAの核外輸送におけるRNA結合アダプタータンパク質のひとつAly/REFは,PHAXやhnRNP Cヘテロ四量体と同じように,キャップ構造結合タンパク質複合体とその近傍のRNAの両方に結合するものと考えられている7,8).であるから,Aly/REFがmRNAに結合するためには,少なくともキャップ構造の近傍に結合するhnRNP Cヘテロ四量体と置き換わらなければならない.hnRNP Cヘテロ四量体はmRNAといっしょに核から細胞質へと輸送されることはないと考えられているので9),核あるいは核膜孔においてなんらかの機構によりmRNAから解離しmRNAの輸送タンパク質と置き換わるのであろう.
おわりに
筆者らは,RNAポリメラーゼIIの転写産物が長さに応じ仕分けされる分子機構を明らかにした.しかし,RNAポリメラーゼIIの転写産物はmRNAとU snRNAだけではない.マイクロRNAの一次転写産物もRNAポリメラーゼIIにより合成される.マイクロRNAの一次転写産物は核においてマイクロプロセッサー複合体により約70塩基長のヘアピン型マイクロRNA前駆体へと切断されてから核外輸送されるが,この切断によりキャップ構造を失う.このマイクロプロセッサー複合体の呼び込みを介助するタンパク質としてArs2が同定されているが,Ars2もキャップ構造結合タンパク質複合体とRNAの両方に結合する10).このArs2がRNAポリメラーゼIIの転写産物がマイクロRNAへと仕分けされる際に活躍しているのであろう.また,U3やU8などの核小体内低分子RNA(small nucleolar RNA:snoRNA)もRNAポリメラーゼIIにより合成され,キャップ構造結合タンパク質複合体やPHAXが結合するところまでU snRNAと類似している.しかし,これらのRNAは核外輸送に対し優性にはたらく核小体局在シグナルをもっているため,細胞質へと輸送されることなく核小体へと輸送される11).
文 献
- Kohler, A. & Hurt, E.: Exporting RNA from the nucleus to the cytoplasm. Nat. Rev. Mol. Cell Biol., 8, 761-773 (2007)[PubMed]
- Ohno, M., Segref, A., Bachi, A. et al.: PHAX, a mediator of U snRNA nuclear export whose activity is regulated by phosphorylation. Cell, 101, 187-198 (2000)[PubMed]
- Ohno, M., Segref, A., Kuersten, S. et al.: Identity elements used in export of mRNAs. Mol. Cell, 9, 659-671 (2002)[PubMed]
- Fuke, H. & Ohno, M.: Role of poly (A) tail as an identity element for mRNA nuclear export. Nucleic Acids Res., 36, 1037-1049 (2008)[PubMed]
- Masuyama, M., Taniguchi, I., Kataoka, N. et al.: RNA length defines RNA export pathway. Genes Dev., 18, 2074-2085 (2004)[PubMed]
- Huang, M., Rech, J. E., Northington, S. J. et al.: The C-protein tetramer binds 230 to 240 nucleotides of pre-mRNA and nucleates the assembly of 40S heterogeneous nuclear ribonucleoprotein particles. Mol. Cell. Biol., 14, 518-533 (1994)[PubMed]
- Cheng, H., Dufu, K., Lee, C. S. et al.: Human mRNA export machinery recruited to the 5' end of mRNA. Cell, 127, 1389-1400 (2006)[PubMed]
- Nojima, T., Hirose, T., Kimura, H. et al.: The interaction between cap-binding complex and RNA export factor is required for intronless mRNA export. J. Biol. Chem., 282, 15645-15651 (2007)[PubMed]
- Nakielny, S. & Dreyfuss, G.: The hnRNP C proteins contain a nuclear retention sequence that can override nuclear export signals. J. Cell Biol., 134, 1365-1373 (1996)[PubMed]
- Gruber, J. J., Zatechka, D. S., Sabin, L. R. et al.: Ars2 links the nuclear cap-binding complex to RNA interference and cell proliferation. Cell, 138, 328-339 (2009)[PubMed]
- Kiss, T., Fayet, E., Jady, B. E. et al.: Biogenesis and intranuclear trafficking of human box C/D and H/ACA RNPs. Cold Spring Harb. Symp. Quant. Biol., 71, 407-417 (2006)[PubMed]
著者プロフィール
略歴:1989年 京都大学大学院理学研究科にて博士号取得,米国留学,京都大学理学部 助手,ドイツEuropean Molecular Biology Laboratory客員研究員などを経て,2001年より京都大学ウイルス研究所 教授.
研究テーマ:RNAと遺伝子発現の制御機構.RNAはDNAから転写されるやいなやタンパク質が結合しリボ核タンパク質(RNP)として誕生する.RNA/RNPは改変されてからそのはたらく場所まで輸送され機能する.RNA/RNPは古くなったり機能がなくなったりすると壊され死んでゆく.このような“RNA/RNPの一生”のさまざまな局面に興味をもっている.
関心事:やる気のある大学院生を常時募集中.研究室見学大歓迎.
研究室URL:http://www.virus.kyoto-u.ac.jp/Lab/ohnolab.html
© 2012 大野 睦人 Licensed under CC 表示 2.1 日本
