生体組織のマテリアル解析から形態制御を探る
中山 尚美
(スイスBern大学Institute of Plant Sciences)
email:中山尚美
DOI: 10.7875/first.author.2012.041
Elastic domains regulate growth and organogenesis in the plant shoot apical meristem.
Daniel Kierzkowski, Naomi Nakayama, Anne-Lise Routier-Kierzkowska, Alain Weber, Emmanuelle Bayer, Martine Schorderet, Didier Reinhardt, Cris Kuhlemeier, Richard S. Smith
Science, 335, 1096-1099 (2012)
生体組織の力学的な性質はその組織の形態変化の可能性を大きく左右する.植物の茎頂に存在する成長点であるメリステムでは,成長の遅く形態の安定した中心部と,成長の速く器官形成の起こる周縁部とが隣接している.この形態変化の差異に力学的な根拠があるかどうかを調べるため,生きた成長中の茎頂組織においてマテリアル解析を行った.浸透圧の原理により組織の内圧を定量的に変え,表面張力を操作することで生じる組織の表面のひずみを観測した.メリステムの部位的な伸長や縮小のしやすさを新しく開発した三次元イメージ解析ソフトウェアを用いて測定し,マテリアルとしての性質を有限要素法によりシミュレーションした.その結果,成長速度などメリステムの部位のあいだの形態変化の差異は,マテリアルの性質のささいな違いにより説明できることを見い出した.
ある素材でなんらかのかたちをつくろうとしたとき,その素材の力学的な性質により,つくりやすいかたちとつくりにくいかたちがある.同じように,生体組織の素材(マテリアル)としての性質は生物の形態形成に多大な影響を及ぼす.そのため,成長中の組織では主要な構造成分の性質が絶妙な精確さをもって時空的に制御されているものと考えられている.その逆もしかりで,マテリアルそのものがかたちのあり方を決めている,つまり,形態を直接的に制御していることもありうる.
そのような重要な役割にもかかわらず,発生の過程にある組織の力学的な性質はあまり分析されていない.とくに,形態変化の活発な組織では凹凸のある複雑な三次元構造体がダイナミックに変形するなかでの解析となり技術的にもむずかしくなる.しかし,近年,生物学と物理学,工学,数学,コンピューターサイエンスなどとの学際的な融合が進むにつれ,発生にかぎらずさまざまな生命現象の力学的な研究がさかんになり,つぎつぎと新しい技術や知見が生み出されている.これにならい,筆者らは,発生段階にある植物組織のマテリアルとしての解析をin situにおいて行う新しい手法を開発した.
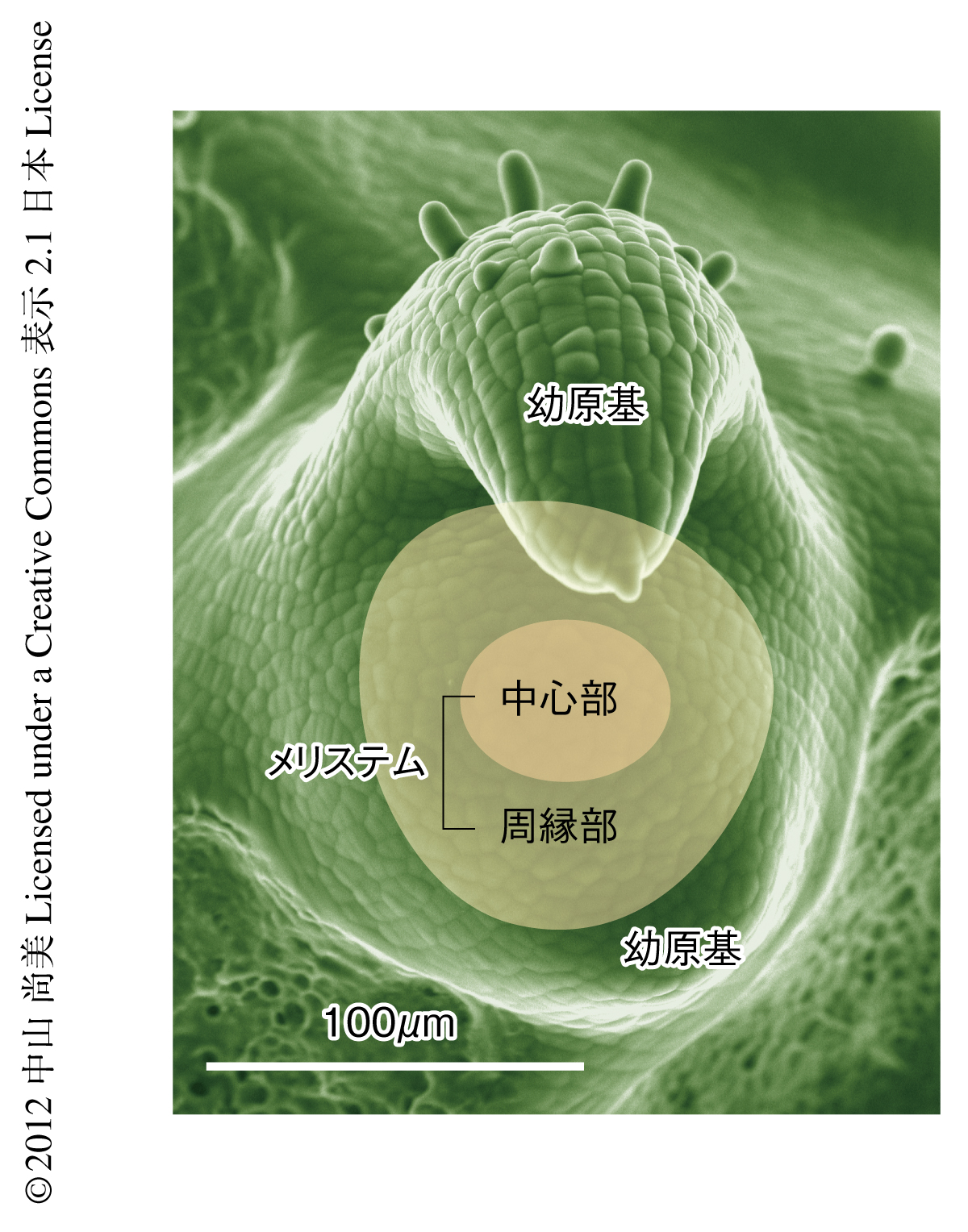
解析には植物の茎頂を使った(図1).古くは19世紀から,その形態形成が力学的に制御されていると考えられてきた,力学的な発生制御において古典的な系である1).植物は動物と異なり,胚発生をすぎても根や葉などの新しい器官をつくりつづける.茎頂には新しい葉や茎が現われる成長点であるメリステムがある.メリステムで新しく生まれた葉の原基は急速に成長しかたちを変えていく.はじめはただの球状の突起だった幼原基も,1日ほどで細長いシリンダー状に育ち,のちには個々の種や発生段階で決められた形態形成プログラムをたどる.
茎頂の形態形成に関しては力学的な解析がさかんに行われており,力学は器官形成や成長における重要な制御因子であることが明らかになってきた.メリステムや幼原基の細胞は組織にかかる力の方向に応じて成長する方向を調整するが,この力学的なシグナルへの反応なしでは幼原基は球状のまま成長し本来のかたちをつくることができない2).器官形成や成長を誘発するホルモンであるオーキシンの輸送もまた,メリステムでは力学的なシグナルに反応する3).オーキシンの下流で器官形成において必要なのが細胞壁におけるペクチンの変容で,新しい原基の内部の細胞壁を柔らかくするという力学的な変化に関与しているといわれている4,5).
被子植物の茎頂に存在するメリステムは2つの部位に分けることができる.成長の遅く細胞分裂が少ない中心部と,成長の速く活発に細胞分裂を行っている周縁部である6).周縁部は新しい葉や茎が生まれ分化をはじめる場所である.たとえば,トマトのメリステムでは周縁部は中心部に比べ倍以上の速度で成長している.中心部は幹細胞を含み,未分化の状態にとどまりつつ器官形成のもととなる細胞を周囲に送りつづける.形態変化が活発な組織の真ん中で変化を抑えて未分化な状態を保つことが,メリステムが正しく機能し新しい器官を供給していくために必要なのである.
これまで,遺伝学的な手法によりメリステムの2つの部位をつかさどる分子機構が解明されてきたが,それらは細胞膜にあるシグナル受容体や転写因子など分子経路を大きくゆるがす上流のスイッチ的な因子が主で,形態変化の違いを直接に制御する下流の因子はまだわかっていない6).そこで,メリステムのマテリアルとしての性質を明らかにすることにより,部位に特有の形態変化を力学的な視点からボトムアップ的に理解しようと試みた.
あるマテリアルの力学的な性質を示すのによく使われるのが,応力-ひずみ曲線である.これはその素材に,どれだけの応力(すなわち,単位面積にかかっている力)により,どれだけのひずみ(または,変形)が起こるかを示したグラフである.金属でできた“ばね”などほとんど直線的な関係を示す素材もあるが,ゴムのように力をくわえていくとはじめは直線的に変形するが,ある一定の値をこえると変形しにくくなるといった素材も多い.そのような非直線的な応力とひずみとの関係は,バイオマテリアルなどの複数の素材要素により構成される複合体によくみられる.
植物の細胞のかたちは,硬い外殻である細胞壁のかたちで決まる.そして,植物の組織のかたちは,だいたいにして細胞壁の連続体というマテリアルにより決められているといえるだろう.細胞壁はセルロースやヘミセルロースなど多糖性のポリマーの網目の合間をゲル状のペクチンが埋めることでできている7).単離された植物の細胞や組織の切片に力をかけると,ひずみは非直線的に生じることが多い8).メリステムも非直線的な応力-ひずみ曲線を示すものと推測できる.それでは,同じ非直線でも,メリステムの中心部と周縁部とでは応力とひずみとの関係に違いはあるのだろうか? それを調べるには,メリステムの表面にかかっている応力を定量的に変え,それぞれの部位において生じるひずみを測定する必要がある.
茎頂のメリステムは何百もの細胞からなる半球状の組織だが,ここで重要なのは表面の細胞壁の連続体に起こるひずみであり,内部の細胞壁は考慮しなくてもよい.組織の内部の細胞は互いに押し合っていて,相互にはたらいている力は打ち消されているからである.内部の細胞の発する力は組織の内圧として表皮を圧迫し,それをもちこたえる組織の表面の細胞壁は張りつめて構造強度を増す.透過型電子顕微鏡で観察すると,組織の表面の細胞壁は内部の細胞の細胞壁に比べ約7倍の厚さがあった.表面の細胞壁の連続体がメリステムの主要な力学的な構造であり,組織の全体の形態変化を大きく左右しているといえるだろう.メリステムにかぎらず,動物の胚など未分化で骨など強固な内部の支柱構造を含まない組織は,風船のように抽象化し表面層を中心として力学的に解析することができる.
これまでにも,植物の組織の力学的な解析は行われてきた.もっとも親しまれている方法は,暗所で長く育った胚軸を切り取り,片側からおもりをつけて引っ張るというものである7)(図2a).この方法は,長年にわたり植物の成長を促す因子の証明などに使われ,さまざまな発見を導いてきたが,ほかの組織への応用はむずかしい.近年,いくつかの研究室で開発されているのが,nmまたはμmのオーダーでの押し込みにより組織の圧縮されやすさを測定するデバイスである9)(図2b).2011年には,原子間力顕微鏡(atomic force microscope:AFM)を使った押し込みによるメリステムの表面のマテリアル解析も発表されている5,10).ただし,この押し込みは表面に対し垂直に行わなくてはならないため,メリステムのような立体的な組織を使用する場合には誤測定が懸念される.さらに,細胞壁など複雑なバイオマテリアルは均質でないことが多く,一方向に圧縮しやすいからといってほかの方向にも変形しやすいとはかぎらない.
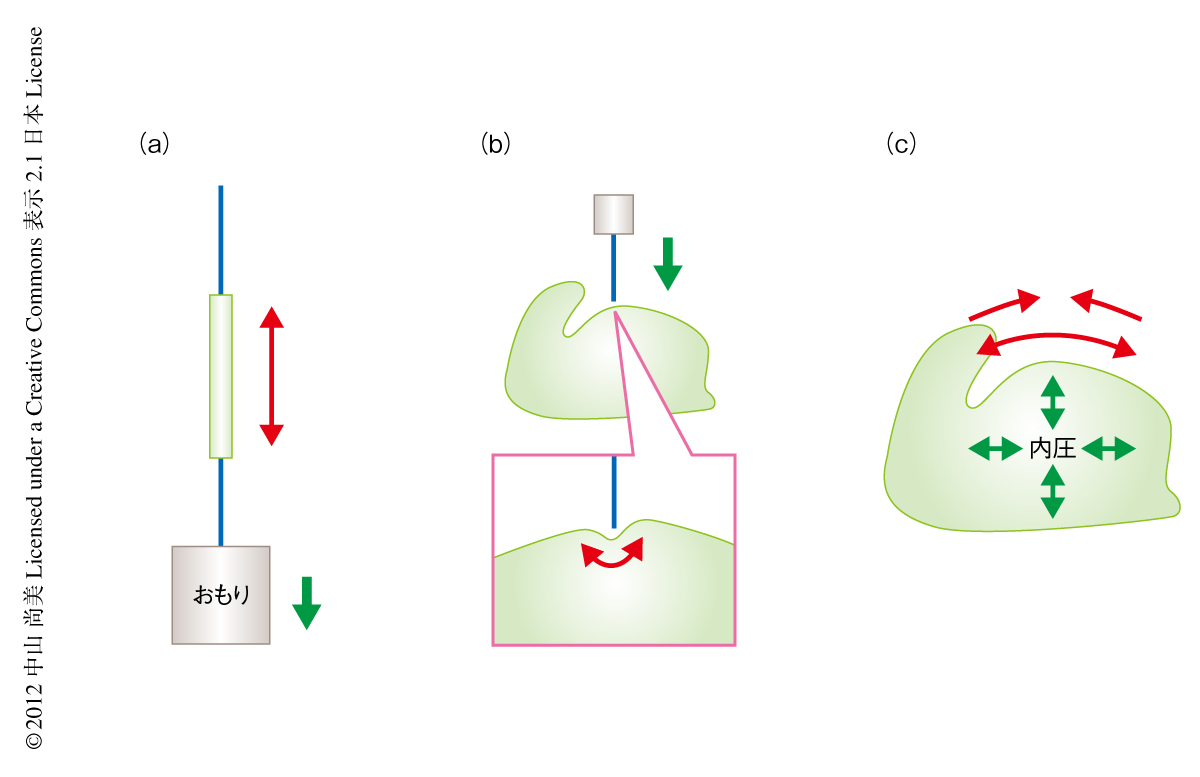
成長や形態形成にもっとも影響を及ぼすのは,組織の表面の細胞壁における表面と平行方向への伸長しやすさ,あるいは,しにくさである.そこで,組織の表面の張力を定量的に変えてどのくらいの伸長や縮小が促されるかをメリステムの全体で測定することにした(図2c).組織の表面の張力は組織の内圧から生じているから,この内圧を定量的に変化させればよい.浸透圧の原理を使えば,細胞を傷めることなく自然に組織の内圧を変えることができる.そこでまず,0.2 Osmolの水溶液中ならメリステムの本来の内圧やかたちを保てることを定めた.また,0.4 Osmolの水溶液中だとメリステムの細胞はすべて原形質分離したので組織の内圧は0になった.この2つの点から,メリステムを0 Osmolまたは0.4 Osmolの水溶液にさらすと,内圧が通常値の約5気圧から上下に5気圧変わると想定できた.
内圧の変化や組織の膨張あるいは縮小にともなう表面のひずみを測定するため,メリステムの表面につけた印の位置を浸透圧の変化の前後で比較した.細胞壁を蛍光染色することにより,組織の表面層のかたちの変化を細胞レベルの解像度でおうことのできる.共焦点レーザー顕微鏡を三次元スキャナーとして使用し,収集した画像を精確に空間解析するため,グラフィック機能を重視したソフトウェアMorphoGraphXを開発した.このMorphoGraphXの強みは,三次元のデータの外形を取り出して凹凸のある表面を二次元の平面のように扱えることである.ある一定の深さ(たとえば,表皮細胞の中間地点)からのデータを表面に投影し,平面のデータのようにシグナル解析を行うこともできる.
MorphoGraphXを使いメリステムの表皮細胞の表面の面積が5気圧の内圧の変化に対しどう反応するかをみた.すると,メリステムの中心部は縮みやすいが伸びにくいのに対し,周縁部は伸びやすいが縮みにくいことがわかった.内圧を通常値にもどすと表面の面積もほぼもとにもどった.このことから,5気圧の内圧の変化による組織の表面の細胞壁のひずみは弾性範囲にあるといえた.
膨張縮小試験の実験結果については,生化学反応も含めたさまざまで複雑な解釈が可能であろうが,実験データだけをもとにもっとも簡潔に説明しようと力学的なモデルに焦点をおいた.実験データからマテリアルの力学的な性質を推定するには工学的なモデリングを活用した.メリステムの表面を内圧のある外殻と見立て,有限要素法を用いて連続体力学シミュレーションを行った.
ひとつの組織においてみられるひずみの差異はマテリアルのかたちで説明できることがある.かたち自体が組織の内部の応力の差異を生むからである.内圧を含む外殻なら凸に曲がっている部位ほど応力は小さくなる.そこで,メリステムの表面の曲面性を測定した.中心部のほうが周縁部よりもより凸に曲がっていたので,周縁部の応力のほうが大きいことになった.メリステムの表面全体が均一の,徐々にひずみの生じにくくなる非直線的なマテリアルでできているとすると,周縁部のほうが縮小しやすいはずである.しかし実際は,周縁部のほうが中心部よりも縮小しにくかった.かたちだけでは2つの部位のひずみの違いを説明できそうにない.
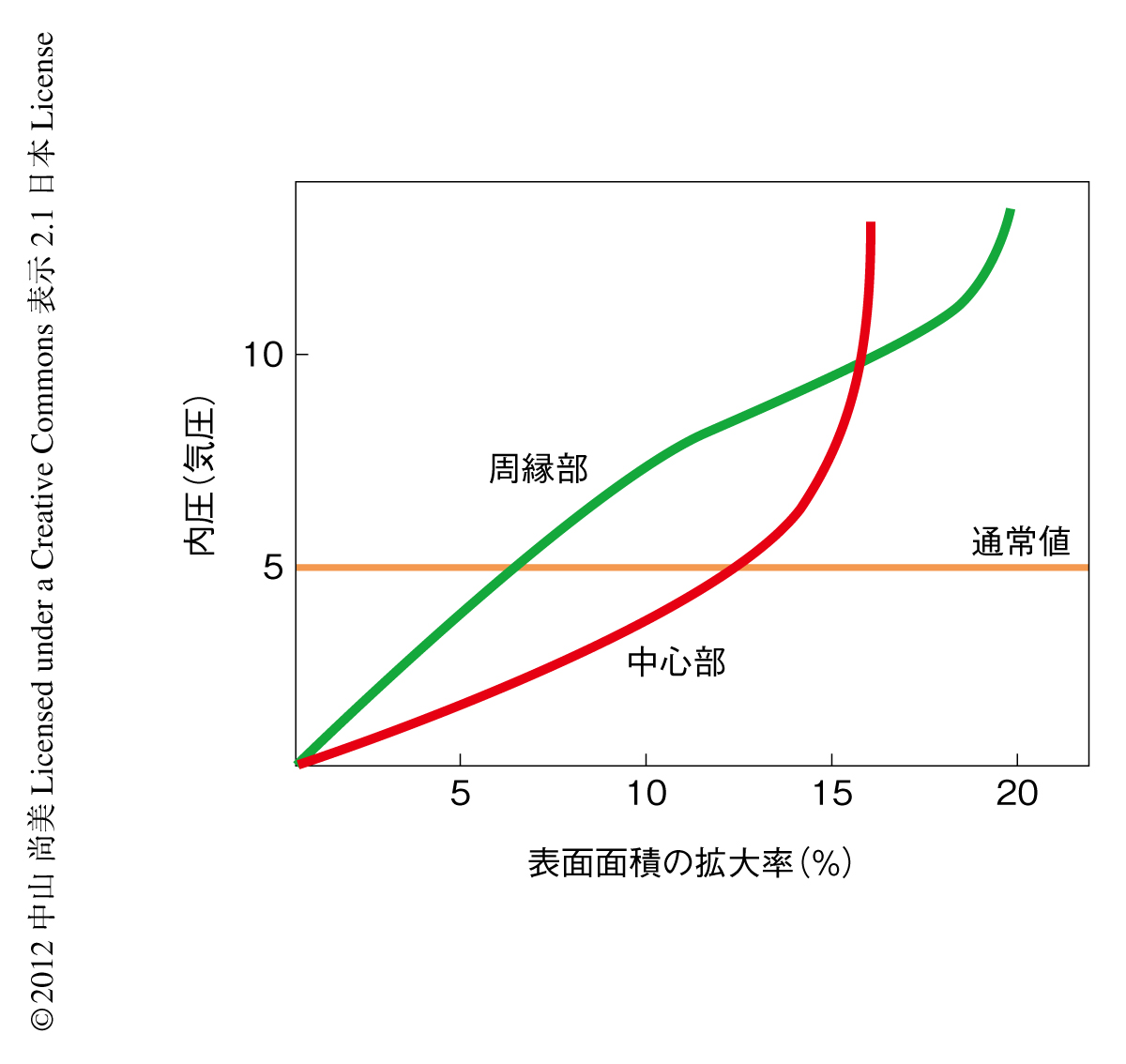
そこで,メリステムの中心部と周辺部では表面の細胞壁のマテリアルが異なると想定した.メリステムを均一な曲面の半球としてかたちによる応力の差をなくし,中心部と周縁部において膨張縮小試験で得られたのと同じひずみをシミュレーションできる,非直線的な応力-ひずみ曲線を選出した.その結果,部位による違いは,周縁部では中心部よりもマテリアルが“重たい”と仮定するだけで説明できることがわかった.ちょうど,ひとつに結ばれた細い輪ゴムと太い輪ゴムのように,同じ応力の変化をうけ異なるひずみを生じたのである.硬い周縁部は通常の内圧では応力-ひずみ曲線の直線的な部分に位置し,さらにまだ伸びる余地があった.その一方で,柔らかい素材でできた中心部は伸びきっていて,それ以上のひずみを生じにくい,いわゆる,ひずみ硬化とよばれる状態にあった(図3).
形態変化に乏しい中心部のほうが柔らかいマテリアルできているというのは意外に感じられるかもしれない.一般に,器官形成や成長などの形態変化は組織を柔らかくすることで活性化すると考えられている.生体組織の成長は不可逆性で,マテリアルがひずみ硬化をこえ弾性範囲を脱したときに起こる塑性ひずみに似ている.しかし,少なくともトマトの茎頂のメリステムでは,弾性範囲の伸長性を増加することで形態変化が活性化されていることを膨張縮小試験は示唆した.そうなると,弾性伸長がどのようにして細胞壁の内部の複合体の構造を不可逆的に変えるのかが今後の課題のひとつとなる.
メリステムのマテリアル解析により,組織の力学的な性質がメリステムにおける部位に特有の形態変化やメリステムの機能を直接的に制御している可能性がみえてきた.ひずみ硬化がメリステムの中心部において形態変化を抑え,未分化な幹細胞の居場所を守る.そのマテリアルが周囲に移動する途中で“重たく”なることにより,周縁部での形態形成が可能になる.どのような分子機構がこのマテリアルの性質の変化を起こしているのだろうか? そして,そのような下流の現象がメリステムの部位を特定するどの上流因子につながるのだろうか? メリステムの形態形成制御の鍵となる分子ネットワークの解明に力学的な解析が役立つことはまちがいないだろう.
発生の力学的な制御は,化学的な制御と対立するものではなく,互いにフィードバックして協力関係にあることが多い.生化学系は,環境に敏感なほど,そして,関連因子が多く複雑になるほど,ノイズや誤作動が起こりやすくなる.マテリアルの性質による力学的な制御は,そんな生化学系の不安定性が形態変化として現われるのをとめる防波堤のようなものなのかもしれない.それは,茎頂のメリステムにおける成長や分化の空間的な制御のような,まちがいの取り返しがつかない現象ではことさら大切であるものと思われる.
略歴:2006年 米国Yale大学にて博士号 取得,同年よりスイスBern大学 ポスドク研究者.
研究テーマ:植物の発生における非線形物理学的な制御.
抱負:植物のつくりや成り立ちに関する,おもしろい研究をしていきたい.
© 2012 中山 尚美 Licensed under CC 表示 2.1 日本
(スイスBern大学Institute of Plant Sciences)
email:中山尚美
DOI: 10.7875/first.author.2012.041
Elastic domains regulate growth and organogenesis in the plant shoot apical meristem.
Daniel Kierzkowski, Naomi Nakayama, Anne-Lise Routier-Kierzkowska, Alain Weber, Emmanuelle Bayer, Martine Schorderet, Didier Reinhardt, Cris Kuhlemeier, Richard S. Smith
Science, 335, 1096-1099 (2012)
要 約
生体組織の力学的な性質はその組織の形態変化の可能性を大きく左右する.植物の茎頂に存在する成長点であるメリステムでは,成長の遅く形態の安定した中心部と,成長の速く器官形成の起こる周縁部とが隣接している.この形態変化の差異に力学的な根拠があるかどうかを調べるため,生きた成長中の茎頂組織においてマテリアル解析を行った.浸透圧の原理により組織の内圧を定量的に変え,表面張力を操作することで生じる組織の表面のひずみを観測した.メリステムの部位的な伸長や縮小のしやすさを新しく開発した三次元イメージ解析ソフトウェアを用いて測定し,マテリアルとしての性質を有限要素法によりシミュレーションした.その結果,成長速度などメリステムの部位のあいだの形態変化の差異は,マテリアルの性質のささいな違いにより説明できることを見い出した.
はじめに
ある素材でなんらかのかたちをつくろうとしたとき,その素材の力学的な性質により,つくりやすいかたちとつくりにくいかたちがある.同じように,生体組織の素材(マテリアル)としての性質は生物の形態形成に多大な影響を及ぼす.そのため,成長中の組織では主要な構造成分の性質が絶妙な精確さをもって時空的に制御されているものと考えられている.その逆もしかりで,マテリアルそのものがかたちのあり方を決めている,つまり,形態を直接的に制御していることもありうる.
そのような重要な役割にもかかわらず,発生の過程にある組織の力学的な性質はあまり分析されていない.とくに,形態変化の活発な組織では凹凸のある複雑な三次元構造体がダイナミックに変形するなかでの解析となり技術的にもむずかしくなる.しかし,近年,生物学と物理学,工学,数学,コンピューターサイエンスなどとの学際的な融合が進むにつれ,発生にかぎらずさまざまな生命現象の力学的な研究がさかんになり,つぎつぎと新しい技術や知見が生み出されている.これにならい,筆者らは,発生段階にある植物組織のマテリアルとしての解析をin situにおいて行う新しい手法を開発した.
1.題材と問い
解析には植物の茎頂を使った(図1).古くは19世紀から,その形態形成が力学的に制御されていると考えられてきた,力学的な発生制御において古典的な系である1).植物は動物と異なり,胚発生をすぎても根や葉などの新しい器官をつくりつづける.茎頂には新しい葉や茎が現われる成長点であるメリステムがある.メリステムで新しく生まれた葉の原基は急速に成長しかたちを変えていく.はじめはただの球状の突起だった幼原基も,1日ほどで細長いシリンダー状に育ち,のちには個々の種や発生段階で決められた形態形成プログラムをたどる.
茎頂の形態形成に関しては力学的な解析がさかんに行われており,力学は器官形成や成長における重要な制御因子であることが明らかになってきた.メリステムや幼原基の細胞は組織にかかる力の方向に応じて成長する方向を調整するが,この力学的なシグナルへの反応なしでは幼原基は球状のまま成長し本来のかたちをつくることができない2).器官形成や成長を誘発するホルモンであるオーキシンの輸送もまた,メリステムでは力学的なシグナルに反応する3).オーキシンの下流で器官形成において必要なのが細胞壁におけるペクチンの変容で,新しい原基の内部の細胞壁を柔らかくするという力学的な変化に関与しているといわれている4,5).
被子植物の茎頂に存在するメリステムは2つの部位に分けることができる.成長の遅く細胞分裂が少ない中心部と,成長の速く活発に細胞分裂を行っている周縁部である6).周縁部は新しい葉や茎が生まれ分化をはじめる場所である.たとえば,トマトのメリステムでは周縁部は中心部に比べ倍以上の速度で成長している.中心部は幹細胞を含み,未分化の状態にとどまりつつ器官形成のもととなる細胞を周囲に送りつづける.形態変化が活発な組織の真ん中で変化を抑えて未分化な状態を保つことが,メリステムが正しく機能し新しい器官を供給していくために必要なのである.
これまで,遺伝学的な手法によりメリステムの2つの部位をつかさどる分子機構が解明されてきたが,それらは細胞膜にあるシグナル受容体や転写因子など分子経路を大きくゆるがす上流のスイッチ的な因子が主で,形態変化の違いを直接に制御する下流の因子はまだわかっていない6).そこで,メリステムのマテリアルとしての性質を明らかにすることにより,部位に特有の形態変化を力学的な視点からボトムアップ的に理解しようと試みた.
2.生体組織の力学的な解析
あるマテリアルの力学的な性質を示すのによく使われるのが,応力-ひずみ曲線である.これはその素材に,どれだけの応力(すなわち,単位面積にかかっている力)により,どれだけのひずみ(または,変形)が起こるかを示したグラフである.金属でできた“ばね”などほとんど直線的な関係を示す素材もあるが,ゴムのように力をくわえていくとはじめは直線的に変形するが,ある一定の値をこえると変形しにくくなるといった素材も多い.そのような非直線的な応力とひずみとの関係は,バイオマテリアルなどの複数の素材要素により構成される複合体によくみられる.
植物の細胞のかたちは,硬い外殻である細胞壁のかたちで決まる.そして,植物の組織のかたちは,だいたいにして細胞壁の連続体というマテリアルにより決められているといえるだろう.細胞壁はセルロースやヘミセルロースなど多糖性のポリマーの網目の合間をゲル状のペクチンが埋めることでできている7).単離された植物の細胞や組織の切片に力をかけると,ひずみは非直線的に生じることが多い8).メリステムも非直線的な応力-ひずみ曲線を示すものと推測できる.それでは,同じ非直線でも,メリステムの中心部と周縁部とでは応力とひずみとの関係に違いはあるのだろうか? それを調べるには,メリステムの表面にかかっている応力を定量的に変え,それぞれの部位において生じるひずみを測定する必要がある.
茎頂のメリステムは何百もの細胞からなる半球状の組織だが,ここで重要なのは表面の細胞壁の連続体に起こるひずみであり,内部の細胞壁は考慮しなくてもよい.組織の内部の細胞は互いに押し合っていて,相互にはたらいている力は打ち消されているからである.内部の細胞の発する力は組織の内圧として表皮を圧迫し,それをもちこたえる組織の表面の細胞壁は張りつめて構造強度を増す.透過型電子顕微鏡で観察すると,組織の表面の細胞壁は内部の細胞の細胞壁に比べ約7倍の厚さがあった.表面の細胞壁の連続体がメリステムの主要な力学的な構造であり,組織の全体の形態変化を大きく左右しているといえるだろう.メリステムにかぎらず,動物の胚など未分化で骨など強固な内部の支柱構造を含まない組織は,風船のように抽象化し表面層を中心として力学的に解析することができる.
これまでにも,植物の組織の力学的な解析は行われてきた.もっとも親しまれている方法は,暗所で長く育った胚軸を切り取り,片側からおもりをつけて引っ張るというものである7)(図2a).この方法は,長年にわたり植物の成長を促す因子の証明などに使われ,さまざまな発見を導いてきたが,ほかの組織への応用はむずかしい.近年,いくつかの研究室で開発されているのが,nmまたはμmのオーダーでの押し込みにより組織の圧縮されやすさを測定するデバイスである9)(図2b).2011年には,原子間力顕微鏡(atomic force microscope:AFM)を使った押し込みによるメリステムの表面のマテリアル解析も発表されている5,10).ただし,この押し込みは表面に対し垂直に行わなくてはならないため,メリステムのような立体的な組織を使用する場合には誤測定が懸念される.さらに,細胞壁など複雑なバイオマテリアルは均質でないことが多く,一方向に圧縮しやすいからといってほかの方向にも変形しやすいとはかぎらない.
成長や形態形成にもっとも影響を及ぼすのは,組織の表面の細胞壁における表面と平行方向への伸長しやすさ,あるいは,しにくさである.そこで,組織の表面の張力を定量的に変えてどのくらいの伸長や縮小が促されるかをメリステムの全体で測定することにした(図2c).組織の表面の張力は組織の内圧から生じているから,この内圧を定量的に変化させればよい.浸透圧の原理を使えば,細胞を傷めることなく自然に組織の内圧を変えることができる.そこでまず,0.2 Osmolの水溶液中ならメリステムの本来の内圧やかたちを保てることを定めた.また,0.4 Osmolの水溶液中だとメリステムの細胞はすべて原形質分離したので組織の内圧は0になった.この2つの点から,メリステムを0 Osmolまたは0.4 Osmolの水溶液にさらすと,内圧が通常値の約5気圧から上下に5気圧変わると想定できた.
内圧の変化や組織の膨張あるいは縮小にともなう表面のひずみを測定するため,メリステムの表面につけた印の位置を浸透圧の変化の前後で比較した.細胞壁を蛍光染色することにより,組織の表面層のかたちの変化を細胞レベルの解像度でおうことのできる.共焦点レーザー顕微鏡を三次元スキャナーとして使用し,収集した画像を精確に空間解析するため,グラフィック機能を重視したソフトウェアMorphoGraphXを開発した.このMorphoGraphXの強みは,三次元のデータの外形を取り出して凹凸のある表面を二次元の平面のように扱えることである.ある一定の深さ(たとえば,表皮細胞の中間地点)からのデータを表面に投影し,平面のデータのようにシグナル解析を行うこともできる.
MorphoGraphXを使いメリステムの表皮細胞の表面の面積が5気圧の内圧の変化に対しどう反応するかをみた.すると,メリステムの中心部は縮みやすいが伸びにくいのに対し,周縁部は伸びやすいが縮みにくいことがわかった.内圧を通常値にもどすと表面の面積もほぼもとにもどった.このことから,5気圧の内圧の変化による組織の表面の細胞壁のひずみは弾性範囲にあるといえた.
3.解釈と知見
膨張縮小試験の実験結果については,生化学反応も含めたさまざまで複雑な解釈が可能であろうが,実験データだけをもとにもっとも簡潔に説明しようと力学的なモデルに焦点をおいた.実験データからマテリアルの力学的な性質を推定するには工学的なモデリングを活用した.メリステムの表面を内圧のある外殻と見立て,有限要素法を用いて連続体力学シミュレーションを行った.
ひとつの組織においてみられるひずみの差異はマテリアルのかたちで説明できることがある.かたち自体が組織の内部の応力の差異を生むからである.内圧を含む外殻なら凸に曲がっている部位ほど応力は小さくなる.そこで,メリステムの表面の曲面性を測定した.中心部のほうが周縁部よりもより凸に曲がっていたので,周縁部の応力のほうが大きいことになった.メリステムの表面全体が均一の,徐々にひずみの生じにくくなる非直線的なマテリアルでできているとすると,周縁部のほうが縮小しやすいはずである.しかし実際は,周縁部のほうが中心部よりも縮小しにくかった.かたちだけでは2つの部位のひずみの違いを説明できそうにない.
そこで,メリステムの中心部と周辺部では表面の細胞壁のマテリアルが異なると想定した.メリステムを均一な曲面の半球としてかたちによる応力の差をなくし,中心部と周縁部において膨張縮小試験で得られたのと同じひずみをシミュレーションできる,非直線的な応力-ひずみ曲線を選出した.その結果,部位による違いは,周縁部では中心部よりもマテリアルが“重たい”と仮定するだけで説明できることがわかった.ちょうど,ひとつに結ばれた細い輪ゴムと太い輪ゴムのように,同じ応力の変化をうけ異なるひずみを生じたのである.硬い周縁部は通常の内圧では応力-ひずみ曲線の直線的な部分に位置し,さらにまだ伸びる余地があった.その一方で,柔らかい素材でできた中心部は伸びきっていて,それ以上のひずみを生じにくい,いわゆる,ひずみ硬化とよばれる状態にあった(図3).
形態変化に乏しい中心部のほうが柔らかいマテリアルできているというのは意外に感じられるかもしれない.一般に,器官形成や成長などの形態変化は組織を柔らかくすることで活性化すると考えられている.生体組織の成長は不可逆性で,マテリアルがひずみ硬化をこえ弾性範囲を脱したときに起こる塑性ひずみに似ている.しかし,少なくともトマトの茎頂のメリステムでは,弾性範囲の伸長性を増加することで形態変化が活性化されていることを膨張縮小試験は示唆した.そうなると,弾性伸長がどのようにして細胞壁の内部の複合体の構造を不可逆的に変えるのかが今後の課題のひとつとなる.
メリステムのマテリアル解析により,組織の力学的な性質がメリステムにおける部位に特有の形態変化やメリステムの機能を直接的に制御している可能性がみえてきた.ひずみ硬化がメリステムの中心部において形態変化を抑え,未分化な幹細胞の居場所を守る.そのマテリアルが周囲に移動する途中で“重たく”なることにより,周縁部での形態形成が可能になる.どのような分子機構がこのマテリアルの性質の変化を起こしているのだろうか? そして,そのような下流の現象がメリステムの部位を特定するどの上流因子につながるのだろうか? メリステムの形態形成制御の鍵となる分子ネットワークの解明に力学的な解析が役立つことはまちがいないだろう.
おわりに
発生の力学的な制御は,化学的な制御と対立するものではなく,互いにフィードバックして協力関係にあることが多い.生化学系は,環境に敏感なほど,そして,関連因子が多く複雑になるほど,ノイズや誤作動が起こりやすくなる.マテリアルの性質による力学的な制御は,そんな生化学系の不安定性が形態変化として現われるのをとめる防波堤のようなものなのかもしれない.それは,茎頂のメリステムにおける成長や分化の空間的な制御のような,まちがいの取り返しがつかない現象ではことさら大切であるものと思われる.
文 献
- Thompson, D. W.: On Growth and Form, 2nd ed. Cambridge University Press, Cambridge (1942)
- Hamant, O., Heisler, M. G., Jonsson, H. et al.: Developmental patterning by mechanical signals in Arabidopsis. Science, 322, 1650-1655 (2008)[PubMed]
- Heisler, M. G., Hamant, O., Krupinski, P. et al.: Alignment between PIN1 polarity and microtuble orientation in the shoot apical meristem reveals a tight coupling between morphogenesis and auxin transport. PLoS Biol., 8, e1000516 (2010)[PubMed]
- Peaucelle, A., Louvet, R., Johansen, J. N. et al.: Arabidopsis phyllotaxis is controlled by the methyl-esterification status of cell wall pectins. Curr. Biol., 18, 1943-1948 (2008)[PubMed]
- Peaucelle, A., Braybrook, S. A., Le Guillou, L. et al.: Pectin-induced changes in cell wall mechanics underlie organ initiation in Arabidopsis. Curr. Biol., 21, 1720-1726 (2011)[PubMed]
- Sablowski, R.: The dynamic plant cell niches. Curr. Opin. Plant Biol., 10, 639-644 (2007)[PubMed]
- Cosgrove, D. J.: Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol., 6, 850-861 (2005)[PubMed]
- Kerstens, S., Decraemer, W. F. & Verbelen, J. P.: Cell walls at the plant surface behave mechanically like fiber-reinforced composite materials. Plant Physiol., 127, 381-385 (2001)[PubMed]
- Geitmann, A.: Experimental approaches used to quantify physical parameters at cellular and subcellular levels. Am. J. Bot., 93, 1380-1390 (2006)[PubMed]
- Milani, P., Gholamirad, M., Traas, J. et al.: In vivo analysis of local wall stiffness at the shoot apical meristem in Arabidopsis using atomic force microscopy. Plant J., 67, 1116-1123 (2011)[PubMed]
著者プロフィール
略歴:2006年 米国Yale大学にて博士号 取得,同年よりスイスBern大学 ポスドク研究者.
研究テーマ:植物の発生における非線形物理学的な制御.
抱負:植物のつくりや成り立ちに関する,おもしろい研究をしていきたい.
© 2012 中山 尚美 Licensed under CC 表示 2.1 日本
