ヒトのムスカリン性アセチルコリン受容体M2サブタイプのX線結晶構造解析
芳賀達也1・Brian K. Kobilka 2・小林拓也3
(1学習院大学理学部 生命科学研究科,2米国Stanford大学School of Medicine,Department of Molecular and Cellular Physiology,3京都大学大学院医学研究科 分子細胞情報学)
email:小林拓也
DOI: 10.7875/first.author.2012.018
Structure of the human M2 muscarinic acetylcholine receptor bound to an antagonist.
Kazuko Haga, Andrew C. Kruse, Hidetsugu Asada, Takami Yurugi-Kobayashi, Mitsunori Shiroishi, Cheng Zhang, William I. Weis, Tetsuji Okada, Brian K. Kobilka, Tatsuya Haga, Takuya Kobayashi
Nature, 482, 547-551 (2012)
Gタンパク質共役受容体はヒトのゲノムにおいて非常に大きなファミリーを形成している.また,市販の医薬品の作用点の約40%をしめるともいわれており,創薬において重要なターゲットと位置づけられている.数多いGタンパク質共役受容体ファミリーのなかでも,ムスカリン受容体はオルソステリック部位とアロステリック部位とをもつといわれ,おのおのの部位を区別して結合する低分子化合物が報告されている.筆者らは,ヒトのムスカリン性アセチルコリン受容体M2サブタイプのオルソステリック部位にアンタゴニストが結合した複合体のX線結晶構造を解明した.ムスカリンM2受容体の立体構造が明らかになり,抗コリン薬の結合部位の詳細な情報を得ることができた.これらの立体構造の情報をもとに,今後,より効果的で副作用のないサブタイプに選択的な薬の探索および設計が可能になるものと考えられる.
アセチルコリンは世界で最初に神経伝達物質のひとつであることが証明された物質である.アセチルコリンにはニコチン作用とムスカリン作用とが存在することが古くから知られており,おのおのに特異的なアセチルコリン受容体(ニコチン受容体とムスカリン受容体)が同定されている.副交感神経や運動神経の末端から分泌されたアセチルコリンは骨格筋や内臓平滑筋などに存在するアセチルコリン受容体にはたらき,筋収縮を促進するほか副交感神経を刺激し,心拍数の減少,脈拍数の減少,唾液の分泌を促進する.このように,アセチルコリンは基本的な生命現象に深く関与している.とくに,脳内のアセチルコリンのバランスがくずれるとさまざまな疾患が生じる.たとえば,アセチルコリンが減少すると自律神経失調症やアルツハイマー病につながるといわれている.また,パーキンソン病の患者ではアセチルコリンの相対的な増加が認められている.アセチルコリンのムスカリン受容体への結合を遮断することでパーキンソン病の症状が緩和することが知られており,古くから治療薬として使われている.一方で,副作用として統合失調症や認知症の症状を悪化させることも知られている.
ムスカリン受容体にはM1からM5までの5種類のサブタイプが存在する1).たとえば,M1受容体は大脳皮質や海馬に多く存在し記憶や学習に関与していると考えられている.また,M2受容体はおもに中枢神経や心臓に分布しその機能を抑制的に制御している.このように,おのおののサブタイプは固有の分布と薬理作用をもっており,構造にもとづいたサブタイプに選択的な創薬が期待されている.
膜タンパク質は市販の医薬品の作用点の50%以上をしめるといわれており,創薬において重要なターゲットと位置づけられている.そのなかでもGタンパク質共役型受容体は創薬の標的として注目されている.“構造にもとづく創薬戦略”によりGタンパク質共役受容体の立体構造を体系的かつ網羅的に解明していくことは,候補化合物を迅速に探索する手法の開発,候補化合物の最適化,副作用の少ない化合物の分子設計につながる.しかし,Gタンパク質共役受容体の構造解析は非常にむずかしく,長いあいだ,その構造は解かれていなかった.その理由として,非常に疎水性領域が多いため(全体の70~80%)界面活性剤で可溶化するとそのほとんどがミセルでおおわれてしまい結晶格子が形成されにくいこと,そして,活性型と不活性型の構造をとりうる柔軟性の高い膜タンパク質であることが考えられていた.
筆者らは2000年から,ヒトをターゲットとしたGタンパク質共役受容体のX線結晶構造解析を目的として結晶化に適した試料の調製を試みてきた.その結果,ヒトGタンパク質共役受容体のうちの3つ,アデノシンA2a受容体,ヒスタミンH1受容体,ムスカリンM2受容体の結晶構造を高分解能で解明することに成功した.アデノシンA2a受容体についてはその立体構造を認識する抗体を作製して抗体との複合体の結晶構造を3.1Åで決定した2)(新着論文レビュー でも掲載).さらに,ヒスタミンH1受容体についてはリゾチームを融合したうえで脂質キュービック相に結晶化し高分解能(3.1Å)でその構造を解いた3).今回,ムスカリンM2受容体についても同様な方法を使い分解能3.0Åで結晶構造を決定した.ヒスタミンH1受容体およびアデノシンA2a受容体はメタノール資化酵母を用い発現させたのち精製したが,ムスカリンM2受容体は昆虫細胞を用いて調製した.よい結晶を生産するには,いずれの場合も培地1リットルあたり1 mg以上の生産量が必要となった.Gタンパク質共役受容体の高発現を実現するためには,柔軟性の高いGタンパク質共役受容体を安定化する必要があった.従来は,多くの変異体を地道に作製しそのリガンド結合活性を測定していたが,相同組換え活性の高い出芽酵母を用いC末端にGFPを融合させることにより短時間に安定性を向上させる系を確立した.現在,1カ月で50~100種類以上の変異体をハイスループットでスクリーニングしている.この技術により,まさに“解けるものを解く研究”から“解きたいものを解く研究”が可能になったといえる.
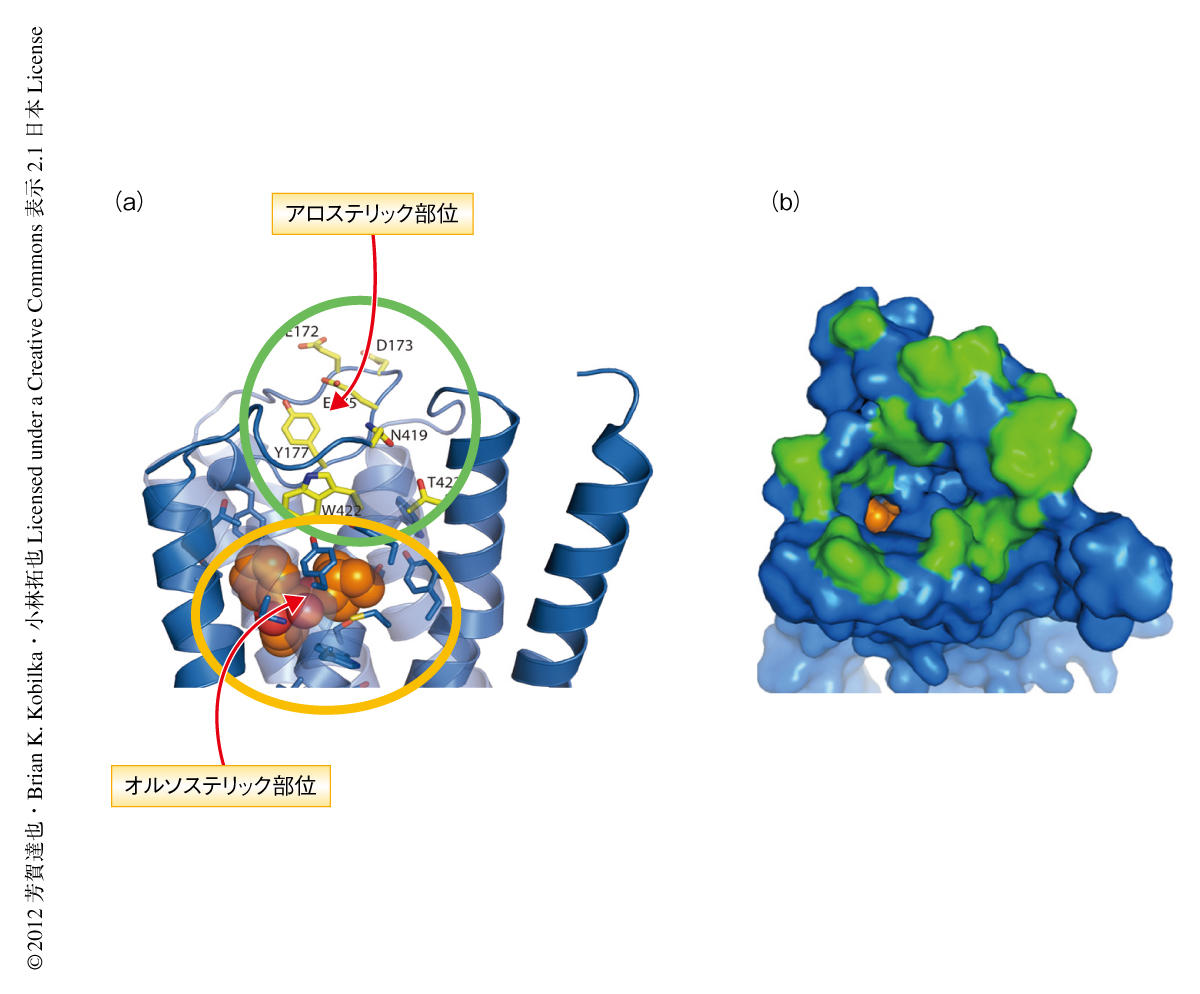
Gタンパク質共役受容体ファミリーのなかでも,ムスカリン性アセチルコリン受容体は2つのリガンド結合部位(オルソステリック部位およびアロステリック部位)をかねそなえた特徴的な受容体である.Gタンパク質共役受容体のアロステリック制御についてはじめて報告されたのは比較的最近であり4),アロステリック部位の立体構造はこれまで明らかにされていなかった.筆者らは,今回,ヒトのムスカリン受容体M2サブタイプのオルソステリック部位にアンタゴニストが結合した複合体の結晶構造を世界ではじめて高分解能(3.0Å)で解くことに成功した(PDB ID:3UON).その結果,神経伝達物質であるアセチルコリンが結合するオルソステリック部位を構成する19個のアミノ酸残基がアンタゴニストであるキヌクリジニルベンジレートと作用しており(図1),そのうち18個のアミノ酸残基はすべてのムスカリン受容体サブタイプ(M1~M5)において保存されていた.このことより,ムスカリン受容体においてサブタイプに選択的に作用するリガンドの開発が非常にむずかしかったことを原子分解能レベルではじめて証明することができた.一方,アロステリック部位はオルソステリック部位の上部にある比較的広い空洞として細胞の外側に広がっていた(図2).アロステリック部位を構成するいくつかのアミノ酸残基は,アミノ酸変異実験によりアロステリックリガンドの結合親和性を低下させるアミノ酸残基と一致していた.アロステリック部位にはそれぞれのサブタイプにおいて特徴的なアミノ酸残基が存在していた.したがって,それぞれのサブタイプに特徴的なアミノ酸残基の側鎖とリガンドの官能基とのあいだに相互作用(水素結合など)を生み出すことができ,これはサブタイプに選択的なリガンドの開発につながると考えられた.

筆者らはこれまでに,Gタンパク質共役受容体のひとつであるプロスタノイド受容体のキメラ受容体やアミノ酸変異受容体を作製してリガンド結合部位を同定してきた5,6).また,生体におけるプロスタノイドの役割を解明するためプロスタノイド受容体ノックアウトマウスに動脈硬化モデルマウスを掛け合わせてダブルノックアウトマウスを作出し,プロスタノイド受容体には動脈硬化の発症や進展を促進させる受容体と抑制させる受容体とがべつべつに存在することを明らかにした7).このことから,より効果的な治療を行うためには受容体に対する選択性の高いアゴニストやアンタゴニストを探索および開発する必要があった.そこで,おのおののGタンパク質共役受容体に選択的なリガンドをデザインしたいと考え,2000年から,Gタンパク質共役受容体のX線結晶構造解析を試みてきた.構造生物学的なアプローチにより,これまで分子生物学的,生化学的,薬理学的なアプローチでは得ることのできなかった新しい知見を得て創薬や生命現象の理解に供したいと考えている.
学習院大学理学部 元 教授.
Brian K. Kobilka
米国Stanford大学School of Medicine教授.
小林 拓也(Takuya Kobayashi)
略歴:1998年 京都大学大学院医学研究科博士課程 修了,同年 同 助手,2004年 英国Imperial College London博士研究員,2006年 科学技術振興機構ERATO岩田ヒト膜受容体構造プロジェクト グループリーダーを経て,2007年より京都大学大学院医学研究科 講師.
研究テーマ:Gタンパク質共役受容体の薬理学から構造生物学.
関心事:自分で解いたGタンパク質共役受容体の構造をもとに新しい薬をデザインし,いつの日かin vivoモデルで評価したい.
© 2012 芳賀達也・Brian K. Kobilka・小林拓也 Licensed under CC 表示 2.1 日本
(1学習院大学理学部 生命科学研究科,2米国Stanford大学School of Medicine,Department of Molecular and Cellular Physiology,3京都大学大学院医学研究科 分子細胞情報学)
email:小林拓也
DOI: 10.7875/first.author.2012.018
Structure of the human M2 muscarinic acetylcholine receptor bound to an antagonist.
Kazuko Haga, Andrew C. Kruse, Hidetsugu Asada, Takami Yurugi-Kobayashi, Mitsunori Shiroishi, Cheng Zhang, William I. Weis, Tetsuji Okada, Brian K. Kobilka, Tatsuya Haga, Takuya Kobayashi
Nature, 482, 547-551 (2012)
要 約
Gタンパク質共役受容体はヒトのゲノムにおいて非常に大きなファミリーを形成している.また,市販の医薬品の作用点の約40%をしめるともいわれており,創薬において重要なターゲットと位置づけられている.数多いGタンパク質共役受容体ファミリーのなかでも,ムスカリン受容体はオルソステリック部位とアロステリック部位とをもつといわれ,おのおのの部位を区別して結合する低分子化合物が報告されている.筆者らは,ヒトのムスカリン性アセチルコリン受容体M2サブタイプのオルソステリック部位にアンタゴニストが結合した複合体のX線結晶構造を解明した.ムスカリンM2受容体の立体構造が明らかになり,抗コリン薬の結合部位の詳細な情報を得ることができた.これらの立体構造の情報をもとに,今後,より効果的で副作用のないサブタイプに選択的な薬の探索および設計が可能になるものと考えられる.
はじめに
アセチルコリンは世界で最初に神経伝達物質のひとつであることが証明された物質である.アセチルコリンにはニコチン作用とムスカリン作用とが存在することが古くから知られており,おのおのに特異的なアセチルコリン受容体(ニコチン受容体とムスカリン受容体)が同定されている.副交感神経や運動神経の末端から分泌されたアセチルコリンは骨格筋や内臓平滑筋などに存在するアセチルコリン受容体にはたらき,筋収縮を促進するほか副交感神経を刺激し,心拍数の減少,脈拍数の減少,唾液の分泌を促進する.このように,アセチルコリンは基本的な生命現象に深く関与している.とくに,脳内のアセチルコリンのバランスがくずれるとさまざまな疾患が生じる.たとえば,アセチルコリンが減少すると自律神経失調症やアルツハイマー病につながるといわれている.また,パーキンソン病の患者ではアセチルコリンの相対的な増加が認められている.アセチルコリンのムスカリン受容体への結合を遮断することでパーキンソン病の症状が緩和することが知られており,古くから治療薬として使われている.一方で,副作用として統合失調症や認知症の症状を悪化させることも知られている.
ムスカリン受容体にはM1からM5までの5種類のサブタイプが存在する1).たとえば,M1受容体は大脳皮質や海馬に多く存在し記憶や学習に関与していると考えられている.また,M2受容体はおもに中枢神経や心臓に分布しその機能を抑制的に制御している.このように,おのおののサブタイプは固有の分布と薬理作用をもっており,構造にもとづいたサブタイプに選択的な創薬が期待されている.
1.Gタンパク質共役受容体のX線結晶構造解析のむずかしい点
膜タンパク質は市販の医薬品の作用点の50%以上をしめるといわれており,創薬において重要なターゲットと位置づけられている.そのなかでもGタンパク質共役型受容体は創薬の標的として注目されている.“構造にもとづく創薬戦略”によりGタンパク質共役受容体の立体構造を体系的かつ網羅的に解明していくことは,候補化合物を迅速に探索する手法の開発,候補化合物の最適化,副作用の少ない化合物の分子設計につながる.しかし,Gタンパク質共役受容体の構造解析は非常にむずかしく,長いあいだ,その構造は解かれていなかった.その理由として,非常に疎水性領域が多いため(全体の70~80%)界面活性剤で可溶化するとそのほとんどがミセルでおおわれてしまい結晶格子が形成されにくいこと,そして,活性型と不活性型の構造をとりうる柔軟性の高い膜タンパク質であることが考えられていた.
2.X線結晶構造解析に適したGタンパク質共役受容体の生産
筆者らは2000年から,ヒトをターゲットとしたGタンパク質共役受容体のX線結晶構造解析を目的として結晶化に適した試料の調製を試みてきた.その結果,ヒトGタンパク質共役受容体のうちの3つ,アデノシンA2a受容体,ヒスタミンH1受容体,ムスカリンM2受容体の結晶構造を高分解能で解明することに成功した.アデノシンA2a受容体についてはその立体構造を認識する抗体を作製して抗体との複合体の結晶構造を3.1Åで決定した2)(新着論文レビュー でも掲載).さらに,ヒスタミンH1受容体についてはリゾチームを融合したうえで脂質キュービック相に結晶化し高分解能(3.1Å)でその構造を解いた3).今回,ムスカリンM2受容体についても同様な方法を使い分解能3.0Åで結晶構造を決定した.ヒスタミンH1受容体およびアデノシンA2a受容体はメタノール資化酵母を用い発現させたのち精製したが,ムスカリンM2受容体は昆虫細胞を用いて調製した.よい結晶を生産するには,いずれの場合も培地1リットルあたり1 mg以上の生産量が必要となった.Gタンパク質共役受容体の高発現を実現するためには,柔軟性の高いGタンパク質共役受容体を安定化する必要があった.従来は,多くの変異体を地道に作製しそのリガンド結合活性を測定していたが,相同組換え活性の高い出芽酵母を用いC末端にGFPを融合させることにより短時間に安定性を向上させる系を確立した.現在,1カ月で50~100種類以上の変異体をハイスループットでスクリーニングしている.この技術により,まさに“解けるものを解く研究”から“解きたいものを解く研究”が可能になったといえる.
3.ヒトのムスカリン性アセチルコリン受容体M2サブタイプの結晶構造
Gタンパク質共役受容体ファミリーのなかでも,ムスカリン性アセチルコリン受容体は2つのリガンド結合部位(オルソステリック部位およびアロステリック部位)をかねそなえた特徴的な受容体である.Gタンパク質共役受容体のアロステリック制御についてはじめて報告されたのは比較的最近であり4),アロステリック部位の立体構造はこれまで明らかにされていなかった.筆者らは,今回,ヒトのムスカリン受容体M2サブタイプのオルソステリック部位にアンタゴニストが結合した複合体の結晶構造を世界ではじめて高分解能(3.0Å)で解くことに成功した(PDB ID:3UON).その結果,神経伝達物質であるアセチルコリンが結合するオルソステリック部位を構成する19個のアミノ酸残基がアンタゴニストであるキヌクリジニルベンジレートと作用しており(図1),そのうち18個のアミノ酸残基はすべてのムスカリン受容体サブタイプ(M1~M5)において保存されていた.このことより,ムスカリン受容体においてサブタイプに選択的に作用するリガンドの開発が非常にむずかしかったことを原子分解能レベルではじめて証明することができた.一方,アロステリック部位はオルソステリック部位の上部にある比較的広い空洞として細胞の外側に広がっていた(図2).アロステリック部位を構成するいくつかのアミノ酸残基は,アミノ酸変異実験によりアロステリックリガンドの結合親和性を低下させるアミノ酸残基と一致していた.アロステリック部位にはそれぞれのサブタイプにおいて特徴的なアミノ酸残基が存在していた.したがって,それぞれのサブタイプに特徴的なアミノ酸残基の側鎖とリガンドの官能基とのあいだに相互作用(水素結合など)を生み出すことができ,これはサブタイプに選択的なリガンドの開発につながると考えられた.
おわりに
筆者らはこれまでに,Gタンパク質共役受容体のひとつであるプロスタノイド受容体のキメラ受容体やアミノ酸変異受容体を作製してリガンド結合部位を同定してきた5,6).また,生体におけるプロスタノイドの役割を解明するためプロスタノイド受容体ノックアウトマウスに動脈硬化モデルマウスを掛け合わせてダブルノックアウトマウスを作出し,プロスタノイド受容体には動脈硬化の発症や進展を促進させる受容体と抑制させる受容体とがべつべつに存在することを明らかにした7).このことから,より効果的な治療を行うためには受容体に対する選択性の高いアゴニストやアンタゴニストを探索および開発する必要があった.そこで,おのおののGタンパク質共役受容体に選択的なリガンドをデザインしたいと考え,2000年から,Gタンパク質共役受容体のX線結晶構造解析を試みてきた.構造生物学的なアプローチにより,これまで分子生物学的,生化学的,薬理学的なアプローチでは得ることのできなかった新しい知見を得て創薬や生命現象の理解に供したいと考えている.
文 献
- Hulme, E. C., Birdsall, N. J. & Buckley, N. J.: Muscarinic receptor subtypes. Annu. Rev. Pharmacol. Toxicol., 30, 633-673 (1990)[PubMed]
- Hino, T., Arakawa, T., Iwanari, H. et al.: G-protein-coupled receptor inactivation by an allosteric inverse-agonist antibody. Nature, 482, 237-240 (2012)[PubMed] [新着論文レビュー]
- Shimamura, T., Shiroishi, M., Weyand, S. et al.: Structure of the human histamine H1 receptor complex with doxepin. Nature, 475, 65-70 (2011)[PubMed]
- Jensen, A. A. & Spalding, T. A.: Allosteric modulation of G-protein coupled receptors. Eur. J. Pharm. Sci., 21, 407-420 (2004)[PubMed]
- Kobayashi, T., Kiriyama, M., Hirata, T. et al.: Identification of domains conferring ligand binding specificity to the prostanoid receptor. Studies on chimeric prostacyclin/prostaglandin D receptors. J. Biol. Chem., 272, 15154-15160 (1997)[PubMed]
- Kobayashi, T., Ushikubi, F. & Narumiya, S.: Amino acid residues conferring ligand binding properties of prostaglandin I and prostaglandin D receptors. Identification by site-directed mutagenesis. J. Biol. Chem., 275, 24294-242303 (2000)[PubMed]
- Kobayashi, T., Tahara, Y., Matsumoto, M. et al.: Roles of thromboxane A2 and prostacyclin in the development of atheroscrelosis in apoE-deficient mice. J. Clin. Invest., 114, 784-794 (2004)[PubMed]
著者プロフィール
学習院大学理学部 元 教授.
Brian K. Kobilka
米国Stanford大学School of Medicine教授.
小林 拓也(Takuya Kobayashi)
略歴:1998年 京都大学大学院医学研究科博士課程 修了,同年 同 助手,2004年 英国Imperial College London博士研究員,2006年 科学技術振興機構ERATO岩田ヒト膜受容体構造プロジェクト グループリーダーを経て,2007年より京都大学大学院医学研究科 講師.
研究テーマ:Gタンパク質共役受容体の薬理学から構造生物学.
関心事:自分で解いたGタンパク質共役受容体の構造をもとに新しい薬をデザインし,いつの日かin vivoモデルで評価したい.
© 2012 芳賀達也・Brian K. Kobilka・小林拓也 Licensed under CC 表示 2.1 日本
