表現型の解析を基盤とした遺伝学的なスクリーニングを行うためのホモ変異体ES細胞バンクの構築
堀江恭二・竹田潤二
(大阪大学大学院医学系研究科 環境・生体機能学)
email:堀江恭二
DOI: 10.7875/first.author.2011.169
A homozygous mutant embryonic stem cell bank applicable for phenotype-driven genetic screening.
Kyoji Horie, Chikara Kokubu, Junko Yoshida, Keiko Akagi, Ayako Isotani, Akiko Oshitani, Kosuke Yusa, Ryuji Ikeda, Yue Huang, Allan Bradley, Junji Takeda
Nature Methods, 8, 1071-1077 (2011)
マウスES細胞を用いた変異導入法は遺伝子の機能を解析するうえで強力な手法である.しかし,表現型を示すには両方の対立遺伝子に変異を導入する必要があるため,迅速な解析は困難であった.筆者らは,以前,ヘテロ変異をもつマウスES細胞からホモ変異体を誘導する方法を報告したが,ホモ変異体のみを単離する方法がなかったため適用できる表現型はきわめてかぎられていた.そこで,ヘテロ変異体からホモ変異体を効率的に単離する方法を開発した.この手法では,変異遺伝子のコピー数の増加を検出するために新たに開発した遺伝子トラップ型ベクターと,変異部位に連鎖した1塩基多型の消失を利用した.そして,この手法により単離したホモ変異体を用いて,表現型の解析を基盤とした遺伝学的なスクリーニングの可能なことを実証した.
遺伝子を破壊してその表現型を調べる手法は,さまざまな遺伝子の機能解析に適用の可能な一般性の高い方法である.遺伝子破壊法は,特定の遺伝子にしぼって変異を導入し詳細に表現型を解析する逆遺伝学的な方法と,多数の遺伝子に対して変異を導入しそのなかから目的の表現型を示す変異体をスクリーニングする順遺伝学的な方法の2つに大別される.大腸菌や酵母などのモデル生物においては,いずれの方法も多用され必須の手法として確立している.一方,マウスにおいては,特定の遺伝子を改変した個体を作製するノックアウトマウス法,つまり,逆遺伝学的な方法が広く用いられており,多数の変異体を対象とした順遺伝学的な方法はたち遅れている.これは,マウスにおいてホモ変異体を得るには,ES細胞の改変からキメラマウスの作製および交配という多大な時間と労力を要す過程をへるため,多くの遺伝子に対し順遺伝学的な方法を適用するのは困難であるという事情による.一方,二本鎖RNAを導入して対応するmRNAを破壊するRNAi法は,操作が簡便なため順遺伝学的な方法と逆遺伝学的な方法のいずれにも用いることが可能だが,遺伝子機能を完全に破壊することは困難であり,かつ,目的以外の遺伝子の機能を阻害する可能性もある1).
筆者らは以前に,Bloom症候群の原因遺伝子であるBlm遺伝子の発現を一過性に抑制することにより,ヘテロ変異をもつマウスES細胞からホモ変異体を誘導する方法を報告した2).この手法をさらに発展させ,かつ,さきに述べた問題点を補完する方法として,両方の対立遺伝子を改変したホモ変異体ES細胞バンクを作製し,それを用いた培養細胞レベルでの遺伝子の機能解析を行った.ES細胞はさまざまな細胞系譜への分化誘導が可能であり,個体での生命現象を培養細胞のレベルでモデル化することができる.マウス個体での実験とは異なり,培養細胞のレベルであればひとりの研究者が多数の変異体を扱うことができるので,順遺伝学的なアプローチが可能になるものと期待できる.また,遺伝子自体が改変されているため,RNAi法とは異なり完全な遺伝子破壊を期待できる.
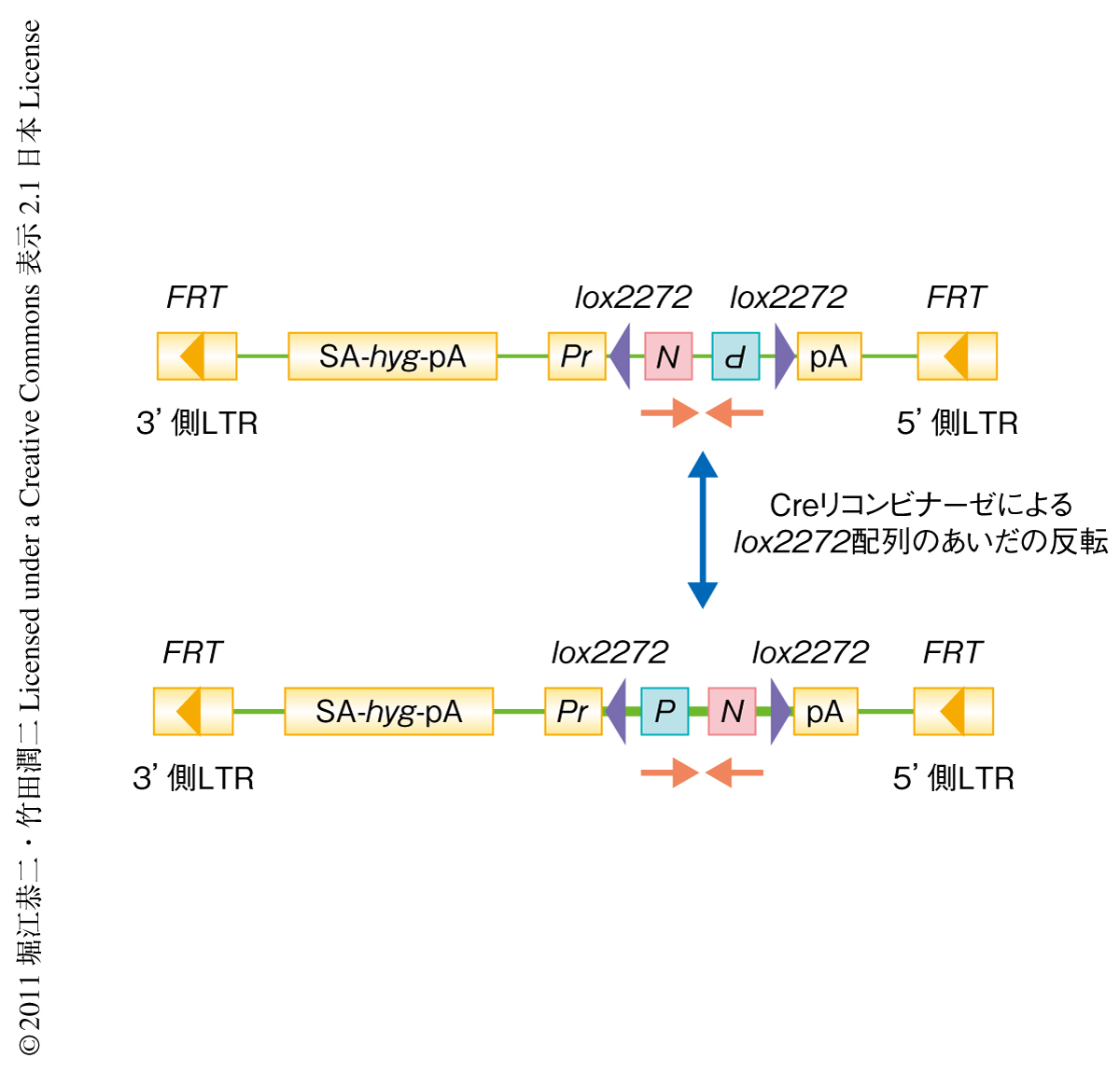
この手法で変異の導入に用いた遺伝子トラップ型ベクターは,発現している遺伝子へ挿入すると内在性の遺伝子からの転写産物がベクターのもつスプライシング受容部位によりハイグロマイシン耐性遺伝子へとスプライシングされ,ハイグロマイシン耐性株として選択される(図1).遺伝子トラップ型ベクターにはネオマイシン耐性遺伝子とピューロマイシン耐性遺伝子が逆方向をむいて配置されており,はじめはネオマイシン耐性かつピューロマイシン感受性である.これら薬剤耐性遺伝子のセットの両側には互いに逆向きのlox2272配列が存在する.このため,Creリコンビナーゼを一過性に作用させるとネオマイシン耐性遺伝子とピューロマイシン耐性遺伝子が一過性にくり返し反転し,ピューロマイシン耐性遺伝子とネオマイシン耐性遺伝子の向きが逆転した状態で固定化すると,まえとは逆にピューロマイシン耐性かつネオマイシン感受性となる.この原理は,あとで述べるホモ変異体の単離において利用する.また,この遺伝子トラップ型ベクターにはFRT配列が配置されており,FLPリコンビナーゼによりベクターの大部分を除去することが可能である.これにより表現型が消失すれば,遺伝子トラップ型ベクターの挿入が表現型の原因であることを確定できる.
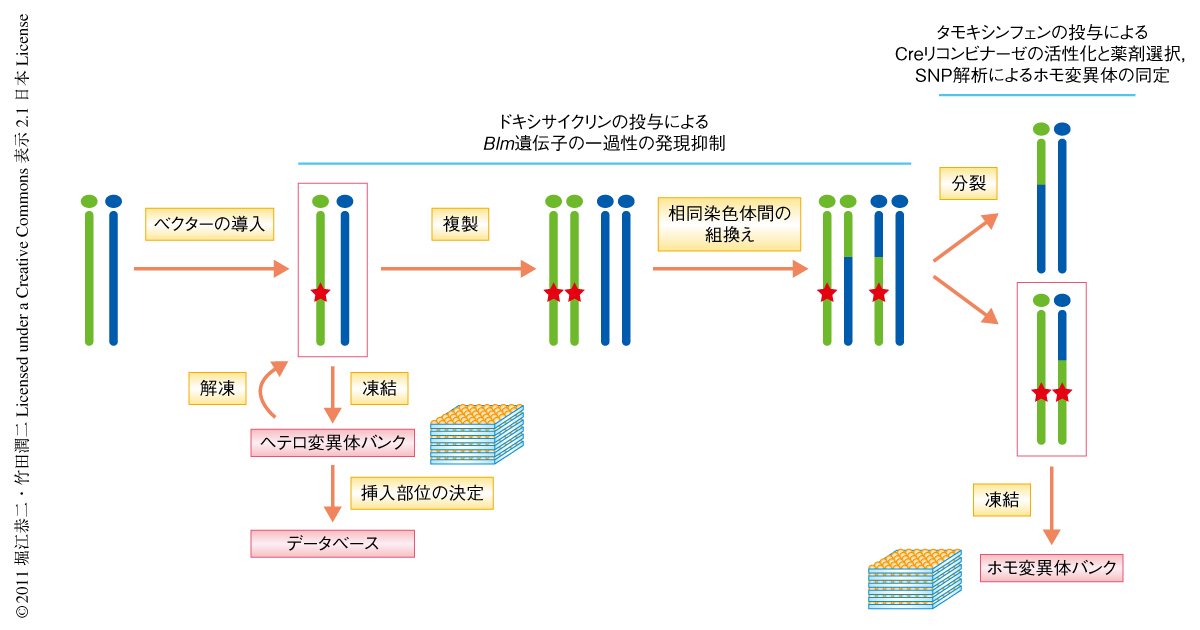
この遺伝子トラップ型ベクターをマウスのES細胞へ導入し,ES細胞で発現している遺伝子のヘテロ変異体をハイグロマイシン耐性株として単離した(図2).このとき,細胞あたり遺伝子トラップ型ベクターが1コピーのみ挿入する条件を用いた.ヘテロ変異体を単離し凍結保存したのち,ゲノムDNAを抽出して遺伝子トラップ型ベクターの挿入部位を決定し,約1000個のヘテロ変異体に関するデータベースを構築した.このヘテロ変異体を解凍したのち,あらかじめドキシサイクリンの投与による発現制御を可能にしておいたBlm遺伝子を4日間にわたり発現抑制した.このあいだに相同染色体のあいだの組換え頻度が亢進すると考えられ,そののちの細胞分裂の過程で変異の導入された染色体どうしが同じ娘細胞へひき継がれるとホモ変異体が出現する.
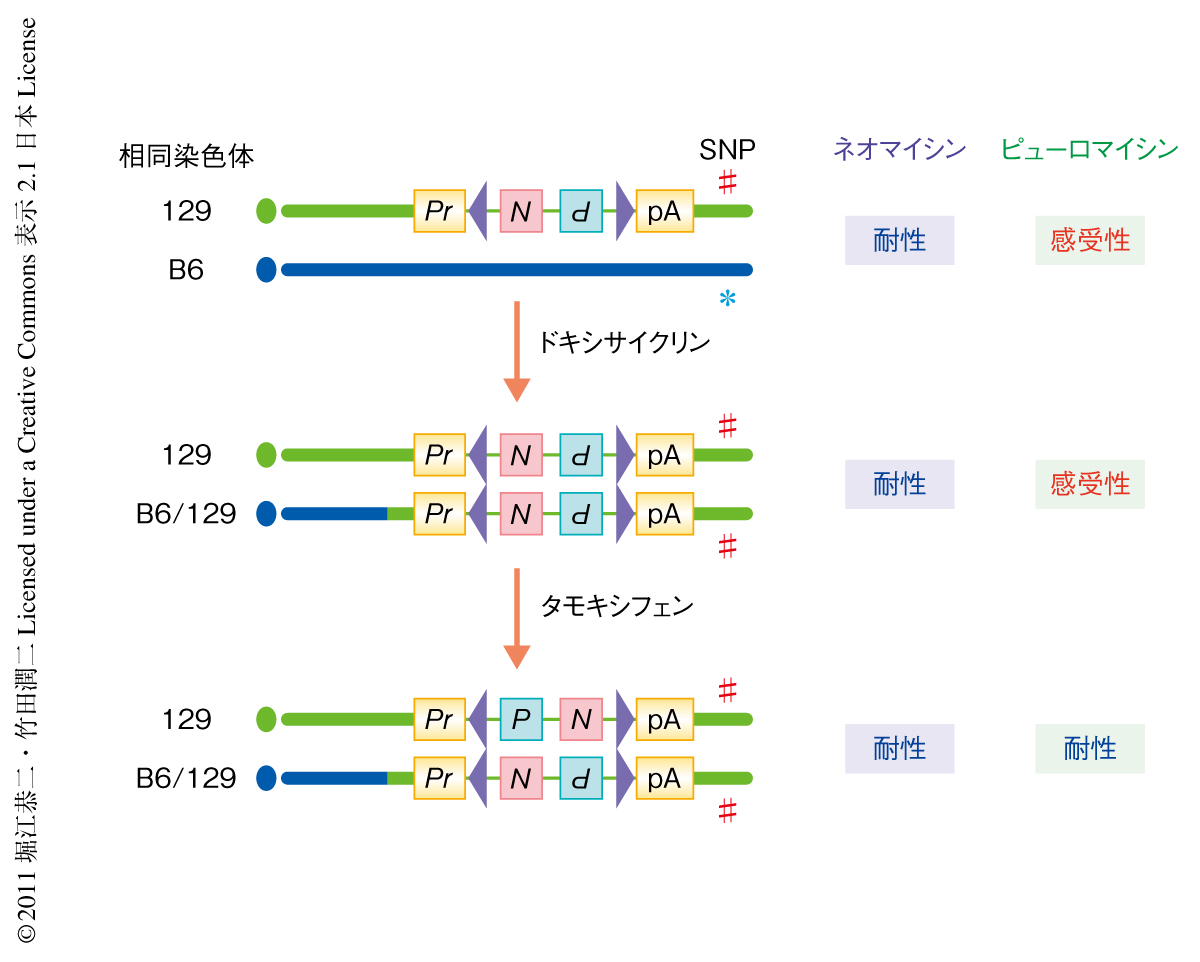
筆者らの過去の実験では,Blm遺伝子を発現抑制したときの特定の遺伝子座におけるホモ変異体の出現頻度は細胞の世代あたり2.3×10-4であり2),培養皿の大部分の細胞は依然としてヘテロ変異体でしめられているものと思われた.ここで,あらかじめタモキシフェンの投与による発現制御を可能にしておいた,Rosa26遺伝子座へノックインしたCreリコンビナーゼを一過性に活性化すると,さきに述べたように,遺伝子トラップ型ベクターに配置したlox2272配列のあいだで組換えが生じ,ネオマイシン耐性遺伝子とピューロマイシン耐性遺伝子のセットが反転する.その結果,ホモ変異体がもつ2コピーの遺伝子トラップ型ベクターのうち,一方からはネオマイシン耐性遺伝子が,もう一方からはピューロマイシン耐性遺伝子が発現する状態が生じ,両方の薬剤に対し耐性を示すようになる(図3).一方,ヘテロ変異体は遺伝子トラップ型ベクターを1コピーしかもたないため,いずれかの薬剤にしか耐性を示さない.このため,ネオマイシンおよびピューロマイシンの投与によりホモ変異体を選択できる(図3).ただし,実験の過程で,両方の薬剤に対する耐性株のうち約9割ではホモ変異体の誘導以外の要因で遺伝子トラップ型ベクターのもつ薬剤耐性遺伝子カセットが増加していることがわかり,ホモ変異体の特定のための障壁となった.そこで,遺伝子トラップ型ベクターが挿入した染色体においてテロメア側の1塩基多型(single nucleotide polymorphism:SNP)の多型性を解析し,多型性が失われた細胞株をホモ変異体の候補として同定することにした(図3).SNPの解析は薬剤耐性コロニーの単離から1日以内で結果を得ることが可能であった.最終的には,遺伝子トラップ型ベクターの挿入部位においてPCR法により野生型の対立遺伝子の消失を確認し,ホモ変異体ES細胞バンクを構築した.
約1000株からなるヘテロ変異体ES細胞バンクを構築したのち,そこからホモ変異体の誘導および単離を進めた.ホモ変異体を誘導する遺伝子の選定にあたっては,詳細な解析がなされている遺伝子やノックアウトマウスにおいて非致死性となる遺伝子はさけ,機能未知の遺伝子やノックアウトマウスにおいて胎生致死となる遺伝子を優先した.347個のヘテロ変異株について,Blm遺伝子の一過性の発現抑制と薬剤選択,SNP解析を行った結果,225株については変異遺伝子と連鎖したSNPに関するホモ変異体を取得できた.そのなかから165株について,遺伝子トラップ型ベクターの挿入部位における遺伝子型をPCR法により調べ,138株についてホモ変異体を特定した.よって,ヘテロ変異体からホモ変異体を取得する効率は (225/347)×(138/165) となり約55%であった.単離できなかった約45%のなかにはホモ変異により細胞増殖が低下したものや細胞死をきたしたものなどが存在すると考えられたが,それらがすべてとは考えにくい.ホモ変異体ではなかったES細胞の解析から染色体数の増加した細胞も検出されたことから,ホモ変異体の出現以外の要因でも薬剤耐性遺伝子カセットが2コピー以上に増加したES細胞が出現しうることは明らかであり,今後,この手法を改良するため重要な点と考えられた.
この手法の有効性を調べるため,まず,すでに変異体の表現型の報告されている遺伝子についてホモ変異体を単離して表現型を解析した.Dgcr8遺伝子はマイクロRNAの生合成に必須の遺伝子である.この手法で単離されたホモ変異体では,発現アレイの解析によりマイクロRNAの発現低下が示され,また,すでに報告されているように,ES細胞の増殖速度も低下した.また,Ptpn11遺伝子の欠損ではES細胞が分化抵抗性を示すことが報告されているが,これについてもこの手法で単離されたホモ変異体において再現できた.
ホモ変異体ES細胞バンクからランダムに変異体を選んで,分化能や細胞増殖能に異常を示す変異体のスクリーニングを試みた.その結果,神経系への分化抵抗性を示すものとしてAxin1遺伝子のホモ変異体を同定した.また,ES細胞の自己増殖能の低下性を示すものとしてCsnk2b遺伝子のホモ変異体やIlf2遺伝子のホモ変異体を同定した.これらの表現型はいずれもFLPリコンビナーゼにより遺伝子トラップ型ベクターの配列を除去することで消失し,ベクターの挿入が原因であることが確認された.Ilf2はES細胞の未分化性の維持に重要なNanogに結合することが報告されており,Nanogによる未分化の維持機構を明らかにするうえで有用である可能性が示唆された3).
この論文で報告した方法は,ヘテロ変異体からホモ変異体の単離を単純なプロトコールで行えるため,ひとりの実験者あたり年間100個程度の遺伝子についてホモ変異体を単離することができると考えられ,従来法に比べはるかに迅速であった.ただし,約2万個ある遺伝子に対し網羅的な遺伝子の機能解析を行うためにはさらなる改良を要する.そのためには,薬剤耐性遺伝子カセットのコピー数の安定性を高めることや,Blm遺伝子を発現抑制したときの相同染色体のあいだの組換えの頻度を高めるくふうが必要であろう.
ES細胞をさまざまな細胞系譜へと分化させる実験は,再生医学への応用の期待のもと,近年,きわめて精力的に行われている.とりわけ,iPS細胞の作製が報告されて以来,この潮流は加速しており,培養細胞レベルで個体を模した実験系は将来的により広がりをもつものと考えられる.ノックアウトマウス実験を加速するため,現在,すべて遺伝子についてヘテロ変異体ES細胞バンクを作製する国際プロジェクトが進行している4).筆者らが作製しているホモ変異体ES細胞のリソースは決して個体レベルの実験に置き換わるものではないが,培養細胞ならではの柔軟な実験が可能な点で相補的なリソースになるものと考えている.また,この手法はヒトのES細胞あるいはiPS細胞へも応用が可能である.ヒトにおいては個体レベルでの実験が困難なため,培養細胞レベルでのモデル実験の発展が強く望まれている.将来的には,ヒトのES細胞およびiPS細胞においてもこの手法を適用し,ヒトにおける実験遺伝学の可能性を広げていきたい.
なお,この研究で得たホモ変異体ES細胞は,さまざまな分野の研究者へのリソースとすることを目的に,Exchangeable Gene Trap Clones(EGTC)データベース(http://egtc.jp/action/main/index)をとおして,International Gene Trap Consortium(IGTC,http://www.genetrap.org/)への登録を進めている.筆者らの研究室のホームページ(http://www.med.osaka-u.ac.jp/pub/mr-envi/www/japanese/index.html)からも配布可能な細胞のリストを公開しており,今後,更新していく予定である.
略歴:1995年 大阪大学医学系研究科博士課程 修了,同年 米国NIHポストドクトラルフェロー,2000年 大阪大学先端科学技術共同研究センター 助手を経て,2007年より大阪大学医学系研究科 准教授.
研究テーマ:マウスおよびヒトの遺伝子の機能解析のための分子遺伝学的な手法の開発,ES細胞およびiPS細胞の多分化能の制御機構の解析.
竹田 潤二(Junji Takeda)
大阪大学医学系研究科 教授.
研究室URL:http://www.med.osaka-u.ac.jp/pub/mr-envi/www/japanese/index.html
© 2011 堀江恭二・竹田潤二 Licensed under CC 表示 2.1 日本
(大阪大学大学院医学系研究科 環境・生体機能学)
email:堀江恭二
DOI: 10.7875/first.author.2011.169
A homozygous mutant embryonic stem cell bank applicable for phenotype-driven genetic screening.
Kyoji Horie, Chikara Kokubu, Junko Yoshida, Keiko Akagi, Ayako Isotani, Akiko Oshitani, Kosuke Yusa, Ryuji Ikeda, Yue Huang, Allan Bradley, Junji Takeda
Nature Methods, 8, 1071-1077 (2011)
要 約
マウスES細胞を用いた変異導入法は遺伝子の機能を解析するうえで強力な手法である.しかし,表現型を示すには両方の対立遺伝子に変異を導入する必要があるため,迅速な解析は困難であった.筆者らは,以前,ヘテロ変異をもつマウスES細胞からホモ変異体を誘導する方法を報告したが,ホモ変異体のみを単離する方法がなかったため適用できる表現型はきわめてかぎられていた.そこで,ヘテロ変異体からホモ変異体を効率的に単離する方法を開発した.この手法では,変異遺伝子のコピー数の増加を検出するために新たに開発した遺伝子トラップ型ベクターと,変異部位に連鎖した1塩基多型の消失を利用した.そして,この手法により単離したホモ変異体を用いて,表現型の解析を基盤とした遺伝学的なスクリーニングの可能なことを実証した.
はじめに
遺伝子を破壊してその表現型を調べる手法は,さまざまな遺伝子の機能解析に適用の可能な一般性の高い方法である.遺伝子破壊法は,特定の遺伝子にしぼって変異を導入し詳細に表現型を解析する逆遺伝学的な方法と,多数の遺伝子に対して変異を導入しそのなかから目的の表現型を示す変異体をスクリーニングする順遺伝学的な方法の2つに大別される.大腸菌や酵母などのモデル生物においては,いずれの方法も多用され必須の手法として確立している.一方,マウスにおいては,特定の遺伝子を改変した個体を作製するノックアウトマウス法,つまり,逆遺伝学的な方法が広く用いられており,多数の変異体を対象とした順遺伝学的な方法はたち遅れている.これは,マウスにおいてホモ変異体を得るには,ES細胞の改変からキメラマウスの作製および交配という多大な時間と労力を要す過程をへるため,多くの遺伝子に対し順遺伝学的な方法を適用するのは困難であるという事情による.一方,二本鎖RNAを導入して対応するmRNAを破壊するRNAi法は,操作が簡便なため順遺伝学的な方法と逆遺伝学的な方法のいずれにも用いることが可能だが,遺伝子機能を完全に破壊することは困難であり,かつ,目的以外の遺伝子の機能を阻害する可能性もある1).
筆者らは以前に,Bloom症候群の原因遺伝子であるBlm遺伝子の発現を一過性に抑制することにより,ヘテロ変異をもつマウスES細胞からホモ変異体を誘導する方法を報告した2).この手法をさらに発展させ,かつ,さきに述べた問題点を補完する方法として,両方の対立遺伝子を改変したホモ変異体ES細胞バンクを作製し,それを用いた培養細胞レベルでの遺伝子の機能解析を行った.ES細胞はさまざまな細胞系譜への分化誘導が可能であり,個体での生命現象を培養細胞のレベルでモデル化することができる.マウス個体での実験とは異なり,培養細胞のレベルであればひとりの研究者が多数の変異体を扱うことができるので,順遺伝学的なアプローチが可能になるものと期待できる.また,遺伝子自体が改変されているため,RNAi法とは異なり完全な遺伝子破壊を期待できる.
1.マウスにおけるホモ変異体の取得の原理
この手法で変異の導入に用いた遺伝子トラップ型ベクターは,発現している遺伝子へ挿入すると内在性の遺伝子からの転写産物がベクターのもつスプライシング受容部位によりハイグロマイシン耐性遺伝子へとスプライシングされ,ハイグロマイシン耐性株として選択される(図1).遺伝子トラップ型ベクターにはネオマイシン耐性遺伝子とピューロマイシン耐性遺伝子が逆方向をむいて配置されており,はじめはネオマイシン耐性かつピューロマイシン感受性である.これら薬剤耐性遺伝子のセットの両側には互いに逆向きのlox2272配列が存在する.このため,Creリコンビナーゼを一過性に作用させるとネオマイシン耐性遺伝子とピューロマイシン耐性遺伝子が一過性にくり返し反転し,ピューロマイシン耐性遺伝子とネオマイシン耐性遺伝子の向きが逆転した状態で固定化すると,まえとは逆にピューロマイシン耐性かつネオマイシン感受性となる.この原理は,あとで述べるホモ変異体の単離において利用する.また,この遺伝子トラップ型ベクターにはFRT配列が配置されており,FLPリコンビナーゼによりベクターの大部分を除去することが可能である.これにより表現型が消失すれば,遺伝子トラップ型ベクターの挿入が表現型の原因であることを確定できる.
この遺伝子トラップ型ベクターをマウスのES細胞へ導入し,ES細胞で発現している遺伝子のヘテロ変異体をハイグロマイシン耐性株として単離した(図2).このとき,細胞あたり遺伝子トラップ型ベクターが1コピーのみ挿入する条件を用いた.ヘテロ変異体を単離し凍結保存したのち,ゲノムDNAを抽出して遺伝子トラップ型ベクターの挿入部位を決定し,約1000個のヘテロ変異体に関するデータベースを構築した.このヘテロ変異体を解凍したのち,あらかじめドキシサイクリンの投与による発現制御を可能にしておいたBlm遺伝子を4日間にわたり発現抑制した.このあいだに相同染色体のあいだの組換え頻度が亢進すると考えられ,そののちの細胞分裂の過程で変異の導入された染色体どうしが同じ娘細胞へひき継がれるとホモ変異体が出現する.
筆者らの過去の実験では,Blm遺伝子を発現抑制したときの特定の遺伝子座におけるホモ変異体の出現頻度は細胞の世代あたり2.3×10-4であり2),培養皿の大部分の細胞は依然としてヘテロ変異体でしめられているものと思われた.ここで,あらかじめタモキシフェンの投与による発現制御を可能にしておいた,Rosa26遺伝子座へノックインしたCreリコンビナーゼを一過性に活性化すると,さきに述べたように,遺伝子トラップ型ベクターに配置したlox2272配列のあいだで組換えが生じ,ネオマイシン耐性遺伝子とピューロマイシン耐性遺伝子のセットが反転する.その結果,ホモ変異体がもつ2コピーの遺伝子トラップ型ベクターのうち,一方からはネオマイシン耐性遺伝子が,もう一方からはピューロマイシン耐性遺伝子が発現する状態が生じ,両方の薬剤に対し耐性を示すようになる(図3).一方,ヘテロ変異体は遺伝子トラップ型ベクターを1コピーしかもたないため,いずれかの薬剤にしか耐性を示さない.このため,ネオマイシンおよびピューロマイシンの投与によりホモ変異体を選択できる(図3).ただし,実験の過程で,両方の薬剤に対する耐性株のうち約9割ではホモ変異体の誘導以外の要因で遺伝子トラップ型ベクターのもつ薬剤耐性遺伝子カセットが増加していることがわかり,ホモ変異体の特定のための障壁となった.そこで,遺伝子トラップ型ベクターが挿入した染色体においてテロメア側の1塩基多型(single nucleotide polymorphism:SNP)の多型性を解析し,多型性が失われた細胞株をホモ変異体の候補として同定することにした(図3).SNPの解析は薬剤耐性コロニーの単離から1日以内で結果を得ることが可能であった.最終的には,遺伝子トラップ型ベクターの挿入部位においてPCR法により野生型の対立遺伝子の消失を確認し,ホモ変異体ES細胞バンクを構築した.
2.ホモ変異体ES細胞バンクの構築
約1000株からなるヘテロ変異体ES細胞バンクを構築したのち,そこからホモ変異体の誘導および単離を進めた.ホモ変異体を誘導する遺伝子の選定にあたっては,詳細な解析がなされている遺伝子やノックアウトマウスにおいて非致死性となる遺伝子はさけ,機能未知の遺伝子やノックアウトマウスにおいて胎生致死となる遺伝子を優先した.347個のヘテロ変異株について,Blm遺伝子の一過性の発現抑制と薬剤選択,SNP解析を行った結果,225株については変異遺伝子と連鎖したSNPに関するホモ変異体を取得できた.そのなかから165株について,遺伝子トラップ型ベクターの挿入部位における遺伝子型をPCR法により調べ,138株についてホモ変異体を特定した.よって,ヘテロ変異体からホモ変異体を取得する効率は (225/347)×(138/165) となり約55%であった.単離できなかった約45%のなかにはホモ変異により細胞増殖が低下したものや細胞死をきたしたものなどが存在すると考えられたが,それらがすべてとは考えにくい.ホモ変異体ではなかったES細胞の解析から染色体数の増加した細胞も検出されたことから,ホモ変異体の出現以外の要因でも薬剤耐性遺伝子カセットが2コピー以上に増加したES細胞が出現しうることは明らかであり,今後,この手法を改良するため重要な点と考えられた.
3.順遺伝学的なアプローチによる表現型の解析の例
この手法の有効性を調べるため,まず,すでに変異体の表現型の報告されている遺伝子についてホモ変異体を単離して表現型を解析した.Dgcr8遺伝子はマイクロRNAの生合成に必須の遺伝子である.この手法で単離されたホモ変異体では,発現アレイの解析によりマイクロRNAの発現低下が示され,また,すでに報告されているように,ES細胞の増殖速度も低下した.また,Ptpn11遺伝子の欠損ではES細胞が分化抵抗性を示すことが報告されているが,これについてもこの手法で単離されたホモ変異体において再現できた.
ホモ変異体ES細胞バンクからランダムに変異体を選んで,分化能や細胞増殖能に異常を示す変異体のスクリーニングを試みた.その結果,神経系への分化抵抗性を示すものとしてAxin1遺伝子のホモ変異体を同定した.また,ES細胞の自己増殖能の低下性を示すものとしてCsnk2b遺伝子のホモ変異体やIlf2遺伝子のホモ変異体を同定した.これらの表現型はいずれもFLPリコンビナーゼにより遺伝子トラップ型ベクターの配列を除去することで消失し,ベクターの挿入が原因であることが確認された.Ilf2はES細胞の未分化性の維持に重要なNanogに結合することが報告されており,Nanogによる未分化の維持機構を明らかにするうえで有用である可能性が示唆された3).
おわりに
この論文で報告した方法は,ヘテロ変異体からホモ変異体の単離を単純なプロトコールで行えるため,ひとりの実験者あたり年間100個程度の遺伝子についてホモ変異体を単離することができると考えられ,従来法に比べはるかに迅速であった.ただし,約2万個ある遺伝子に対し網羅的な遺伝子の機能解析を行うためにはさらなる改良を要する.そのためには,薬剤耐性遺伝子カセットのコピー数の安定性を高めることや,Blm遺伝子を発現抑制したときの相同染色体のあいだの組換えの頻度を高めるくふうが必要であろう.
ES細胞をさまざまな細胞系譜へと分化させる実験は,再生医学への応用の期待のもと,近年,きわめて精力的に行われている.とりわけ,iPS細胞の作製が報告されて以来,この潮流は加速しており,培養細胞レベルで個体を模した実験系は将来的により広がりをもつものと考えられる.ノックアウトマウス実験を加速するため,現在,すべて遺伝子についてヘテロ変異体ES細胞バンクを作製する国際プロジェクトが進行している4).筆者らが作製しているホモ変異体ES細胞のリソースは決して個体レベルの実験に置き換わるものではないが,培養細胞ならではの柔軟な実験が可能な点で相補的なリソースになるものと考えている.また,この手法はヒトのES細胞あるいはiPS細胞へも応用が可能である.ヒトにおいては個体レベルでの実験が困難なため,培養細胞レベルでのモデル実験の発展が強く望まれている.将来的には,ヒトのES細胞およびiPS細胞においてもこの手法を適用し,ヒトにおける実験遺伝学の可能性を広げていきたい.
なお,この研究で得たホモ変異体ES細胞は,さまざまな分野の研究者へのリソースとすることを目的に,Exchangeable Gene Trap Clones(EGTC)データベース(http://egtc.jp/action/main/index)をとおして,International Gene Trap Consortium(IGTC,http://www.genetrap.org/)への登録を進めている.筆者らの研究室のホームページ(http://www.med.osaka-u.ac.jp/pub/mr-envi/www/japanese/index.html)からも配布可能な細胞のリストを公開しており,今後,更新していく予定である.
文 献
- Birmingham, A., Anderson, E. M., Reynolds, A. et al.: 3' UTR seed matches, but not overall identity, are associated with RNAi off-targets. Nat. Methods, 3, 199-204 (2006)[PubMed]
- Yusa, K., Horie, K., Kondoh, G. et al.: Genome-wide phenotype analysis in ES cells by regulated disruption of Bloom’s syndrome gene. Nature, 429, 896-899 (2004)[PubMed]
- Lu, R., Markowetz, F., Unwin, R. D. et al.: Systems-level dynamic analyses of fate change in murine embryonic stem cells. Nature, 462, 358-362 (2009)[PubMed]
- Skarnes, W. C., Rosen, B., West, A. P. et al.: A conditional knockout resource for the genome-wide study of mouse gene function. Nature, 474, 338-342 (2011)[PubMed]
著者プロフィール
略歴:1995年 大阪大学医学系研究科博士課程 修了,同年 米国NIHポストドクトラルフェロー,2000年 大阪大学先端科学技術共同研究センター 助手を経て,2007年より大阪大学医学系研究科 准教授.
研究テーマ:マウスおよびヒトの遺伝子の機能解析のための分子遺伝学的な手法の開発,ES細胞およびiPS細胞の多分化能の制御機構の解析.
竹田 潤二(Junji Takeda)
大阪大学医学系研究科 教授.
研究室URL:http://www.med.osaka-u.ac.jp/pub/mr-envi/www/japanese/index.html
© 2011 堀江恭二・竹田潤二 Licensed under CC 表示 2.1 日本
