嗅球において新生したニューロンは食後に整理される
横山 健・持丸大輔・山口正洋
(東京大学大学院医学系研究科 細胞分子生理学教室)
email:横山 健,持丸大輔,山口正洋
DOI: 10.7875/first.author.2011.156
Elimination of adult-born neurons in the olfactory bulb is promoted during the postprandial period.
Takeshi K. Yokoyama, Daisuke Mochimaru, Koshi Murata, Hiroyuki Manabe, Ko Kobayakawa, Reiko Kobayakawa, Hitoshi Sakano, Kensaku Mori, Masahiro Yamaguchi
Neuron, 71, 883-897 (2011)
通常,哺乳類の成体の脳では新しいニューロンは生み出されないが,嗅覚一次中枢の嗅球ではニューロンが新生し神経回路の可塑性に寄与している.嗅球において新生した顆粒細胞のうち,約半数は既存の神経回路に組み込まれて利用されるが,残りは細胞死を起こし排除される.このような新生した顆粒細胞の整理が動物の行動様式にあわせいつ行われているのかはわかっていなかった.筆者らは,嗅球において新生した顆粒細胞の整理が食餌ののちの時間,典型的には食後の睡眠時に行われていることを見い出した.1日のうち特定の時間にのみ給餌したマウスでは,摂食行動とそののちの休眠行動の組合せにより新生した顆粒細胞の細胞死が促進された.数十分の食後の睡眠が新生した顆粒細胞の整理を促進した.匂い入力により新生した顆粒細胞の生死の調節されることが知られているが,匂い入力を遮断すると食後の休眠時の細胞死が著しく促進された.しかし,この時間帯のほかでは匂い入力を遮断しても細胞死の亢進はみられなかった.これらの結果から,摂食など覚醒時の嗅覚経験を反映して,そののちの休眠時に嗅球において新生した顆粒細胞の整理が行われることが明らかになった.この機構により匂い経験に即した嗅球の神経回路の再編が効率よく行われているものと考えられた.
発生期における神経回路の形成は,まず過剰なニューロンを準備し,そのなかから適切なニューロンを選別することにより行われる.古くは,交感神経節での交感ニューロンの選別から,神経筋接合部での運動ニューロンの選別,小脳プルキンエ細胞での登上繊維の選別まで,数多くの実例が明らかになっている.そして,この原理は成体の脳において新生したニューロンにおいてもあてはまる.嗅球における抑制性の介在ニューロンである顆粒細胞は,1日あたり実に顆粒細胞の総数の1%に相当する数が新しく供給されているが,そのうち約半数が生き残って既存の神経回路に組み込まれ,残りの半数は細胞死により排除されている1)(図1).このような新生した顆粒細胞の生死の選別は匂い経験により調節されており,新しい匂い経験は回路に組み込まれる顆粒細胞の割合を増し,逆に,匂い入力の欠如は排除される顆粒細胞の割合を増す1).このことから,新生した顆粒細胞は匂い経験に応じて必要なものと不要なものとに選別され,嗅球における匂い経験にもとづいた神経回路の再編および可塑的な変化に寄与しているものと考えられる.不要なニューロンを排除することは神経回路のノイズを落とし情報のチューニングを促進するとともに,新たなニューロンを組み入れる余地を確保するためにも重要であろう.実際,嗅球の細胞死を阻害すると正常な嗅覚機能の阻害されることが報告されている2).

このような嗅球における新生した顆粒細胞の整理は,いわば大規模な神経回路の構造変化である.この構造変化が1日のうちいつでも均等に起こっているのか,あるいは,特定の行動時間に起こっているのかがこの論文で取り組んだ疑問である.身近な例になぞらえるなら,われわれは自分の家に日々新しい品物(新聞,雑誌,食材,などなど)を運び入れているが,その片付けをいったいいつ行っているのか,いつ行うのが効率的なのか,という類の疑問である.動物は個体あるいは種の生存と維持にかかわる重要な行動を嗅覚に依存している.嗅球の神経回路は日々新しく生み出された顆粒細胞を適切に利用および整理し,嗅覚経験に即して回路をよりよいものに再編していくため効率のよい方法を採用しているのではないだろうか.
筆者らは,典型的な嗅覚行動である摂食行動に着目し,マウスに対し摂食制限により再現性よく摂食行動を起こさせるようにした.このマウスの嗅球における新生した顆粒細胞の細胞死は食後の休眠時に特異的に促進され,その細胞死の程度は匂い入力に依存して増減した.このことから,嗅球における匂い経験に依存的な新生した顆粒細胞の整理は,食餌行動および休眠行動という特定の行動様式にあわせて行われることが判明した.筆者らは,嗅球における新生した顆粒細胞の整理は,覚醒時の嗅覚入力によるニューロンの“タグづけ”と,そののちの休眠時に起こる仮想的な“再編シグナル”の入力との組合せにより起こるとする“2段階モデル”を提唱した.
嗅球における顆粒細胞の細胞死は,アポトーシスのシグナル経路を担うカスパーゼ3の活性化を免疫組織染色により検出することで評価した.カスパーゼ3が活性化したほとんどの顆粒細胞はDNAの断片化および核のアポトーシス様の形態変化をともなっていた.まず,マウスを12時間-12時間の明暗リズム,自由摂食のもとにおいて個別に飼育し嗅球における顆粒細胞の細胞死の日内変動を調べたところ,明らかな日内リズムは観察されなかったが,個体による変動が大きく,明暗リズムとは別の調節機構の存在が考えられた.
そこで,嗅球における顆粒細胞の細胞死がなんらかの嗅覚行動と関連しているのではないかと考え,典型的な嗅覚行動である摂食行動との関連を調べた.1日のうち4時間だけ餌をあたえる摂食制限によりマウスの摂食行動を管理したところ,食餌の時間に一致して嗅球における顆粒細胞の細胞死が促進された.食餌の開始まえに比べ,食餌の開始2時間後には細胞死を起こす顆粒細胞の数が2~3倍に増加した.一方,食餌の時間のほかでは細胞死は少ない値に抑えられていた.ブロモデオキシウリジン(BrdU)によるニューロンの標識実験から,食餌の時間に細胞死を起こす顆粒細胞の多くは新生したものであり,生死の決定に重要な日齢3) に相当するものであることを確認した.
食餌の時間におけるマウスの行動を観察すると,はじめの1時間は餌を食べることに集中していたが,つぎの1時間には毛づくろい,休憩,睡眠など典型的な食後行動をとっていた.おもしろいことに,顆粒細胞の細胞死は食餌の開始1時間後では増加せず,開始2時間後になって増加が認められ食後行動との関連が示唆された.そこで,マウスの食後行動(毛づくろい,休憩,睡眠)をおだやかなハンドリングで阻害したところ顆粒細胞の細胞死は促進されなくなった.
食後行動を脳波および筋電図の測定により客観的に評価したところ,ほとんどのマウスは食後に睡眠行動をとっており,睡眠時間と細胞死を起こす顆粒細胞数とのあいだには正の相関が認められた.食後の睡眠のほとんどは徐波睡眠であり(レム睡眠は少ない),徐波睡眠の時間と細胞死を起こす顆粒細胞数との相関が高かった.20~40分程度の短時間の睡眠が顕著な細胞死の増加と対応しており,“食後のうたた寝”が細胞死を強く誘導しうるものと考えられた.
食餌を制限したマウスは当然ながら食後のほかにも睡眠行動をとるが,その時間帯には顆粒細胞の細胞死は促進されなかった.これらのことから,睡眠の単独ではなく,摂食行動とそののちの睡眠との組合せが嗅球における新生した顆粒細胞の細胞死による整理に重要であると考えられた.
嗅球における食後での新生した顆粒細胞の細胞死が匂い入力によりどのように影響されているかを調べるため,マウスの片側鼻孔を慢性的に閉塞して匂い入力の遮断を行ったところ,遮断側の嗅球では食後での細胞死が著しく促進された.重要なのは,食後のほかには慢性的に匂い入力を遮断しても細胞死の増加が起こらなかった点である.このことは,食餌を制限したマウスにおいては嗅球における匂い入力に依存的な新生した顆粒細胞の整理は食後に特異的に行われ,それ以外の時間帯では起こらないことを示していた.
動物が経験する匂いはさまざまであり,嗅球には匂い経験の種類に対応した応答ドメインが存在する4).嗅覚入力を嗅球の背側ドメインでのみ欠損させたマウス5) では,匂い入力が保たれている腹側ドメインに比べ背側ドメインにおいて食後での新生した顆粒細胞の細胞死が著しく促進されていた.これは,食後での細胞死の程度が嗅球局所への匂い入力により嗅球局所において調節されていることを示していた.食後に嗅球における個別の神経回路が匂い入力に応じて異なる再編をうけるものと考えられた.
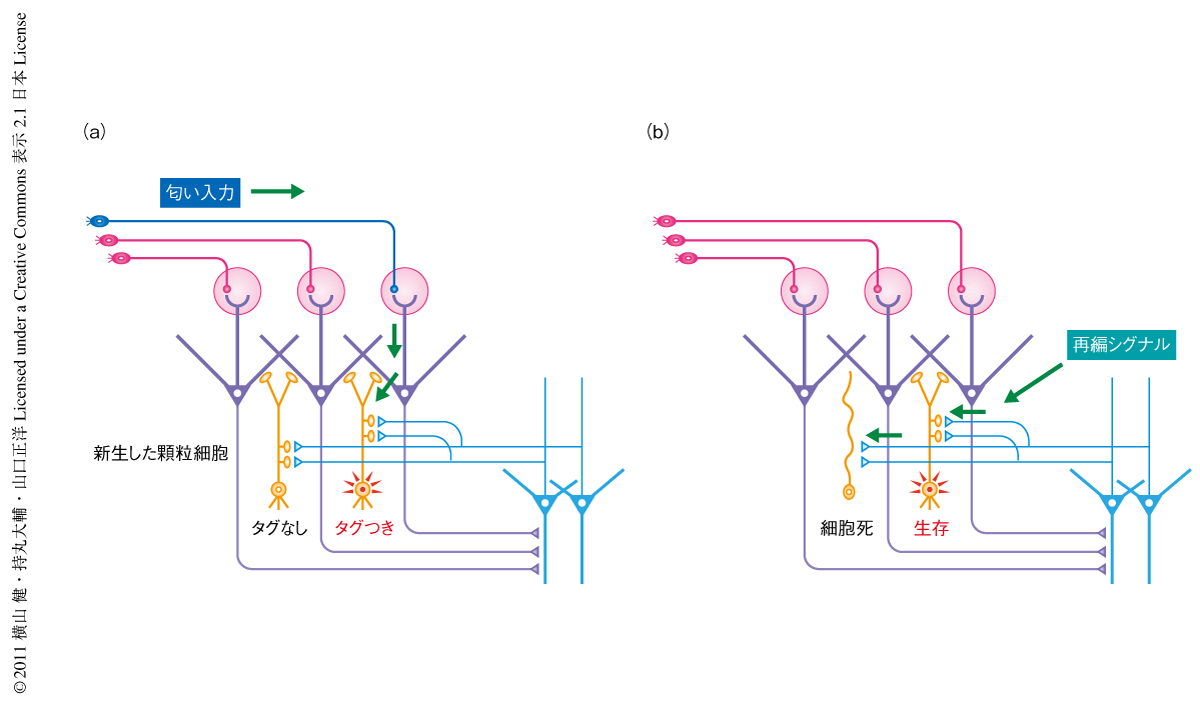
以上の結果から,嗅球における匂い経験に依存的な新生した顆粒細胞の選別の“2段階モデル”を提唱した(図2).2段階とは嗅覚行動時とそののちの休眠時という連続して起こる2つの異なる行動様式の組合せをさす.食餌行動のような嗅覚行動中に,新生した顆粒細胞のうちのあるものは匂い入力により活性化し“タグづけ”が行われる一方,あるものは活性化されずにタグづけが行われず,これにより新生した顆粒細胞の色分けがなされる.しかし,この時点ではタグづけされていない細胞はまだ排除されない.そして,のちの休眠時に,新生した顆粒細胞に仮想的な“再編シグナル”が入力される.このシグナルはタグづけされた顆粒細胞を生存させる一方,タグづけされていない顆粒細胞を細胞死により排除する.この2段階により嗅球における匂い入力に応じた新生した顆粒細胞の利用および整理が行われ,これは神経回路における適切な再編および可塑的な変化に貢献しているものと考えられる.この2段階モデルには,海馬における学習記憶の2段階モデル6) や,覚醒・睡眠行動にともなうシナプス機能のアップスケーリング・ダウンスケーリングモデル7) との共通点を見い出すことができる.
嗅球において新生した顆粒細胞の整理は一日中いつでも均等に起こっているのではなく,食後の休眠という特定の行動様式に対応して起こっていた.動物は日々新しい食べ物に出会い,それがおいしいかまずいか,体にとりよい栄養になったかどうか,など新たな学習をくり返している.このような食べ物の経験に応じ,のちの休眠時に新生した顆粒細胞を整理することで,動物は食べ物の情報を効率よく学習し記憶しているものと考えられる.
食餌行動は典型的な嗅覚行動であるが,嗅覚は天敵からの逃避,交尾相手の選択,なわばりの確保など個体の維持と種の繁栄にかかわるさまざまな行動に寄与している.筆者らは,これらの嗅覚行動のあとの休眠時間にも,嗅覚経験に応じた新生した顆粒細胞の利用および整理が促進されているのではないかと想定している.動物は嗅覚行動中には匂い情報,つまり,外界の匂い環境の情報を適切に処理することに集中しているのかもしれない.そして,のちの休眠時には外界の匂い情報の処理をいったん中断し,さきの匂い経験にもとづいて大規模な神経回路の再編を行うことに集中し,つぎに覚醒したときに,より適切な匂い行動をとれるよう備えているのではないだろうか.
最近,筆者らの研究室では,睡眠中には嗅球の上位中枢である嗅皮質から同期的なトップダウン性シナプス入力が嗅球にむかって入力することを見い出している8).この入力が嗅球において新生した顆粒細胞の整理を促進する“再編シグナル”の実体である可能性があり,現在,この点を検討中である.また,経験に依存的な“タグづけ”の機構は神経科学の全般に共通する大きな課題であり,嗅球の神経回路はその解明にむけて格好の題材を提供しているものと思われる.
略歴:2011年 東京大学大学院医学系研究科博士課程 単位取得退学,同年より東京大学 学術支援職員.
持丸 大輔(Daisuke Mochimaru)
略歴:東京大学大学院医学系研究科博士課程 在学中.
研究テーマ:成体の嗅球における新生ニューロンの生死の決定および組み込み機構の解明.
抱負:新生ニューロンが生き残り嗅球の神経回路の一部としてはたらくようになるには何が必要なのかを解明したい.
山口 正洋(Masahiro Yamaguchi)
東京大学大学院医学系研究科 講師.
© 2011 横山 健・持丸大輔・山口正洋 Licensed under CC 表示 2.1 日本
(東京大学大学院医学系研究科 細胞分子生理学教室)
email:横山 健,持丸大輔,山口正洋
DOI: 10.7875/first.author.2011.156
Elimination of adult-born neurons in the olfactory bulb is promoted during the postprandial period.
Takeshi K. Yokoyama, Daisuke Mochimaru, Koshi Murata, Hiroyuki Manabe, Ko Kobayakawa, Reiko Kobayakawa, Hitoshi Sakano, Kensaku Mori, Masahiro Yamaguchi
Neuron, 71, 883-897 (2011)
要 約
通常,哺乳類の成体の脳では新しいニューロンは生み出されないが,嗅覚一次中枢の嗅球ではニューロンが新生し神経回路の可塑性に寄与している.嗅球において新生した顆粒細胞のうち,約半数は既存の神経回路に組み込まれて利用されるが,残りは細胞死を起こし排除される.このような新生した顆粒細胞の整理が動物の行動様式にあわせいつ行われているのかはわかっていなかった.筆者らは,嗅球において新生した顆粒細胞の整理が食餌ののちの時間,典型的には食後の睡眠時に行われていることを見い出した.1日のうち特定の時間にのみ給餌したマウスでは,摂食行動とそののちの休眠行動の組合せにより新生した顆粒細胞の細胞死が促進された.数十分の食後の睡眠が新生した顆粒細胞の整理を促進した.匂い入力により新生した顆粒細胞の生死の調節されることが知られているが,匂い入力を遮断すると食後の休眠時の細胞死が著しく促進された.しかし,この時間帯のほかでは匂い入力を遮断しても細胞死の亢進はみられなかった.これらの結果から,摂食など覚醒時の嗅覚経験を反映して,そののちの休眠時に嗅球において新生した顆粒細胞の整理が行われることが明らかになった.この機構により匂い経験に即した嗅球の神経回路の再編が効率よく行われているものと考えられた.
はじめに
発生期における神経回路の形成は,まず過剰なニューロンを準備し,そのなかから適切なニューロンを選別することにより行われる.古くは,交感神経節での交感ニューロンの選別から,神経筋接合部での運動ニューロンの選別,小脳プルキンエ細胞での登上繊維の選別まで,数多くの実例が明らかになっている.そして,この原理は成体の脳において新生したニューロンにおいてもあてはまる.嗅球における抑制性の介在ニューロンである顆粒細胞は,1日あたり実に顆粒細胞の総数の1%に相当する数が新しく供給されているが,そのうち約半数が生き残って既存の神経回路に組み込まれ,残りの半数は細胞死により排除されている1)(図1).このような新生した顆粒細胞の生死の選別は匂い経験により調節されており,新しい匂い経験は回路に組み込まれる顆粒細胞の割合を増し,逆に,匂い入力の欠如は排除される顆粒細胞の割合を増す1).このことから,新生した顆粒細胞は匂い経験に応じて必要なものと不要なものとに選別され,嗅球における匂い経験にもとづいた神経回路の再編および可塑的な変化に寄与しているものと考えられる.不要なニューロンを排除することは神経回路のノイズを落とし情報のチューニングを促進するとともに,新たなニューロンを組み入れる余地を確保するためにも重要であろう.実際,嗅球の細胞死を阻害すると正常な嗅覚機能の阻害されることが報告されている2).
このような嗅球における新生した顆粒細胞の整理は,いわば大規模な神経回路の構造変化である.この構造変化が1日のうちいつでも均等に起こっているのか,あるいは,特定の行動時間に起こっているのかがこの論文で取り組んだ疑問である.身近な例になぞらえるなら,われわれは自分の家に日々新しい品物(新聞,雑誌,食材,などなど)を運び入れているが,その片付けをいったいいつ行っているのか,いつ行うのが効率的なのか,という類の疑問である.動物は個体あるいは種の生存と維持にかかわる重要な行動を嗅覚に依存している.嗅球の神経回路は日々新しく生み出された顆粒細胞を適切に利用および整理し,嗅覚経験に即して回路をよりよいものに再編していくため効率のよい方法を採用しているのではないだろうか.
筆者らは,典型的な嗅覚行動である摂食行動に着目し,マウスに対し摂食制限により再現性よく摂食行動を起こさせるようにした.このマウスの嗅球における新生した顆粒細胞の細胞死は食後の休眠時に特異的に促進され,その細胞死の程度は匂い入力に依存して増減した.このことから,嗅球における匂い経験に依存的な新生した顆粒細胞の整理は,食餌行動および休眠行動という特定の行動様式にあわせて行われることが判明した.筆者らは,嗅球における新生した顆粒細胞の整理は,覚醒時の嗅覚入力によるニューロンの“タグづけ”と,そののちの休眠時に起こる仮想的な“再編シグナル”の入力との組合せにより起こるとする“2段階モデル”を提唱した.
1.食餌を制限したマウスでは新生した顆粒細胞の細胞死は食餌の時間に促進される
嗅球における顆粒細胞の細胞死は,アポトーシスのシグナル経路を担うカスパーゼ3の活性化を免疫組織染色により検出することで評価した.カスパーゼ3が活性化したほとんどの顆粒細胞はDNAの断片化および核のアポトーシス様の形態変化をともなっていた.まず,マウスを12時間-12時間の明暗リズム,自由摂食のもとにおいて個別に飼育し嗅球における顆粒細胞の細胞死の日内変動を調べたところ,明らかな日内リズムは観察されなかったが,個体による変動が大きく,明暗リズムとは別の調節機構の存在が考えられた.
そこで,嗅球における顆粒細胞の細胞死がなんらかの嗅覚行動と関連しているのではないかと考え,典型的な嗅覚行動である摂食行動との関連を調べた.1日のうち4時間だけ餌をあたえる摂食制限によりマウスの摂食行動を管理したところ,食餌の時間に一致して嗅球における顆粒細胞の細胞死が促進された.食餌の開始まえに比べ,食餌の開始2時間後には細胞死を起こす顆粒細胞の数が2~3倍に増加した.一方,食餌の時間のほかでは細胞死は少ない値に抑えられていた.ブロモデオキシウリジン(BrdU)によるニューロンの標識実験から,食餌の時間に細胞死を起こす顆粒細胞の多くは新生したものであり,生死の決定に重要な日齢3) に相当するものであることを確認した.
2.新生した顆粒細胞の整理は食後の睡眠に関連して起こる
食餌の時間におけるマウスの行動を観察すると,はじめの1時間は餌を食べることに集中していたが,つぎの1時間には毛づくろい,休憩,睡眠など典型的な食後行動をとっていた.おもしろいことに,顆粒細胞の細胞死は食餌の開始1時間後では増加せず,開始2時間後になって増加が認められ食後行動との関連が示唆された.そこで,マウスの食後行動(毛づくろい,休憩,睡眠)をおだやかなハンドリングで阻害したところ顆粒細胞の細胞死は促進されなくなった.
食後行動を脳波および筋電図の測定により客観的に評価したところ,ほとんどのマウスは食後に睡眠行動をとっており,睡眠時間と細胞死を起こす顆粒細胞数とのあいだには正の相関が認められた.食後の睡眠のほとんどは徐波睡眠であり(レム睡眠は少ない),徐波睡眠の時間と細胞死を起こす顆粒細胞数との相関が高かった.20~40分程度の短時間の睡眠が顕著な細胞死の増加と対応しており,“食後のうたた寝”が細胞死を強く誘導しうるものと考えられた.
食餌を制限したマウスは当然ながら食後のほかにも睡眠行動をとるが,その時間帯には顆粒細胞の細胞死は促進されなかった.これらのことから,睡眠の単独ではなく,摂食行動とそののちの睡眠との組合せが嗅球における新生した顆粒細胞の細胞死による整理に重要であると考えられた.
3.食後における新生した顆粒細胞の細胞死の程度は匂い入力により調節される
嗅球における食後での新生した顆粒細胞の細胞死が匂い入力によりどのように影響されているかを調べるため,マウスの片側鼻孔を慢性的に閉塞して匂い入力の遮断を行ったところ,遮断側の嗅球では食後での細胞死が著しく促進された.重要なのは,食後のほかには慢性的に匂い入力を遮断しても細胞死の増加が起こらなかった点である.このことは,食餌を制限したマウスにおいては嗅球における匂い入力に依存的な新生した顆粒細胞の整理は食後に特異的に行われ,それ以外の時間帯では起こらないことを示していた.
動物が経験する匂いはさまざまであり,嗅球には匂い経験の種類に対応した応答ドメインが存在する4).嗅覚入力を嗅球の背側ドメインでのみ欠損させたマウス5) では,匂い入力が保たれている腹側ドメインに比べ背側ドメインにおいて食後での新生した顆粒細胞の細胞死が著しく促進されていた.これは,食後での細胞死の程度が嗅球局所への匂い入力により嗅球局所において調節されていることを示していた.食後に嗅球における個別の神経回路が匂い入力に応じて異なる再編をうけるものと考えられた.
4.嗅球における新生した顆粒細胞の選別の“2段階モデル”
以上の結果から,嗅球における匂い経験に依存的な新生した顆粒細胞の選別の“2段階モデル”を提唱した(図2).2段階とは嗅覚行動時とそののちの休眠時という連続して起こる2つの異なる行動様式の組合せをさす.食餌行動のような嗅覚行動中に,新生した顆粒細胞のうちのあるものは匂い入力により活性化し“タグづけ”が行われる一方,あるものは活性化されずにタグづけが行われず,これにより新生した顆粒細胞の色分けがなされる.しかし,この時点ではタグづけされていない細胞はまだ排除されない.そして,のちの休眠時に,新生した顆粒細胞に仮想的な“再編シグナル”が入力される.このシグナルはタグづけされた顆粒細胞を生存させる一方,タグづけされていない顆粒細胞を細胞死により排除する.この2段階により嗅球における匂い入力に応じた新生した顆粒細胞の利用および整理が行われ,これは神経回路における適切な再編および可塑的な変化に貢献しているものと考えられる.この2段階モデルには,海馬における学習記憶の2段階モデル6) や,覚醒・睡眠行動にともなうシナプス機能のアップスケーリング・ダウンスケーリングモデル7) との共通点を見い出すことができる.
おわりに
嗅球において新生した顆粒細胞の整理は一日中いつでも均等に起こっているのではなく,食後の休眠という特定の行動様式に対応して起こっていた.動物は日々新しい食べ物に出会い,それがおいしいかまずいか,体にとりよい栄養になったかどうか,など新たな学習をくり返している.このような食べ物の経験に応じ,のちの休眠時に新生した顆粒細胞を整理することで,動物は食べ物の情報を効率よく学習し記憶しているものと考えられる.
食餌行動は典型的な嗅覚行動であるが,嗅覚は天敵からの逃避,交尾相手の選択,なわばりの確保など個体の維持と種の繁栄にかかわるさまざまな行動に寄与している.筆者らは,これらの嗅覚行動のあとの休眠時間にも,嗅覚経験に応じた新生した顆粒細胞の利用および整理が促進されているのではないかと想定している.動物は嗅覚行動中には匂い情報,つまり,外界の匂い環境の情報を適切に処理することに集中しているのかもしれない.そして,のちの休眠時には外界の匂い情報の処理をいったん中断し,さきの匂い経験にもとづいて大規模な神経回路の再編を行うことに集中し,つぎに覚醒したときに,より適切な匂い行動をとれるよう備えているのではないだろうか.
最近,筆者らの研究室では,睡眠中には嗅球の上位中枢である嗅皮質から同期的なトップダウン性シナプス入力が嗅球にむかって入力することを見い出している8).この入力が嗅球において新生した顆粒細胞の整理を促進する“再編シグナル”の実体である可能性があり,現在,この点を検討中である.また,経験に依存的な“タグづけ”の機構は神経科学の全般に共通する大きな課題であり,嗅球の神経回路はその解明にむけて格好の題材を提供しているものと思われる.
文 献
- Lledo, P. M., Alonso, M. & Grubb, M. S.: Adult neurogenesis and functional plasticity in neuronal circuits. Nat. Rev. Neurosci., 7, 179-193 (2006)[PubMed]
- Mouret, A., Lepousez, G., Gras, J. et al.: Turnover of newborn olfactory bulb neurons optimizes olfaction. J. Neurosci., 29, 12302-12314 (2009)[PubMed]
- Yamaguchi, M. & Mori, K.: Critical period for sensory experience-dependent survival of newly generated granule cells in the adult mouse olfactory bulb. Proc. Natl. Acad. Sci. USA, 102, 9697-9702 (2005)[PubMed]
- Mori, K., Takahashi, Y. K., Igarashi, K. M. et al.: Maps of odorant molecular features in the mammalian olfactory bulb. Physiol. Rev., 86, 409-433 (2006)[PubMed]
- Kobayakawa, K., Kobayakawa, R., Matsumoto, H. et al.: Innate versus learned odour processing in the mouse olfactory bulb. Nature, 450, 503-508 (2007)[PubMed]
- Diekelmann, S. & Born, J.: The memory function of sleep. Nat. Rev. Neurosci., 11, 114-126 (2010)[PubMed]
- Tononi, G. & Cirelli, C.: Sleep function and synaptic homeostasis. Sleep Med. Rev., 10, 49-62 (2006)[PubMed]
- Manabe, H., Kusumoto-Yoshida, I., Ota, M. et al.: Olfactory cortex generates synchronized top-down inputs to the olfactory bulb during slow-wave sleep. J. Neurosci., 31, 8123-8133 (2011)[PubMed]
著者プロフィール
略歴:2011年 東京大学大学院医学系研究科博士課程 単位取得退学,同年より東京大学 学術支援職員.
持丸 大輔(Daisuke Mochimaru)
略歴:東京大学大学院医学系研究科博士課程 在学中.
研究テーマ:成体の嗅球における新生ニューロンの生死の決定および組み込み機構の解明.
抱負:新生ニューロンが生き残り嗅球の神経回路の一部としてはたらくようになるには何が必要なのかを解明したい.
山口 正洋(Masahiro Yamaguchi)
東京大学大学院医学系研究科 講師.
© 2011 横山 健・持丸大輔・山口正洋 Licensed under CC 表示 2.1 日本
