網膜における神経回路の形成と視覚機能を選択的に制御する膜型セマフォリンの発見
松岡 亮太
(米国California大学San Francisco校,Department of Biochemistry and Biophysics)
email:松岡亮太
DOI: 10.7875/first.author.2011.133
Class 5 transmembrane semaphorins control selective mammalian retinal lamination and function.
Ryota L. Matsuoka, Onanong Chivatakarn, Tudor C. Badea, Ivy S. Samuels, Hugh Cahill, Kei-ichi Katayama, Sumit R. Kumar, Fumikazu Suto, Alain Chédotal, Neal S. Peachey, Jeremy Nathans, Yutaka Yoshida, Roman J. Giger, Alex L. Kolodkin
Neuron, 71, 460-473 (2011)
視覚は聴覚・嗅覚・触覚・味覚とともに五感のひとつであり,われわれが外界から情報を得るうえで重要な役割をはたしている.ヒトでは視覚からの情報の割合が高く,全情報量の約80%を視覚にたよっているともいわれている.外界からの視覚情報は目の網膜に存在するニューロンにより光や色として受容され,電気シグナルに変換されて精密な神経回路をとおし脳に伝達されることにより認知されている.したがって,網膜においてニューロンが視覚情報を処理するしくみを解き明かすことが視覚を理解するうえでの鍵となる.しかしながら,網膜において光や色など異なる視覚情報を処理する神経回路が形成される分子機構は多くが不明である.筆者らは,神経回路の形成に深くかかわるセマフォリンの網膜における機能的な解析をつうじて,光強度の減少に反応する神経回路の形成に必須のタンパク質を発見した.このクラス5に分類される膜型セマフォリンをノックアウトしたマウスでは,光強度の減少に反応する神経回路の層構造と,その関連する視覚機能に異常が生じることをみつけた.これらの研究成果は,特定の視覚機能に必要不可欠なタンパク質を同定しその分子機構を見い出した点において,視覚研究における重大な発見であった.
五感の機能はその機能をつかさどるニューロンどうしが神経組織の発生段階において正しく神経回路ネットワークを形成することにより確立される.目,耳,鼻,手足などの末梢組織で受容された外界からの情報を正しく脳に伝達するのが神経回路ネットワークの役割のひとつであり,さまざまな外界刺激を電気シグナルに変換することでその情報を脳に伝達している.その際,さまざまな外界刺激に対応する手段として,神経組織は機能ごとに整然と分離して回路を組織することにより効率的な情報処理を行っているものと考えられている.その組織の様式のひとつが神経回路の層構造であり,脊髄,視床,大脳皮質など脳神経系の幅広い組織において神経回路の層構造が観察されている1-3).この神経回路の層構造と機能のあいだには密接な関係性が示唆されているが,神経回路の層構造を導く分子機構はその多くが不明である.
網膜は神経組織のなかでもとりわけ層構造の顕著な組織であり,異なる機能をもつニューロンが別々の層でシナプスを形成することにより,視覚情報に必要な神経回路が構築されていることが知られている1)(図1).光強度の増加に反応する回路(オン経路)と減少に反応する回路(オフ経路)とが視覚刺激にたくみに反応することで,物の形状や明暗など光のコントラストを認識するしくみになっている4,5).網膜ではこの異なる機能をもつ神経回路が内網状層において別々の層としてきれいに分離して存在しており,その分離が機能の確立にも重要だと考えられている1).しかしながら,その分離を導く分子機構は不明である.
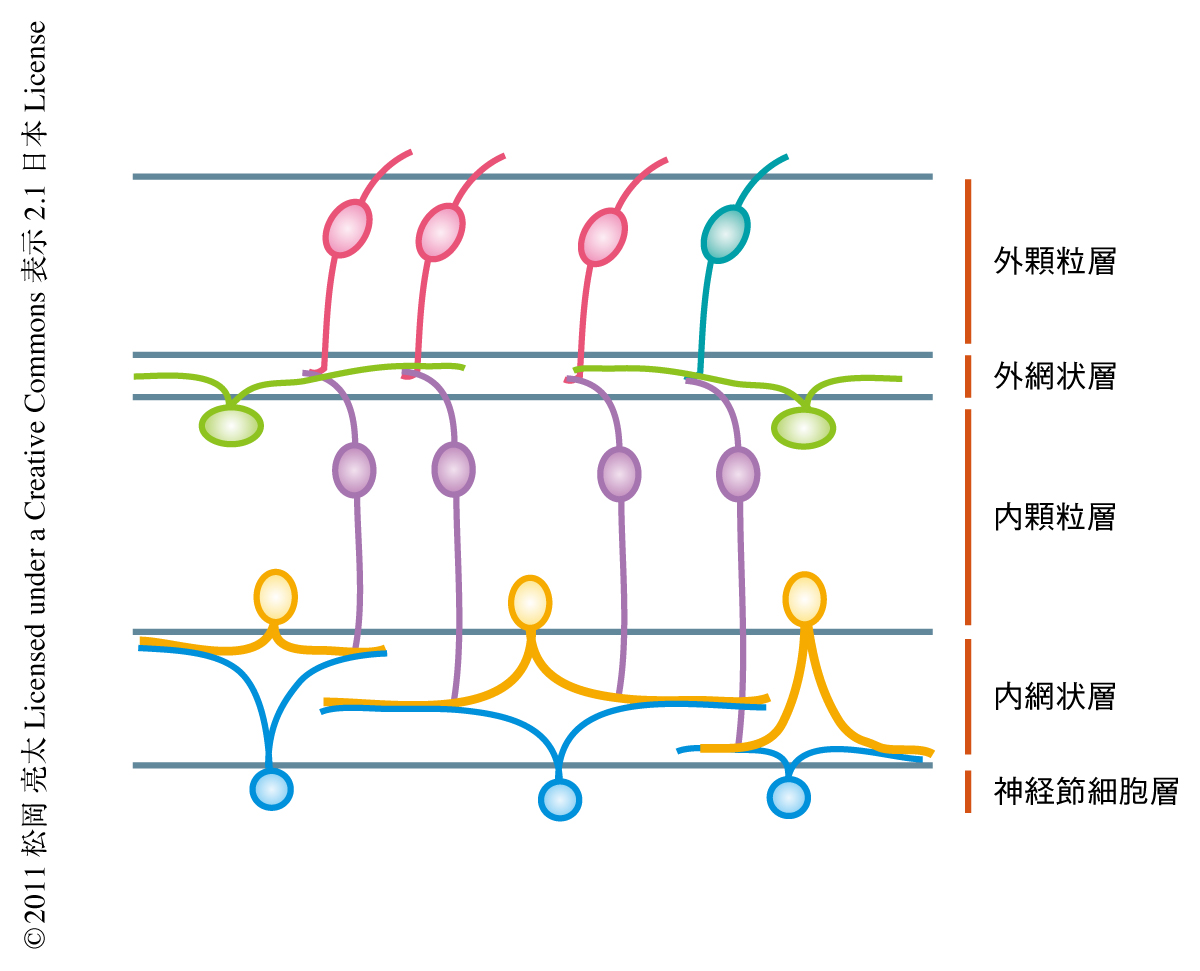
哺乳類の網膜の内網状層では,網膜神経節細胞,アマクリン細胞,双極細胞の異なるサブタイプの細胞が別々の層に神経突起を伸長することによりオン経路とオフ経路の神経回路が分離して構築されることが知られている1).筆者らは以前,神経回路の形成や維持に重要な役割をはたすセマフォリンとその受容体であるニューロピリンおよびプレキシン6,7) のmRNA発現パターンと機能的な解析をつうじて,反発ガイダンスタンパク質である膜型セマフォリンSema6Aおよびその受容体であるプレキシンPlexinA4が内網状層において特定の神経回路の層形成に重要な役割をはたしていることを発見した8)(新着論文レビュー でも掲載).
この研究では,Sema6Aとは異なる2つの膜型セマフォリン,Sema5AおよびSema5Bが網膜のオフ経路にかかわる神経回路の層形成に重要な役割をはたし,なおかつ,視覚機能の確立にも必要不可欠であることを発見した.また,これらのセマフォリンの機能を伝達するプレキシン受容体,PlexinA1およびPlexinA3も同定した.
異なる膜型セマフォリンのmRNA発現パターンの解析をつうじ,クラス5に分類される相同性の高い2つのセマフォリン,Sema5AおよびSema5Bが網膜の発生段階で類似した発現パターンを示すことをみつけた.Sema5AおよびSema5Bの網膜での役割を生体において解析するため,それぞれに対するノックアウトマウスを作製し機能解析を行った.その結果,Sema5Aノックアウトマウスの網膜においては明確な表現型を確認できなかったが,Sema5Bノックアウトマウスでは網膜の神経回路の層形成に異常がみられた.Sema5AとSema5Bはアミノ酸配列の相同性が高く機能の補完性の可能性が考えられたため,ダブルノックアウトマウスを作製し同様の解析を行ったところ,Sema5AとSema5BのダブルノックアウトマウスではSema5Bノックアウトマウスよりも網膜の神経回路の顕著な異常が確認された.興味深いことに,Sema5AとSema5Bのダブルノックアウトマウスでは,以前に筆者らが発見したSema6Aノックアウトマウスでみられたニューロンの神経突起の異常伸展とは,真逆の方向に神経突起が異常伸展していた.
哺乳類の網膜の網膜内層では,形状や機能の異なる数十種類の網膜神経節細胞,アマクリン細胞,双極細胞のサブタイプが内網状層においてシナプスを形成していることが知られている1,9).異なるサブタイプのニューロンを染色できるマーカーを用いてそれぞれのニューロンの神経突起の層投射を検討したところ,Sema5AとSema5Bのダブルノックアウトマウスにおいて,多くのサブタイプのニューロンが内顆粒層と外網状層に神経突起を異常伸展させていることを発見した.Sema5AおよびSema5BのmRNAはマウスの出生初期には外神経芽細胞層に強く発現していることから,外神経芽細胞層に発現しているSema5AおよびSema5Bが反発ガイダンスタンパク質として網膜神経節細胞,アマクリン細胞,双極細胞の神経突起を内網状層の内部に制限しているものと考えられた.これを裏づけるように,Sema5AおよびSema5Bが網膜の培養細胞による実験において神経突起の伸長を阻害することをみつけた.興味深いことに,Sema5AとSema5Bのダブルノックアウトマウスでは,内網状層の神経回路の層形成の異常とは異なり,外網状層の神経回路にはとくに異常はみつからなかった.このことから,Sema5AおよびSema5Bは網膜において内網状層の神経回路の層形成を選択的に制御していることが明らかになった.
Sema5AとSema5Bのダブルノックアウトマウスでの層構造のより詳細な解析をつうじて,内網状層で分離して存在するオン経路とオフ経路のうち,オフ経路に関連する回路がより顕著に層構造の形成異常を起こしていることを発見した.オン経路に関連するニューロンにもわずかながら異常がみられたが,正常なマウスと比較しても形態においてオフ経路ほど大きな差はみつからなかった.
網膜内層の層構造の形成異常が視覚機能と関連するかどうかを調べるため,視覚刺激に対する網膜神経節細胞の反応性を測定した.その結果,Sema5AとSema5Bのダブルノックアウトマウスでの表現型と一致するように,光強度の増加(オン経路)に応答する神経回路の反応性においては正常なマウスとSema5AとSema5Bのダブルノックアウトマウスとで大きな違いはみられなかったが,光強度の減少(オフ経路)に応答する神経回路の反応性がSema5AとSema5Bのダブルノックアウトマウスでは著しく乏しいことを発見した.このことより,Sema5AおよびSema5Bは視覚機能をつかさどる神経回路のなかでも,とりわけ光強度の減少(オフ経路)に応答する神経回路の層形成と機能に必須であることがわかった.これにくわえて,Sema5AとSema5Bのダブルノックアウトマウスでは,網膜内層の機能に関連することの知られている活動電位の減少もみつかった.しかしながら,網膜外層の機能に関連する活動電位に異常はみられなかった.これらの実験結果から,Sema5AおよびSema5Bは網膜内層の機能ならびにオフ経路の神経回路の機能を選択的に制御しているものと結論づけた.この結果は,Sema5AとSema5Bのダブルノックアウトマウスで観察された形態の表現型と完全に一致するものであった.
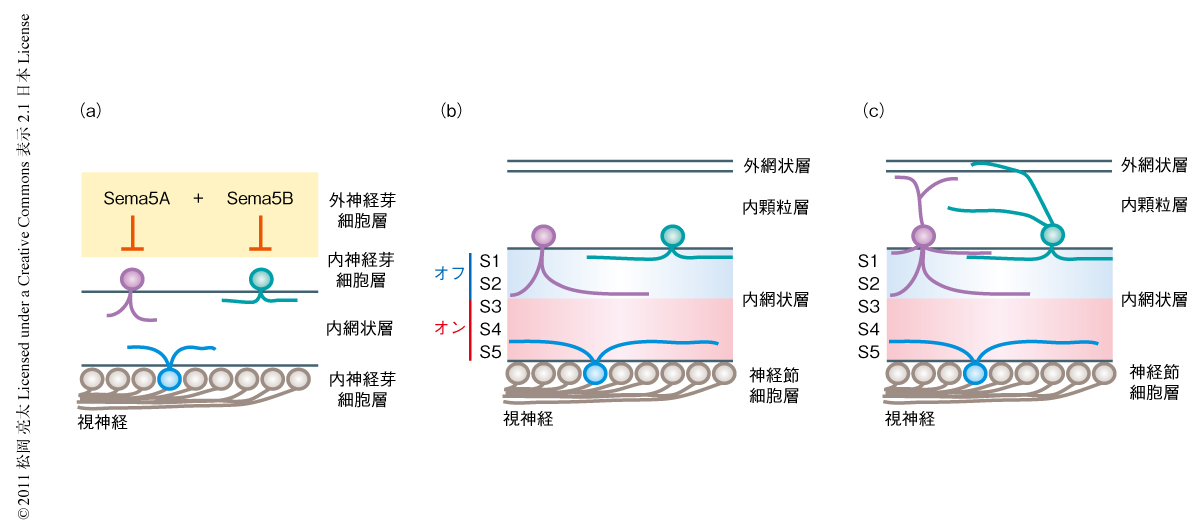
生体でのSema5AおよびSema5Bの受容体は同定されていなかったため,網膜におけるSema5AおよびSema5Bの受容体の同定を試みた.これまで,多くのセマフォリンが生体においてプレキシンおよびニューロピリンを受容体として利用することが知られているため6,7),まず,マウスに存在するすべてのプレキシン(9種類)およびニューロピリン(2種類)のノックアウトマウスを用いてその網膜を解析した8).その結果,ひとつひとつのプレキシンとニューロピリンをノックアウトしたマウスでは,Sema5AとSema5Bのダブルノックアウトマウスにおいて観察された層構造の異常と同様の表現型は確認できなかった.このことから,受容体にも機能的な補完性があると考えて,それぞれのプレキシンとニューロピリンのmRNA発現パターンをより詳細に調べたところ,PlexinA1およびPlexinA3が網膜内層において類似した発現パターンを示すことを観察した.このことから,PlexinA1とPlexinA3のダブルノックアウトマウスを作製し網膜を解析したところ,Sema5AとSema5Bのダブルノックアウトマウスでみられたのと同様の異常を発見した.また,Sema5AおよびSema5Bによる網膜の培養細胞に対する神経突起の伸長阻害も,PlexinA1とPlexinA3のダブルノックアウトマウスから得たニューロンを使用したところ観察できなくなった.このことから,PlexinA1とPlexinA3とが協同して,Sema5AとSema5Bの反発ガイダンスタンパク質としての機能を網膜において仲介しているものと結論づけた(図2).
この研究をつうじて,クラス5に属する膜型セマフォリンとその受容体となるプレキシンが網膜内層の層構造の形成に重要な役割をはたしていること,オフ経路の神経回路の層形成と機能とを選択的に制御していることが示された.これまで,脊椎動物の網膜の整然とした層構造と神経回路が構築される分子機構はその多くが謎であったが,近年の細胞接着分子による制御の発見にくわえて10,11),筆者らの研究により,膜型セマフォリンによる反発ガイダンスシグナルが網膜の神経回路の層構造の構築に重要であることがわかってきた.また,この研究により網膜の神経回路の層構造と視覚機能とが深く関係していることが明らかになってきており,神経回路の根本的な組織ユニットとしての層構造の確立における分子機構の解明が,ほかの神経組織においてもよりいっそう重要になってくることであろう.その際にこの研究での知見が役立つことを期待したい.
略歴:2011年 米国Johns Hopkins大学大学院博士課程 修了,同年より米国California大学San Francisco校 博士研究員.
研究テーマ:神経回路と脈管網の形成における分子機構の解明.
抱負:マウスとゼブラフィッシュを駆使して,発生研究の分野で質の高い仕事を継続して発表できる研究者をめざして努力したい.まずは,ゼブラフィッシュを使う新しい研究テーマに全力で挑みます.
© 2011 松岡 亮太 Licensed under CC 表示 2.1 日本
(米国California大学San Francisco校,Department of Biochemistry and Biophysics)
email:松岡亮太
DOI: 10.7875/first.author.2011.133
Class 5 transmembrane semaphorins control selective mammalian retinal lamination and function.
Ryota L. Matsuoka, Onanong Chivatakarn, Tudor C. Badea, Ivy S. Samuels, Hugh Cahill, Kei-ichi Katayama, Sumit R. Kumar, Fumikazu Suto, Alain Chédotal, Neal S. Peachey, Jeremy Nathans, Yutaka Yoshida, Roman J. Giger, Alex L. Kolodkin
Neuron, 71, 460-473 (2011)
要 約
視覚は聴覚・嗅覚・触覚・味覚とともに五感のひとつであり,われわれが外界から情報を得るうえで重要な役割をはたしている.ヒトでは視覚からの情報の割合が高く,全情報量の約80%を視覚にたよっているともいわれている.外界からの視覚情報は目の網膜に存在するニューロンにより光や色として受容され,電気シグナルに変換されて精密な神経回路をとおし脳に伝達されることにより認知されている.したがって,網膜においてニューロンが視覚情報を処理するしくみを解き明かすことが視覚を理解するうえでの鍵となる.しかしながら,網膜において光や色など異なる視覚情報を処理する神経回路が形成される分子機構は多くが不明である.筆者らは,神経回路の形成に深くかかわるセマフォリンの網膜における機能的な解析をつうじて,光強度の減少に反応する神経回路の形成に必須のタンパク質を発見した.このクラス5に分類される膜型セマフォリンをノックアウトしたマウスでは,光強度の減少に反応する神経回路の層構造と,その関連する視覚機能に異常が生じることをみつけた.これらの研究成果は,特定の視覚機能に必要不可欠なタンパク質を同定しその分子機構を見い出した点において,視覚研究における重大な発見であった.
はじめに
五感の機能はその機能をつかさどるニューロンどうしが神経組織の発生段階において正しく神経回路ネットワークを形成することにより確立される.目,耳,鼻,手足などの末梢組織で受容された外界からの情報を正しく脳に伝達するのが神経回路ネットワークの役割のひとつであり,さまざまな外界刺激を電気シグナルに変換することでその情報を脳に伝達している.その際,さまざまな外界刺激に対応する手段として,神経組織は機能ごとに整然と分離して回路を組織することにより効率的な情報処理を行っているものと考えられている.その組織の様式のひとつが神経回路の層構造であり,脊髄,視床,大脳皮質など脳神経系の幅広い組織において神経回路の層構造が観察されている1-3).この神経回路の層構造と機能のあいだには密接な関係性が示唆されているが,神経回路の層構造を導く分子機構はその多くが不明である.
網膜は神経組織のなかでもとりわけ層構造の顕著な組織であり,異なる機能をもつニューロンが別々の層でシナプスを形成することにより,視覚情報に必要な神経回路が構築されていることが知られている1)(図1).光強度の増加に反応する回路(オン経路)と減少に反応する回路(オフ経路)とが視覚刺激にたくみに反応することで,物の形状や明暗など光のコントラストを認識するしくみになっている4,5).網膜ではこの異なる機能をもつ神経回路が内網状層において別々の層としてきれいに分離して存在しており,その分離が機能の確立にも重要だと考えられている1).しかしながら,その分離を導く分子機構は不明である.
哺乳類の網膜の内網状層では,網膜神経節細胞,アマクリン細胞,双極細胞の異なるサブタイプの細胞が別々の層に神経突起を伸長することによりオン経路とオフ経路の神経回路が分離して構築されることが知られている1).筆者らは以前,神経回路の形成や維持に重要な役割をはたすセマフォリンとその受容体であるニューロピリンおよびプレキシン6,7) のmRNA発現パターンと機能的な解析をつうじて,反発ガイダンスタンパク質である膜型セマフォリンSema6Aおよびその受容体であるプレキシンPlexinA4が内網状層において特定の神経回路の層形成に重要な役割をはたしていることを発見した8)(新着論文レビュー でも掲載).
この研究では,Sema6Aとは異なる2つの膜型セマフォリン,Sema5AおよびSema5Bが網膜のオフ経路にかかわる神経回路の層形成に重要な役割をはたし,なおかつ,視覚機能の確立にも必要不可欠であることを発見した.また,これらのセマフォリンの機能を伝達するプレキシン受容体,PlexinA1およびPlexinA3も同定した.
1.クラス5セマフォリンは網膜の発生段階で発現し網膜内層の神経回路の層形成に必須である
異なる膜型セマフォリンのmRNA発現パターンの解析をつうじ,クラス5に分類される相同性の高い2つのセマフォリン,Sema5AおよびSema5Bが網膜の発生段階で類似した発現パターンを示すことをみつけた.Sema5AおよびSema5Bの網膜での役割を生体において解析するため,それぞれに対するノックアウトマウスを作製し機能解析を行った.その結果,Sema5Aノックアウトマウスの網膜においては明確な表現型を確認できなかったが,Sema5Bノックアウトマウスでは網膜の神経回路の層形成に異常がみられた.Sema5AとSema5Bはアミノ酸配列の相同性が高く機能の補完性の可能性が考えられたため,ダブルノックアウトマウスを作製し同様の解析を行ったところ,Sema5AとSema5BのダブルノックアウトマウスではSema5Bノックアウトマウスよりも網膜の神経回路の顕著な異常が確認された.興味深いことに,Sema5AとSema5Bのダブルノックアウトマウスでは,以前に筆者らが発見したSema6Aノックアウトマウスでみられたニューロンの神経突起の異常伸展とは,真逆の方向に神経突起が異常伸展していた.
2.クラス5セマフォリンは網膜内層に存在する多様なニューロンの神経突起を内網状層に制限させる
哺乳類の網膜の網膜内層では,形状や機能の異なる数十種類の網膜神経節細胞,アマクリン細胞,双極細胞のサブタイプが内網状層においてシナプスを形成していることが知られている1,9).異なるサブタイプのニューロンを染色できるマーカーを用いてそれぞれのニューロンの神経突起の層投射を検討したところ,Sema5AとSema5Bのダブルノックアウトマウスにおいて,多くのサブタイプのニューロンが内顆粒層と外網状層に神経突起を異常伸展させていることを発見した.Sema5AおよびSema5BのmRNAはマウスの出生初期には外神経芽細胞層に強く発現していることから,外神経芽細胞層に発現しているSema5AおよびSema5Bが反発ガイダンスタンパク質として網膜神経節細胞,アマクリン細胞,双極細胞の神経突起を内網状層の内部に制限しているものと考えられた.これを裏づけるように,Sema5AおよびSema5Bが網膜の培養細胞による実験において神経突起の伸長を阻害することをみつけた.興味深いことに,Sema5AとSema5Bのダブルノックアウトマウスでは,内網状層の神経回路の層形成の異常とは異なり,外網状層の神経回路にはとくに異常はみつからなかった.このことから,Sema5AおよびSema5Bは網膜において内網状層の神経回路の層形成を選択的に制御していることが明らかになった.
3.クラス5セマフォリンはオフ経路にかかわる神経回路の層形成を選択的に制御する
Sema5AとSema5Bのダブルノックアウトマウスでの層構造のより詳細な解析をつうじて,内網状層で分離して存在するオン経路とオフ経路のうち,オフ経路に関連する回路がより顕著に層構造の形成異常を起こしていることを発見した.オン経路に関連するニューロンにもわずかながら異常がみられたが,正常なマウスと比較しても形態においてオフ経路ほど大きな差はみつからなかった.
4.クラス5セマフォリンは網膜内層の視覚機能に重要かつオフ経路にかかわる神経回路の機能に必須である
網膜内層の層構造の形成異常が視覚機能と関連するかどうかを調べるため,視覚刺激に対する網膜神経節細胞の反応性を測定した.その結果,Sema5AとSema5Bのダブルノックアウトマウスでの表現型と一致するように,光強度の増加(オン経路)に応答する神経回路の反応性においては正常なマウスとSema5AとSema5Bのダブルノックアウトマウスとで大きな違いはみられなかったが,光強度の減少(オフ経路)に応答する神経回路の反応性がSema5AとSema5Bのダブルノックアウトマウスでは著しく乏しいことを発見した.このことより,Sema5AおよびSema5Bは視覚機能をつかさどる神経回路のなかでも,とりわけ光強度の減少(オフ経路)に応答する神経回路の層形成と機能に必須であることがわかった.これにくわえて,Sema5AとSema5Bのダブルノックアウトマウスでは,網膜内層の機能に関連することの知られている活動電位の減少もみつかった.しかしながら,網膜外層の機能に関連する活動電位に異常はみられなかった.これらの実験結果から,Sema5AおよびSema5Bは網膜内層の機能ならびにオフ経路の神経回路の機能を選択的に制御しているものと結論づけた.この結果は,Sema5AとSema5Bのダブルノックアウトマウスで観察された形態の表現型と完全に一致するものであった.
5.PlexinA1とPlexinA3は協同してSema5AとSema5Bのシグナルを伝達し網膜の層形成にも必須である
生体でのSema5AおよびSema5Bの受容体は同定されていなかったため,網膜におけるSema5AおよびSema5Bの受容体の同定を試みた.これまで,多くのセマフォリンが生体においてプレキシンおよびニューロピリンを受容体として利用することが知られているため6,7),まず,マウスに存在するすべてのプレキシン(9種類)およびニューロピリン(2種類)のノックアウトマウスを用いてその網膜を解析した8).その結果,ひとつひとつのプレキシンとニューロピリンをノックアウトしたマウスでは,Sema5AとSema5Bのダブルノックアウトマウスにおいて観察された層構造の異常と同様の表現型は確認できなかった.このことから,受容体にも機能的な補完性があると考えて,それぞれのプレキシンとニューロピリンのmRNA発現パターンをより詳細に調べたところ,PlexinA1およびPlexinA3が網膜内層において類似した発現パターンを示すことを観察した.このことから,PlexinA1とPlexinA3のダブルノックアウトマウスを作製し網膜を解析したところ,Sema5AとSema5Bのダブルノックアウトマウスでみられたのと同様の異常を発見した.また,Sema5AおよびSema5Bによる網膜の培養細胞に対する神経突起の伸長阻害も,PlexinA1とPlexinA3のダブルノックアウトマウスから得たニューロンを使用したところ観察できなくなった.このことから,PlexinA1とPlexinA3とが協同して,Sema5AとSema5Bの反発ガイダンスタンパク質としての機能を網膜において仲介しているものと結論づけた(図2).
おわりに
この研究をつうじて,クラス5に属する膜型セマフォリンとその受容体となるプレキシンが網膜内層の層構造の形成に重要な役割をはたしていること,オフ経路の神経回路の層形成と機能とを選択的に制御していることが示された.これまで,脊椎動物の網膜の整然とした層構造と神経回路が構築される分子機構はその多くが謎であったが,近年の細胞接着分子による制御の発見にくわえて10,11),筆者らの研究により,膜型セマフォリンによる反発ガイダンスシグナルが網膜の神経回路の層構造の構築に重要であることがわかってきた.また,この研究により網膜の神経回路の層構造と視覚機能とが深く関係していることが明らかになってきており,神経回路の根本的な組織ユニットとしての層構造の確立における分子機構の解明が,ほかの神経組織においてもよりいっそう重要になってくることであろう.その際にこの研究での知見が役立つことを期待したい.
文 献
- Sanes, J. R. & Zipursky, S. L.: Design principles of insect and vertebrate visual systems. Neuron, 66, 15-36 (2010)[PubMed]
- Molyneaux, B. J., Arlotta, P., Menezes, J. R. et al.: Neuronal subtype specification in the cerebral cortex. Nat. Rev. Neurosci., 8, 427-437 (2007)[PubMed]
- Ozaki, S. & Snider, W. D.: Initial trajectories of sensory axons toward laminar targets in the developing mouse spinal cord. J. Comp. Neurol., 380, 215-229 (1997)[PubMed]
- Wassle, H.: Parallel processing in the mammalian retina. Nat. Rev. Neurosci., 5, 747-757 (2004)[PubMed]
- Famiglietti, E. V., Jr. & Kolb, H.: Structural basis for ON-and OFF-center responses in retinal ganglion cells. Science, 194, 193-195 (1976)[PubMed]
- Yazdani, U. & Terman, J. R.: The semaphorins. Genome Biol., 7, 211 (2006)[PubMed]
- Tran, T. S., Kolodkin, A. L. & Bharadwaj, R.: Semaphorin regulation of cellular morphology. Annu. Rev. Cell Dev. Biol., 23, 263-292 (2007)[PubMed]
- Matsuoka, R. L., Nguyen-Ba-Charvet, K. T., Parray, A. et al.: Transmembrane semaphorin signalling controls laminar stratification in the mammalian retina. Nature, 470, 259-263 (2011)[PubMed] [新着論文レビュー]
- Huberman, A. D., Clandinin, T. R. & Baier, H.: Molecular and cellular mechanisms of lamina-specific axon targeting. Cold Spring Harb. Perspect. Biol., 2, a001743 (2010)[PubMed]
- Yamagata, M. & Sanes, J. R.: Dscam and Sidekick proteins direct lamina-specific synaptic connections in vertebrate retina. Nature, 451, 465-469 (2008)[PubMed]
- Yamagata, M., Weiner, J. A. & Sanes, J. R.: Sidekicks: synaptic adhesion molecules that promote lamina-specific connectivity in the retina. Cell, 110, 649-660 (2002)[PubMed]
著者プロフィール
略歴:2011年 米国Johns Hopkins大学大学院博士課程 修了,同年より米国California大学San Francisco校 博士研究員.
研究テーマ:神経回路と脈管網の形成における分子機構の解明.
抱負:マウスとゼブラフィッシュを駆使して,発生研究の分野で質の高い仕事を継続して発表できる研究者をめざして努力したい.まずは,ゼブラフィッシュを使う新しい研究テーマに全力で挑みます.
© 2011 松岡 亮太 Licensed under CC 表示 2.1 日本
