海馬の神経回路は神経活動に依存的な競合により精緻化される
安田昌弘・梅森久視
(米国Michigan大学Medical School,Molecular & Behavioral Neuroscience Institute)
email:安田昌弘,梅森久視
DOI: 10.7875/first.author.2011.122
Multiple forms of activity-dependent competition refine hippocampal circuits in vivo.
Masahiro Yasuda, Erin M. Johnson-Venkatesh, Helen Zhang, Jack M. Parent, Michael A. Sutton, Hisashi Umemori
Neuron, 70, 1128-1142 (2011)
記憶の形成には海馬での機能的な神経回路の形成が必要である.効率的な神経回路の形成においては,神経活動に依存的なシナプス結合の精緻化が起こるものと考えられている.しかしながら,生体における海馬での神経回路の形成がどのように神経活動に依存的に精緻化されているのかはこれまで調べられていなかった.筆者らは,マウスの海馬において限局された領域のニューロンを遺伝子工学的に不活性化する系を開発し,海馬での神経回路の形成過程における神経活動の影響を調べ,その結果,不活性な軸索は活性な軸索との競合により排除されることを示した.興味深いことに,新しいニューロンが一生をつうじ産生される歯状回では,シナプス結合の精緻化は新生ニューロンと成熟ニューロンとのあいだの競合により達成されることが示された.これらの結果より,活性な軸索と不活性な軸索とのシナプス結合の競合が機能的な記憶回路の形成に重要な役割をはたしていることが明らかになった.
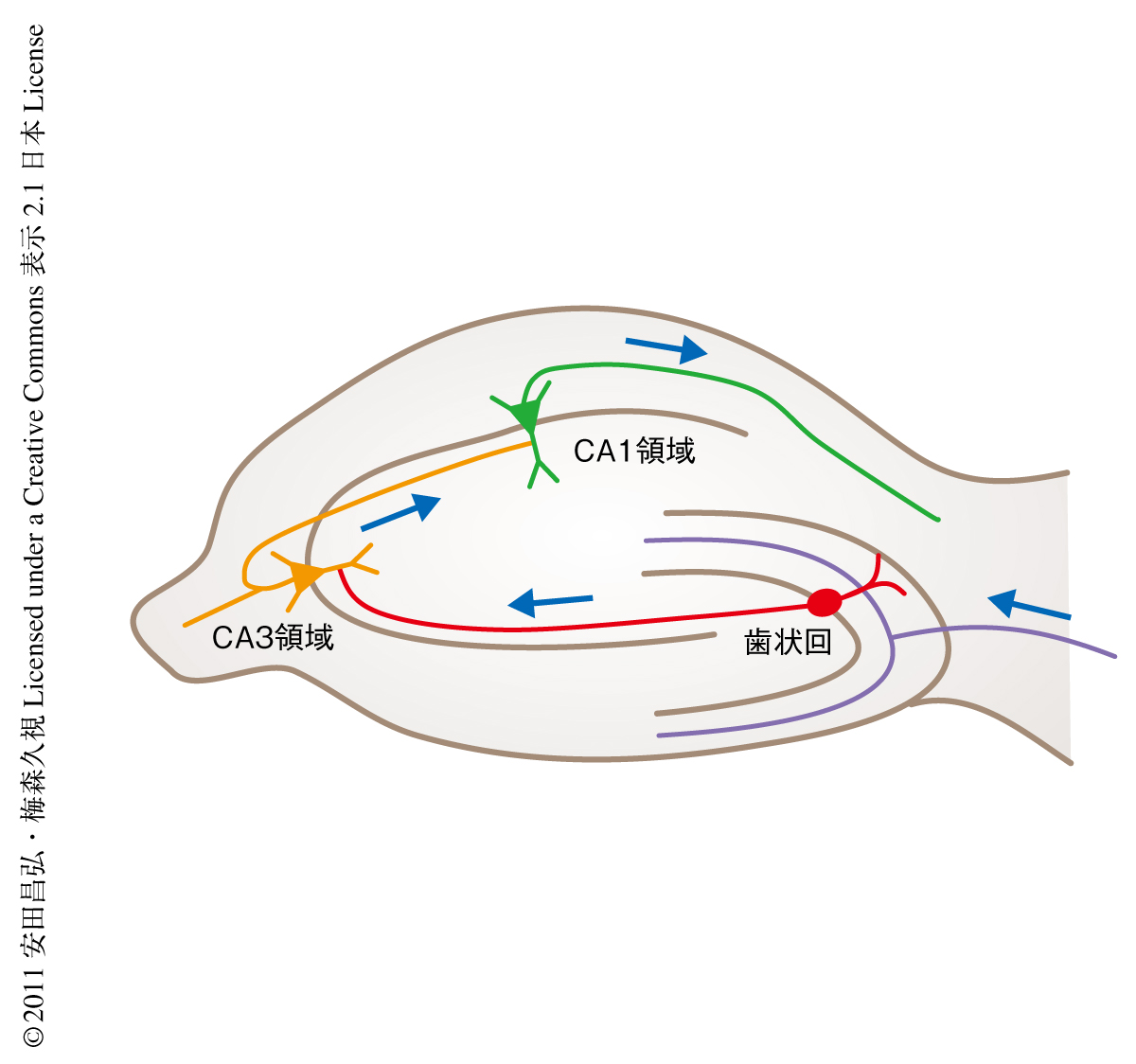
海馬は記憶の形成およびその保持に重要な役割をはたしていることが知られている1,2).海馬の神経回路は“三シナプス回路”(trisynaptic pathway)とよばれる,嗅内皮質のニューロンよりはじまり,歯状回,CA1領域,CA3領域をへて,再び嗅内皮質へともどる高度に組織化された一方向的なシナプス結合により形成されている2,3)(図1).最近の領域特異的な遺伝子改変系の導入などにより,この一方向的なシナプス結合のそれぞれの活動が記憶の形成あるいは保持にかかわっていることが明らかにされてきている.
このような正しいシナプス結合は,分泌タンパク質や接着分子などガイダンスタンパク質によるプログラミングされた神経回路の形成が行われたのち,シナプス結合が神経活動に依存的に精緻化されて形成されるものと考えられている4,5).神経活動に依存的なシナプス結合の精緻化は神経-筋結合部,感覚神経系,運動神経系では重要な役割をはたすことが知られており,いずれの場合も発生の初期においては標的となるニューロンは複数のニューロンからの入力をうけているが,発生が進むにしたがいシナプス結合が神経活動に依存的に精緻化され,最終的には少数のより活性なニューロンからの入力のみが維持される4,5).
これまで,おもに技術的な問題から,海馬を含む記憶系における神経活動に依存的なシナプス結合の精緻化については調べられていなかったが,筆者らは,領域特異的にニューロンの活動を制御できるトランスジェニックマウスを作製し,海馬での神経回路の形成における神経活動に依存的なシナプス結合の精緻化を解析した.
海馬での三シナプス回路の形成における神経活動の役割を解析するにあたり,テトラサイクリントランスアクチベーターを領域特異的に発現するトランスジェニックマウス6) と,このテトラサイクリントランスアクチベーターの制御のもと破傷風菌毒素を発現するトランスジェニックマウス7) とを掛け合わせ,領域特異的にニューロンを不活性化できるトランスジェニックマウスを作製した.
まず,嗅内皮質と歯状回とのあいだの神経活動が神経回路の形成に及ぼす影響を,嗅内皮質のニューロンを特異的に不活性化するトランスジェニックマウスを用いて検討した.対照マウスにおいてシナプス結合の形成を調べると,活性をもつ正常な軸索は生後12~16日までに標的の領域である歯状回の分子層に到達し,そののちも維持されていた.ところが,嗅内皮質のニューロンの約40%を不活性化させたトランスジェニックマウスでは,不活性な軸索は歯状回に到達したのち生後12日以降に著しく減少し,生後18日にはほとんど検出されなくなった.このあいだ,嗅内皮質においてニューロン死は確認されなかった.この結果は,嗅内皮質において不活性なニューロンの軸索は,初期の標的の領域への投射は正常であるが,生後12日以降に標的の領域より排除されることを示した.
嗅内皮質において不活性なニューロンの軸索が神経回路の形成過程で排除されることが示されたが,これは不活性なニューロンと活性なニューロンとのあいだの神経活動に依存的な競合により不活性なニューロンが排除された可能性が考えられた.ここで用いたトランスジェニックマウスでは,嗅内皮質において約40%のニューロンが破傷風菌毒素を発現することで不活性な状態になっており,残り約60%のニューロンは活性な状態であると考えられた.もし,嗅内皮質の神経回路すべてにおいてニューロンの神経活動を抑制すれば,ニューロンどうしの神経活動に依存的な競合を抑制することができる.そこで,生後9日からフグ毒素テトロドトキシンを海馬に投与しその全体の神経活動を抑制した.すると,破傷風菌毒素の発現により不活性な状態となったニューロンの軸索の排除は,テトロドトキシン非投与群と比較して著しく阻害された.この結果より,破傷風菌毒素を発現することで不活性な状態となったニューロンの軸索の排除は神経活動に依存的な競合の結果であることが示された.
海馬での領域により活動に依存的な競合の様式には違いがあるのか,歯状回とCA3領域とのあいだのシナプス結合について検討を行った.この研究のため,歯状回において約半分のニューロンが不活性化したトランスジェニックマウスと,ほとんどのニューロンが不活性化したトランスジェニックマウスの2系統のトランスジェニックマウスを作製した.半分のニューロンが不活性化したトランスジェニックマウスの場合は競合的な状況,ほとんどのニューロンが不活性化したトランスジェニックマウスの場合は非競合的な状況を遺伝子工学的につくりだせたことになる.生後15日のマウスにおいて電気生理学的に興奮性シナプス後電位を測定すると,これらのトランスジェニックマウスでは対照のマウスに対しそれぞれ44%および80%の減少が確認された.この値は,歯状回において不活性なニューロンの割合をほぼ反映していた.
歯状回において半分のニューロンを不活性化したトランスジェニックマウス(競合的な状況)では,不活性な軸索は生後12日にはその標的の領域であるCA3領域の透明層に到達していた.このことは,歯状回とCA3領域とのあいだの神経回路の形成においては,嗅内皮質と歯状回とのあいだと同様に,初期の軸索投射はニューロンが不活性な状態でもその影響をうけないことを示していた.しかし,生後20日においては不活性な軸索は減少し,生後25日以降にはほとんど検出されなくなった.これに対し,歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウス(非競合的な状況)では,不活性な軸索は排除されないものと予想された.しかし実際には,このトランスジェニックマウスでも不活性な軸索は生後15日~25日のあいだに排除された.対照となるマウスではこのような軸索の排除は生じないことからこれは発生過程における軸索の排除ではなく,ニューロンの不活性化が軸索の排除を促しているものと考えられた.このトランスジェニックマウスにおける不活性な軸索の排除の分子機構を明らかにするため,共焦点顕微鏡で詳細を観察してみると,生後20日において不活性な軸索の末端部には膨張した軸索あるいは軸索の断片が確認された.これは神経-筋結合部におけるシナプス結合の精緻化のときと類似していた8).
歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウスでの不活性な軸索の排除は,ニューロンの神経活動には非依存的,あるいは,歯状回の成熟ニューロン以外のニューロンとの神経活動に依存的な競合,という2つの可能性が考えられた.この2つを区別するため,テトロドトキシンを海馬に投与しその全体の神経活動を抑制した.すると,破傷風菌毒素の発現により不活性な状態となったニューロンの軸索の排除は,テトロドトキシン非投与群に比較して著しく阻害された.このことより,歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウスにおける軸索の排除は,神経活動に依存的な競合によるものであることがわかった.
それでは,歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウスでは,不活性なニューロンはどのニューロンと競合しているのだろうか? 可能性のひとつとして,歯状回では新しいニューロンが一生をつうじて産生されることから9),この新生ニューロンが不活性な成熟ニューロンと神経活動に依存的に競合していることが考えられた.新生ニューロンに破傷風菌毒素が発現しているかどうか新生ニューロンのマーカーを使い調べてみたところ発現していなかったことから,この可能性が支持された.その証明のため,歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウスを細胞分裂の阻害剤であるAraCにより処理することで新生ニューロンの産生を抑制し,不活性な軸索の排除への影響を調べてみた.すると,新生ニューロンの産生を抑制すると不活性な軸索の排除は著しく抑制されることがわかった.この発見を補強するため別の細胞分裂の阻害剤であるTemozolomideによる処理も行ったが,やはり同様の結果が得られた.
これらの結果から,歯状回において精緻化の起こる期間に新生したニューロンは,成熟ニューロンと効率的に競合しシナプス結合の精緻化にかかわるものと考えられた.そうであるなら,歯状回の新生ニューロンはすみやかにCA3領域のニューロンとシナプスを形成するだろう.これを証明するため,生後15日において歯状回の新生ニューロンにGFPを発現するレトロウイルスを感染させてこれを標識し,生後23日においてGFPを発現する軸索とシナプスの形態を観察した.その結果,生後15日に新生した歯状回のニューロンの軸索は,生後23日にはすでにCA3領域において標的の領域の末端にまで到達しており,歯状回の成熟ニューロンの軸索に形成されたシナプスと同等かそれ以上のシナプスが形成されていた.これらのシナプスは興奮性シナプス小胞や機能的前シナプスのマーカーにより染色されることから,すでに機能しているものと考えられた.
非競合的な状況である歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウスと同様に,競合的な状況である歯状回において半分のニューロンが不活性化したトランスジェニックマウスでも,新生ニューロンの産生を抑制することで不活性な軸索の排除の抑制が観察されるのかどうか調べてみた.歯状回において成熟ニューロンどうしの競合があるなら,細胞分裂の阻害剤であるAraCにより処理することでニューロンの新生を抑えた場合でも不活性な軸索は排除されるはずである.ところがこの場合,不活性なニューロンの軸索の排除は抑制された.つまり,歯状回における不活性なニューロンの軸索の排除は,おもに不活性な成熟ニューロンと新生ニューロンとの競合の結果であり,成熟ニューロンどうしの競合によるものではないことが示唆された.
薬剤による処理では副作用の影響やニューロン新生の抑制の程度が問題になりうるため,遺伝学的にニューロン新生を抑制して神経活動の依存的な競合が成熟ニューロンと新生ニューロンのあいだで起こっているのかどうかを検討した.選択的かつ効率的に神経前駆細胞を死滅させることのできるチミジンキナーゼを発現するトランスジェニックマウスと,歯状回において半分のニューロンが不活性化したトランスジェニックマウスとを掛け合わせて実験に用いた10).その結果,このトランスジェニックマウスでは不活性な軸索はほとんど排除されなかった.この結果は,歯状回とCA3領域とのあいだでの神経活動に依存的な競合は成熟ニューロンと新生ニューロンとのあいだで起こり,成熟ニューロンどうしのあいだではほとんど起こらないことを証明した.また,チミジンキナーゼの発現はニューロン新生の起こらないCA1領域では不活性な軸索の排除に影響を及ぼさなかった.
この研究において,筆者らは,神経活動に依存的な競合が海馬での神経回路の形成におけるシナプス結合の精緻化において一般的な原理であることをin vivoではじめて証明した.特筆すべきは,新しいニューロンがたえず産生される歯状回では,発生過程において成熟ニューロンが新生ニューロンと神経活動に依存的に競合することが示されたことである.この結果は,新生ニューロンがすでに形成され確立された成体の神経回路に,どのようにして効率よく組み込まれるのかを説明しているのかもしれない11,12).最近の研究では,歯状回におけるニューロンの新生は記憶の形成に重要であることが示されている.筆者らが発見した歯状回における神経活動に依存的な競合は,認知機能に重要と考えられるすみやかで効率的な新生ニューロンの組み込みの分子機構にかかわっているのかもしれない.
略歴:総合研究大学院大学,米国Scripps Resarch Instituteを経て,米国Michigan大学Medical School博士研究員.
梅森 久視(Hisashi Umemori)
米国Michigan大学Medical SchoolにてAssistant Professor.
© 2011 安田昌弘・梅森久視 Licensed under CC 表示 2.1 日本
(米国Michigan大学Medical School,Molecular & Behavioral Neuroscience Institute)
email:安田昌弘,梅森久視
DOI: 10.7875/first.author.2011.122
Multiple forms of activity-dependent competition refine hippocampal circuits in vivo.
Masahiro Yasuda, Erin M. Johnson-Venkatesh, Helen Zhang, Jack M. Parent, Michael A. Sutton, Hisashi Umemori
Neuron, 70, 1128-1142 (2011)
要 約
記憶の形成には海馬での機能的な神経回路の形成が必要である.効率的な神経回路の形成においては,神経活動に依存的なシナプス結合の精緻化が起こるものと考えられている.しかしながら,生体における海馬での神経回路の形成がどのように神経活動に依存的に精緻化されているのかはこれまで調べられていなかった.筆者らは,マウスの海馬において限局された領域のニューロンを遺伝子工学的に不活性化する系を開発し,海馬での神経回路の形成過程における神経活動の影響を調べ,その結果,不活性な軸索は活性な軸索との競合により排除されることを示した.興味深いことに,新しいニューロンが一生をつうじ産生される歯状回では,シナプス結合の精緻化は新生ニューロンと成熟ニューロンとのあいだの競合により達成されることが示された.これらの結果より,活性な軸索と不活性な軸索とのシナプス結合の競合が機能的な記憶回路の形成に重要な役割をはたしていることが明らかになった.
はじめに
海馬は記憶の形成およびその保持に重要な役割をはたしていることが知られている1,2).海馬の神経回路は“三シナプス回路”(trisynaptic pathway)とよばれる,嗅内皮質のニューロンよりはじまり,歯状回,CA1領域,CA3領域をへて,再び嗅内皮質へともどる高度に組織化された一方向的なシナプス結合により形成されている2,3)(図1).最近の領域特異的な遺伝子改変系の導入などにより,この一方向的なシナプス結合のそれぞれの活動が記憶の形成あるいは保持にかかわっていることが明らかにされてきている.
このような正しいシナプス結合は,分泌タンパク質や接着分子などガイダンスタンパク質によるプログラミングされた神経回路の形成が行われたのち,シナプス結合が神経活動に依存的に精緻化されて形成されるものと考えられている4,5).神経活動に依存的なシナプス結合の精緻化は神経-筋結合部,感覚神経系,運動神経系では重要な役割をはたすことが知られており,いずれの場合も発生の初期においては標的となるニューロンは複数のニューロンからの入力をうけているが,発生が進むにしたがいシナプス結合が神経活動に依存的に精緻化され,最終的には少数のより活性なニューロンからの入力のみが維持される4,5).
これまで,おもに技術的な問題から,海馬を含む記憶系における神経活動に依存的なシナプス結合の精緻化については調べられていなかったが,筆者らは,領域特異的にニューロンの活動を制御できるトランスジェニックマウスを作製し,海馬での神経回路の形成における神経活動に依存的なシナプス結合の精緻化を解析した.
1.領域特異的にニューロンの活動を制御できるトランスジェニックマウスの作製
海馬での三シナプス回路の形成における神経活動の役割を解析するにあたり,テトラサイクリントランスアクチベーターを領域特異的に発現するトランスジェニックマウス6) と,このテトラサイクリントランスアクチベーターの制御のもと破傷風菌毒素を発現するトランスジェニックマウス7) とを掛け合わせ,領域特異的にニューロンを不活性化できるトランスジェニックマウスを作製した.
2.嗅内皮質において不活性なニューロンの軸索は標的の領域に到達したのち排除される
まず,嗅内皮質と歯状回とのあいだの神経活動が神経回路の形成に及ぼす影響を,嗅内皮質のニューロンを特異的に不活性化するトランスジェニックマウスを用いて検討した.対照マウスにおいてシナプス結合の形成を調べると,活性をもつ正常な軸索は生後12~16日までに標的の領域である歯状回の分子層に到達し,そののちも維持されていた.ところが,嗅内皮質のニューロンの約40%を不活性化させたトランスジェニックマウスでは,不活性な軸索は歯状回に到達したのち生後12日以降に著しく減少し,生後18日にはほとんど検出されなくなった.このあいだ,嗅内皮質においてニューロン死は確認されなかった.この結果は,嗅内皮質において不活性なニューロンの軸索は,初期の標的の領域への投射は正常であるが,生後12日以降に標的の領域より排除されることを示した.
3.嗅内皮質における不活性な軸索の排除は活性な軸索との競合の結果である
嗅内皮質において不活性なニューロンの軸索が神経回路の形成過程で排除されることが示されたが,これは不活性なニューロンと活性なニューロンとのあいだの神経活動に依存的な競合により不活性なニューロンが排除された可能性が考えられた.ここで用いたトランスジェニックマウスでは,嗅内皮質において約40%のニューロンが破傷風菌毒素を発現することで不活性な状態になっており,残り約60%のニューロンは活性な状態であると考えられた.もし,嗅内皮質の神経回路すべてにおいてニューロンの神経活動を抑制すれば,ニューロンどうしの神経活動に依存的な競合を抑制することができる.そこで,生後9日からフグ毒素テトロドトキシンを海馬に投与しその全体の神経活動を抑制した.すると,破傷風菌毒素の発現により不活性な状態となったニューロンの軸索の排除は,テトロドトキシン非投与群と比較して著しく阻害された.この結果より,破傷風菌毒素を発現することで不活性な状態となったニューロンの軸索の排除は神経活動に依存的な競合の結果であることが示された.
4.歯状回において不活性なニューロンの軸索は標的の領域に到達したのち排除される
海馬での領域により活動に依存的な競合の様式には違いがあるのか,歯状回とCA3領域とのあいだのシナプス結合について検討を行った.この研究のため,歯状回において約半分のニューロンが不活性化したトランスジェニックマウスと,ほとんどのニューロンが不活性化したトランスジェニックマウスの2系統のトランスジェニックマウスを作製した.半分のニューロンが不活性化したトランスジェニックマウスの場合は競合的な状況,ほとんどのニューロンが不活性化したトランスジェニックマウスの場合は非競合的な状況を遺伝子工学的につくりだせたことになる.生後15日のマウスにおいて電気生理学的に興奮性シナプス後電位を測定すると,これらのトランスジェニックマウスでは対照のマウスに対しそれぞれ44%および80%の減少が確認された.この値は,歯状回において不活性なニューロンの割合をほぼ反映していた.
歯状回において半分のニューロンを不活性化したトランスジェニックマウス(競合的な状況)では,不活性な軸索は生後12日にはその標的の領域であるCA3領域の透明層に到達していた.このことは,歯状回とCA3領域とのあいだの神経回路の形成においては,嗅内皮質と歯状回とのあいだと同様に,初期の軸索投射はニューロンが不活性な状態でもその影響をうけないことを示していた.しかし,生後20日においては不活性な軸索は減少し,生後25日以降にはほとんど検出されなくなった.これに対し,歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウス(非競合的な状況)では,不活性な軸索は排除されないものと予想された.しかし実際には,このトランスジェニックマウスでも不活性な軸索は生後15日~25日のあいだに排除された.対照となるマウスではこのような軸索の排除は生じないことからこれは発生過程における軸索の排除ではなく,ニューロンの不活性化が軸索の排除を促しているものと考えられた.このトランスジェニックマウスにおける不活性な軸索の排除の分子機構を明らかにするため,共焦点顕微鏡で詳細を観察してみると,生後20日において不活性な軸索の末端部には膨張した軸索あるいは軸索の断片が確認された.これは神経-筋結合部におけるシナプス結合の精緻化のときと類似していた8).
5.歯状回における不活性な軸索の排除は神経活動に依存的な競合の結果である
歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウスでの不活性な軸索の排除は,ニューロンの神経活動には非依存的,あるいは,歯状回の成熟ニューロン以外のニューロンとの神経活動に依存的な競合,という2つの可能性が考えられた.この2つを区別するため,テトロドトキシンを海馬に投与しその全体の神経活動を抑制した.すると,破傷風菌毒素の発現により不活性な状態となったニューロンの軸索の排除は,テトロドトキシン非投与群に比較して著しく阻害された.このことより,歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウスにおける軸索の排除は,神経活動に依存的な競合によるものであることがわかった.
6.歯状回における軸索の排除は新生ニューロンの軸索との競合の結果である
それでは,歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウスでは,不活性なニューロンはどのニューロンと競合しているのだろうか? 可能性のひとつとして,歯状回では新しいニューロンが一生をつうじて産生されることから9),この新生ニューロンが不活性な成熟ニューロンと神経活動に依存的に競合していることが考えられた.新生ニューロンに破傷風菌毒素が発現しているかどうか新生ニューロンのマーカーを使い調べてみたところ発現していなかったことから,この可能性が支持された.その証明のため,歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウスを細胞分裂の阻害剤であるAraCにより処理することで新生ニューロンの産生を抑制し,不活性な軸索の排除への影響を調べてみた.すると,新生ニューロンの産生を抑制すると不活性な軸索の排除は著しく抑制されることがわかった.この発見を補強するため別の細胞分裂の阻害剤であるTemozolomideによる処理も行ったが,やはり同様の結果が得られた.
これらの結果から,歯状回において精緻化の起こる期間に新生したニューロンは,成熟ニューロンと効率的に競合しシナプス結合の精緻化にかかわるものと考えられた.そうであるなら,歯状回の新生ニューロンはすみやかにCA3領域のニューロンとシナプスを形成するだろう.これを証明するため,生後15日において歯状回の新生ニューロンにGFPを発現するレトロウイルスを感染させてこれを標識し,生後23日においてGFPを発現する軸索とシナプスの形態を観察した.その結果,生後15日に新生した歯状回のニューロンの軸索は,生後23日にはすでにCA3領域において標的の領域の末端にまで到達しており,歯状回の成熟ニューロンの軸索に形成されたシナプスと同等かそれ以上のシナプスが形成されていた.これらのシナプスは興奮性シナプス小胞や機能的前シナプスのマーカーにより染色されることから,すでに機能しているものと考えられた.
非競合的な状況である歯状回においてほとんどのニューロンを不活性化したトランスジェニックマウスと同様に,競合的な状況である歯状回において半分のニューロンが不活性化したトランスジェニックマウスでも,新生ニューロンの産生を抑制することで不活性な軸索の排除の抑制が観察されるのかどうか調べてみた.歯状回において成熟ニューロンどうしの競合があるなら,細胞分裂の阻害剤であるAraCにより処理することでニューロンの新生を抑えた場合でも不活性な軸索は排除されるはずである.ところがこの場合,不活性なニューロンの軸索の排除は抑制された.つまり,歯状回における不活性なニューロンの軸索の排除は,おもに不活性な成熟ニューロンと新生ニューロンとの競合の結果であり,成熟ニューロンどうしの競合によるものではないことが示唆された.
薬剤による処理では副作用の影響やニューロン新生の抑制の程度が問題になりうるため,遺伝学的にニューロン新生を抑制して神経活動の依存的な競合が成熟ニューロンと新生ニューロンのあいだで起こっているのかどうかを検討した.選択的かつ効率的に神経前駆細胞を死滅させることのできるチミジンキナーゼを発現するトランスジェニックマウスと,歯状回において半分のニューロンが不活性化したトランスジェニックマウスとを掛け合わせて実験に用いた10).その結果,このトランスジェニックマウスでは不活性な軸索はほとんど排除されなかった.この結果は,歯状回とCA3領域とのあいだでの神経活動に依存的な競合は成熟ニューロンと新生ニューロンとのあいだで起こり,成熟ニューロンどうしのあいだではほとんど起こらないことを証明した.また,チミジンキナーゼの発現はニューロン新生の起こらないCA1領域では不活性な軸索の排除に影響を及ぼさなかった.
おわりに
この研究において,筆者らは,神経活動に依存的な競合が海馬での神経回路の形成におけるシナプス結合の精緻化において一般的な原理であることをin vivoではじめて証明した.特筆すべきは,新しいニューロンがたえず産生される歯状回では,発生過程において成熟ニューロンが新生ニューロンと神経活動に依存的に競合することが示されたことである.この結果は,新生ニューロンがすでに形成され確立された成体の神経回路に,どのようにして効率よく組み込まれるのかを説明しているのかもしれない11,12).最近の研究では,歯状回におけるニューロンの新生は記憶の形成に重要であることが示されている.筆者らが発見した歯状回における神経活動に依存的な競合は,認知機能に重要と考えられるすみやかで効率的な新生ニューロンの組み込みの分子機構にかかわっているのかもしれない.
文 献
- O’Keefe, J. & Nadel, L.: The Hippocampus as a Cognitive Map. Oxford University Press, Oxford (1978)
- Squire, L. R., Stark, C. E. & Clark, R. E.: The medial temporal lobe. Annu. Rev. Neurosci., 27, 279-306 (2004)[PubMed]
- Amaral, D. G. & Witter, M. P.: The three-dimensional organization of the hippocampal formation: a review of anatomical data. Neuroscience, 31, 571-591 (1989)[PubMed]
- Sanes, J. R. & Lichtman, J. W.: Development of the vertebrate neuromuscular junction. Annu. Rev. Neurosci., 22, 389-442 (1999)[PubMed]
- Katz, L. C. & Shatz, C. J.: Synaptic activity and the construction of cortical circuits. Science, 274, 1133-1138 (1996)[PubMed]
- Yasuda, M. & Mayford, M. R.: CaMKII activation in the entorhinal cortex disrupts previously encoded spatial memory. Neuron, 50, 309-318 (2006)[PubMed]
- Yu, C. R., Power, J., Barnea, G. et al.: Spontaneous neural activity is required for the establishment and maintenance of the olfactory sensory map. Neuron, 42, 553-566 (2004)[PubMed]
- Bishop, D. L., Misgeld, T., Walsh, M. K. et al.: Axon branch removal at developing synapses by axosome shedding. Neuron, 44, 651-661 (2004)[PubMed]
- Gage, F. H.: Mammalian neural stem cells. Science, 287, 1433-1438 (2000)[PubMed]
- Singer, B. H., Jutkiewicz, E. M., Fuller, C. L. et al.: Conditional ablation and recovery of forebrain neurogenesis in the mouse. J. Comp. Neurol., 514, 567-582 (2009)[PubMed]
- Ge, S., Goh, E. L., Sailor, K. A. et al.: GABA regulates synaptic integration of newly generated neurons in the adult brain. Nature, 439, 589-593 (2006)[PubMed]
- Toni, N., Laplagne, D. A., Zhao, C. et al.: Neurons born in the adult dentate gyrus form functional synapses with target cells. Nat. Neurosci., 11, 901-907 (2008)[PubMed]
著者プロフィール
略歴:総合研究大学院大学,米国Scripps Resarch Instituteを経て,米国Michigan大学Medical School博士研究員.
梅森 久視(Hisashi Umemori)
米国Michigan大学Medical SchoolにてAssistant Professor.
© 2011 安田昌弘・梅森久視 Licensed under CC 表示 2.1 日本
