転写因子Glis1による体細胞初期化の促進
前川桃子1・山中伸弥2
(1京都大学ウイルス研究所 細胞生物学研究部門,2京都大学iPS細胞研究所 初期化機構研究部門)
email:前川桃子,山中伸弥
DOI: 10.7875/first.author.2011.102
Direct reprogramming of somatic cells is promoted by maternal transcription factor Glis1.
Momoko Maekawa, Kei Yamaguchi, Tomonori Nakamura, Ran Shibukawa, Ikumi Kodanaka, Tomoko Ichisaka, Yoshifumi Kawamura, Hiromi Mochizuki, Naoki Goshima, Shinya Yamanaka
Nature, 474, 225-229 (2011)
iPS細胞の最初の報告から5年が経過し臨床現場での早期活用が期待されるなか,応用のまえに解決すべき問題点がいくつか指摘されている.代表的な問題点のひとつが,山中4因子とよばれる最初に報告された初期化因子のひとつc-Mycを使用して作製したiPS細胞には腫瘍化するリスクのある点である.c-Mycを用いないiPS細胞の作製法も報告されているが,この場合,作製効率が格段に低くなってしまう.世界中の研究者によりさまざまなiPS細胞の作製法が試行錯誤的に検討および報告されてきたが,基礎となる初期化の分子機構が不明のままでは臨床応用の実現までどのくらいの時間がかかるのかわからない.筆者らは,さまざまな転写因子の初期化の能力を網羅的に調べることで新規の初期化因子をみつけるとともに,蓄積したデータを分子機構の解明に役立てようと計画した.この論文では,網羅的な探索からみつかった複数の新規の初期化因子のうちGlis1に注目し,iPS細胞の誘導過程におけるGlis1の効果を調べるとともにその機能解析を行った.
筆者らのグループは,2006年に世界ではじめて,マウスの分化細胞に4つの転写因子Oct3/4,Sox2,Klf4,c-Myc(山中4因子)を導入することで初期化を誘導し未分化の状態にもどすことのできることを報告した1).この初期化を誘導された細胞をiPS細胞と名づけ,2007年には,ヒトの分化細胞からもiPS細胞の誘導できることを報告した2).そののち,iPS細胞は世界中で熱く研究される分野となり再生医療への応用が大きく期待されている.しかしながら,臨床現場にiPS細胞をもっていくまえに,基礎研究において解決しておかなければならないことがある.いくつかの問題点のあるうち,筆者らは,以下の3つに注目した.1)初期化因子のうちc-Mycはがん遺伝子産物であるという一面をもちあわせ,それゆえc-Mycを用いて作製したiPS細胞は腫瘍化する危険性がある.2)c-Mycを除く3つの初期化因子によりiPS細胞を作製する方法も報告されているが,作製効率が格段に低くなってしまう3).3)分化細胞が初期化されて未分化細胞へともどる分子機構はほとんどわかっていない.筆者らは,新規の初期化因子の網羅的な探索プロジェクトを立ち上げ,よりよい初期化因子を探索するとともに,新規の初期化因子のデータを蓄積することで初期化の分子機構に対する理解を深めることにした.
新規の初期化因子の網羅的な探索においては,これまで構築してきたcDNAライブラリーから選び出した1437個の転写因子を用いた4).偽陽性を減らし網羅的な探索の精度を上げるため,最初の4つの初期化因子のうちiPS細胞の誘導に必須であったOct3/4とKlf4のそれぞれの代替が可能となる転写因子を探した.また,初期化されるとGFP陽性となるマウスの細胞を用いることで結果の解析を行いやすくした5).その結果,1437個の転写因子から,Oct3/4を代替できる転写因子はひとつもみつけることはできなかったが,Klf4の代替となる転写因子は新規に18個同定することができた(図1).
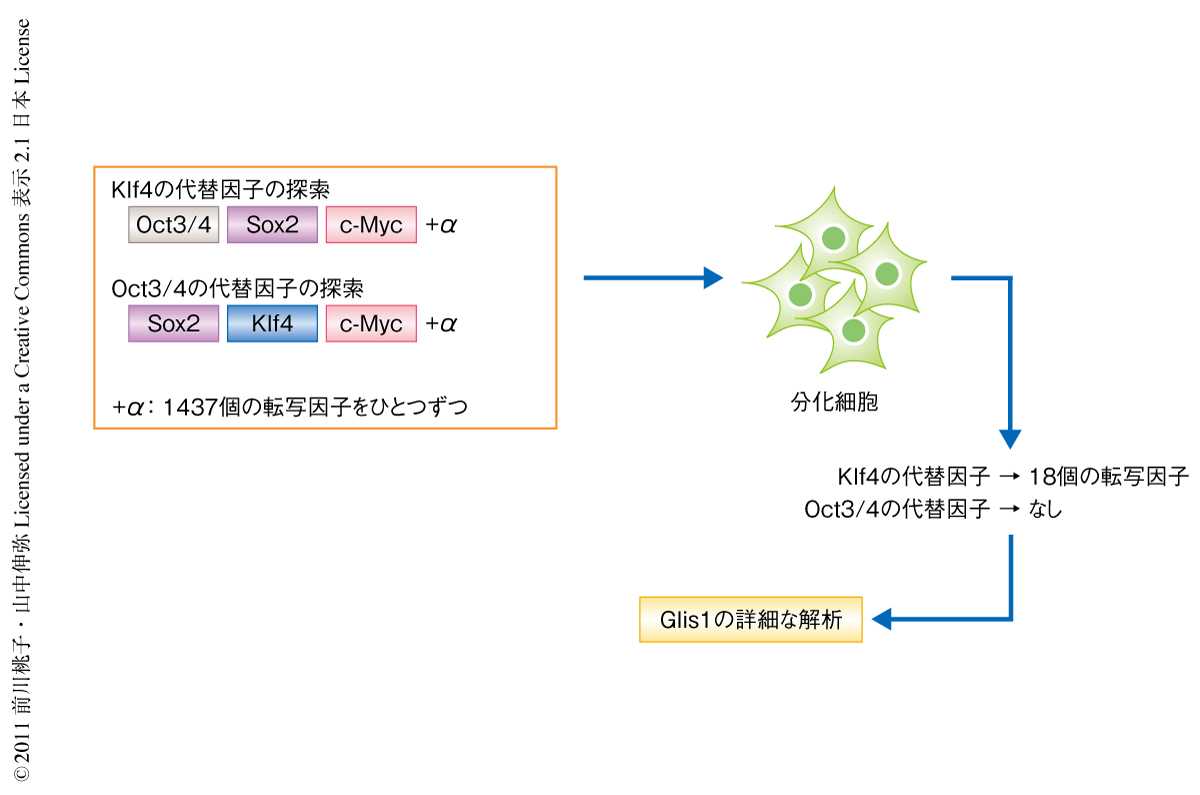
新規の初期化因子18個のうち,Glis1という転写因子に注目して研究を進めた6).このGlis1には初期化に関する報告はまったくなく,最初の4つの初期化因子がES細胞で高発現する遺伝子のなかから選ばれたのに対し,今回,転写因子全体からランダムに探索したことによりはじめてみつかったものといえる.
さまざまな細胞におけるGlis1の発現パターンを調べた.Glis1は未受精卵や受精卵で非常に高く発現しており,いろいろな組織や繊維芽細胞,あるいは,ES細胞ではその発現量は低かった.未受精卵や受精卵で高発現するということからGlis1には大きな期待がもたれた.なぜなら,iPS細胞の作製には数週間という長い時間がかかり効率も非常に低いのに対し,卵への核移植では短時間に高効率に初期化が起こるからである7,8).卵のもつ非常に高い初期化の能力をiPS細胞の作製において再現することができるかもしれないと考えた.
Glis1はKlf4の代替となる転写因子としてみつかってきたが,Klf4の代わりに用いたときのiPS細胞の誘導効率は最初の4つの初期化因子に比較すると低かった.そこで,Glis1とKlf4の両方を用いてiPS細胞の誘導を行ってみたところ非常に驚きの結果が得られた.4つの初期化因子のうちc-Mycを除く3つの初期化因子といっしょに用いた場合,Glis1はc-Mycと同じ程度の数のiPS細胞を誘導した(図2).また,c-Mycを含む4つの初期化因子といっしょに用いた場合,Glis1とc-MycはiPS細胞の数を相乗的に増加させた.
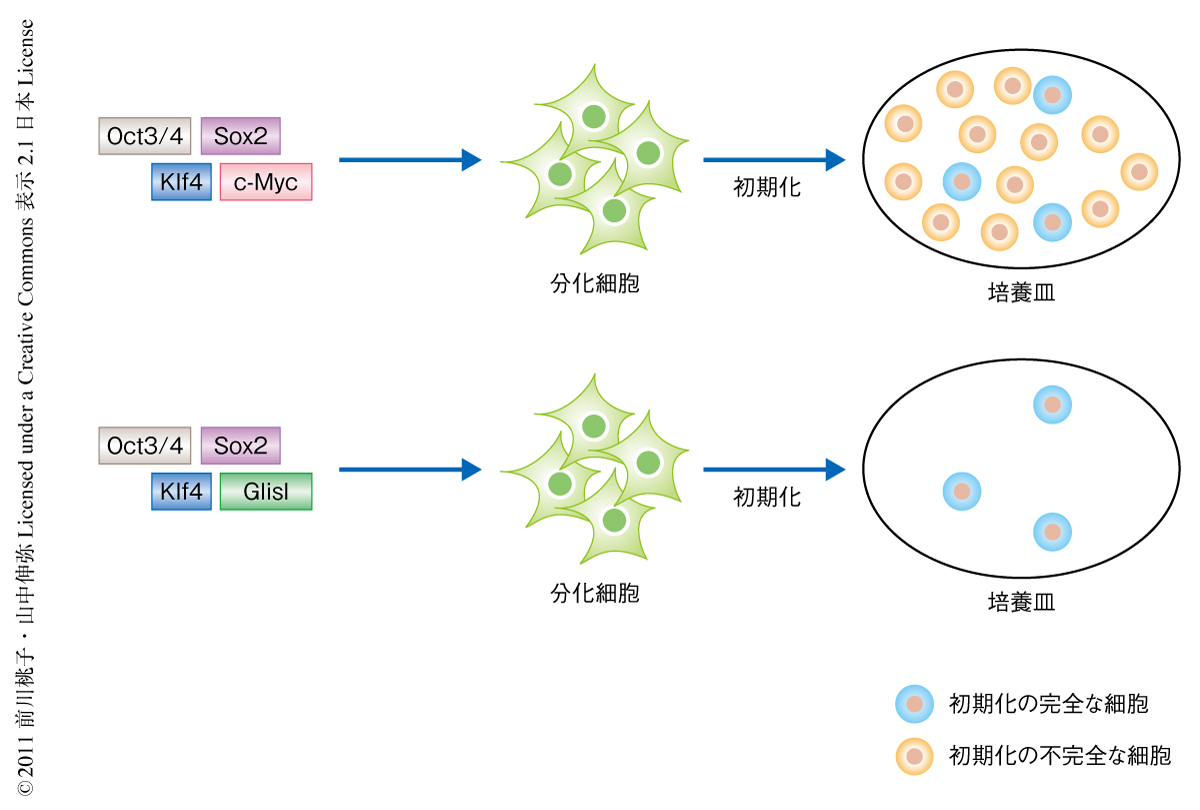
一方,iPS細胞を誘導すると,初期化の完全な細胞すなわちiPS細胞とともに,初期化の不完全な細胞すなわちiPS細胞になりそこねた細胞も現われる.これら初期化の不完全な細胞は生体に移植した場合に腫瘍化するという報告がある.マウスの細胞の場合,c-Mycを用いてiPS細胞の誘導を行うと初期化の完全な細胞を上回る数の初期化の不完全な細胞が現われるが,c-Mycの代わりにGlis1を用いるとほとんどの細胞が初期化の完全な細胞となった(図2).また,Glis1をc-Mycを含む4つの初期化因子といっしょに用いた場合もほとんどの細胞が初期化の完全な細胞となり,Glis1は非常にすぐれた能力をもつことがわかった.また,ヒト細胞を用いた場合もGlis1はよく似た効果を示すことがわかった.
さらに,Glis1を用いて作製したiPS細胞について詳細な解析を行った.c-Mycの代わりにGlis1を用いて作製したマウスのiPS細胞からはキメラマウスを作製することができ,さらに生殖系譜への寄与も確認された.ヒトの細胞についてもGlis1を用いて作製したiPS細胞は試験管内および生体内のどちらにおいても三胚葉へと分化できることを確認した.よって,c-Mycの代わりにGlis1を用いて作製したiPS細胞はマウスとヒトどちらの細胞においても分化能には問題のないことがわかった.
さきに述べたように,ES細胞ではGlis1はほとんど発現していない.そこで,未分化な細胞におけるGlis1の機能を調べてみようと,マウスのES細胞にGlis1を強制発現してみた.その結果,意外にもGlis1を強制発現したES細胞はほとんど増殖できないことがわかった.このことから,iPS細胞の誘導過程において細胞に導入された初期化因子の発現が抑制されないような初期化の不完全な細胞では,Glis1の発現が継続することにより細胞の増殖が抑制されていることが示唆された.つまり,増殖している細胞は完全に初期化されたiPS細胞であることを示していた.Glis1を用いたiPS細胞の作製法は完全に初期化されたiPS細胞のみを増殖させることから,より安全な方法といえるだろう.今後は,ヒト細胞でも同様の検討が必要である.
筆者らのグループは,p53経路を抑制するとiPS細胞の作製効率の向上することを報告している9).そこで,p53経路とGlis1との関係を調べてみることにした.まず,p53ノックアウト細胞にGlis1を導入してiPS細胞を誘導した.もし,野生型の細胞で観察されたようなGlis1の効果がp53ノックアウト細胞において観察されなかったのなら,Glis1とp53は初期化において同じ経路で機能している,つまり,Glis1はp53を抑制することで初期化を促進していることになる.しかしながら,実際にはp53ノックアウト細胞においても野生型の細胞と同様にGlis1は効果を発揮していた.よって,Glis1はp53経路とは別の分子機構で初期化を促進していることが示唆された.
つぎに,初期化の早期における遺伝子発現の解析を行うことでGlis1の機能を調べることにした.マウスの野生型の細胞に最初の4つの初期化因子のうちc-Mycを除く3つの初期化因子,または,それにくわえGlis1を導入したところ,5日目において初期化されていた細胞はそれぞれ約0.2%,または,約0.7%であった.マウスのp53ノックアウト細胞の場合は,c-Mycを除く3つの初期化因子を導入した場合は約2%,それにくわえGlis1を導入した場合は約17%であった.よって,解析に十分な細胞数を得るためp53ノックアウト細胞にc-Mycを除く3つの初期化因子またはそれにくわえGlis1を導入し,初期化された細胞のみを回収して遺伝子発現の解析を行った.その結果,Glis1は初期化の誘導に寄与することが報告されている複数の遺伝子の発現を上昇させていることがわかった.つまり,Glis1は複数の初期化の経路を活性化させることによりiPS細胞の作製を促進していることが明らかになった.
Glis1の発見は少なくとも基礎研究においては非常に画期的なものであった.さきに述べたiPS細胞の作製に残された3つの問題点のうち,1つ目と2つ目については,Glis1がc-Mycと同じ程度に効率よく,さらに初期化の不完全な細胞の増殖を抑えて完全に初期化された安全な細胞のみを増殖させるという効果でほぼ解決できる.また,3つ目の初期化の分子機構については,ここで述べたGlis1の機能解析にくわえ,網羅的な探索においてみつかってきたGlis1以外の新規の初期化因子の情報を活用することで,今後,ますます明らかにされていくであろう.初期化因子の網羅的な探索そしてGlis1の発見が基礎研究から応用研究へと進展し,iPS細胞が臨床現場で活躍する日の1日でも早く実現することを願っている.
略歴:2007年 京都大学大学院生命科学研究科博士課程 修了,同年 同 再生医科学研究所 研究員,2008年 同 物質-細胞統合システム拠点 特任研究員,2010年 同iPS細胞研究所 特任研究員を経て,2011年より同 ウイルス研究所 助教.
研究テーマ:幹細胞の分化過程における対称あるいは非対称の分裂.
抱負:マイペースに楽しく研究.
山中 伸弥(Shinya Yamanaka)
京都大学iPS細胞研究所 教授.
研究室URL:http://www.cira.kyoto-u.ac.jp/yamanaka_group/
© 2011 前川桃子・山中伸弥 Licensed under CC 表示 2.1 日本
(1京都大学ウイルス研究所 細胞生物学研究部門,2京都大学iPS細胞研究所 初期化機構研究部門)
email:前川桃子,山中伸弥
DOI: 10.7875/first.author.2011.102
Direct reprogramming of somatic cells is promoted by maternal transcription factor Glis1.
Momoko Maekawa, Kei Yamaguchi, Tomonori Nakamura, Ran Shibukawa, Ikumi Kodanaka, Tomoko Ichisaka, Yoshifumi Kawamura, Hiromi Mochizuki, Naoki Goshima, Shinya Yamanaka
Nature, 474, 225-229 (2011)
要 約
iPS細胞の最初の報告から5年が経過し臨床現場での早期活用が期待されるなか,応用のまえに解決すべき問題点がいくつか指摘されている.代表的な問題点のひとつが,山中4因子とよばれる最初に報告された初期化因子のひとつc-Mycを使用して作製したiPS細胞には腫瘍化するリスクのある点である.c-Mycを用いないiPS細胞の作製法も報告されているが,この場合,作製効率が格段に低くなってしまう.世界中の研究者によりさまざまなiPS細胞の作製法が試行錯誤的に検討および報告されてきたが,基礎となる初期化の分子機構が不明のままでは臨床応用の実現までどのくらいの時間がかかるのかわからない.筆者らは,さまざまな転写因子の初期化の能力を網羅的に調べることで新規の初期化因子をみつけるとともに,蓄積したデータを分子機構の解明に役立てようと計画した.この論文では,網羅的な探索からみつかった複数の新規の初期化因子のうちGlis1に注目し,iPS細胞の誘導過程におけるGlis1の効果を調べるとともにその機能解析を行った.
はじめに
筆者らのグループは,2006年に世界ではじめて,マウスの分化細胞に4つの転写因子Oct3/4,Sox2,Klf4,c-Myc(山中4因子)を導入することで初期化を誘導し未分化の状態にもどすことのできることを報告した1).この初期化を誘導された細胞をiPS細胞と名づけ,2007年には,ヒトの分化細胞からもiPS細胞の誘導できることを報告した2).そののち,iPS細胞は世界中で熱く研究される分野となり再生医療への応用が大きく期待されている.しかしながら,臨床現場にiPS細胞をもっていくまえに,基礎研究において解決しておかなければならないことがある.いくつかの問題点のあるうち,筆者らは,以下の3つに注目した.1)初期化因子のうちc-Mycはがん遺伝子産物であるという一面をもちあわせ,それゆえc-Mycを用いて作製したiPS細胞は腫瘍化する危険性がある.2)c-Mycを除く3つの初期化因子によりiPS細胞を作製する方法も報告されているが,作製効率が格段に低くなってしまう3).3)分化細胞が初期化されて未分化細胞へともどる分子機構はほとんどわかっていない.筆者らは,新規の初期化因子の網羅的な探索プロジェクトを立ち上げ,よりよい初期化因子を探索するとともに,新規の初期化因子のデータを蓄積することで初期化の分子機構に対する理解を深めることにした.
1.新規の初期化因子の網羅的な探索
新規の初期化因子の網羅的な探索においては,これまで構築してきたcDNAライブラリーから選び出した1437個の転写因子を用いた4).偽陽性を減らし網羅的な探索の精度を上げるため,最初の4つの初期化因子のうちiPS細胞の誘導に必須であったOct3/4とKlf4のそれぞれの代替が可能となる転写因子を探した.また,初期化されるとGFP陽性となるマウスの細胞を用いることで結果の解析を行いやすくした5).その結果,1437個の転写因子から,Oct3/4を代替できる転写因子はひとつもみつけることはできなかったが,Klf4の代替となる転写因子は新規に18個同定することができた(図1).
2.Glis1の発現パターン
新規の初期化因子18個のうち,Glis1という転写因子に注目して研究を進めた6).このGlis1には初期化に関する報告はまったくなく,最初の4つの初期化因子がES細胞で高発現する遺伝子のなかから選ばれたのに対し,今回,転写因子全体からランダムに探索したことによりはじめてみつかったものといえる.
さまざまな細胞におけるGlis1の発現パターンを調べた.Glis1は未受精卵や受精卵で非常に高く発現しており,いろいろな組織や繊維芽細胞,あるいは,ES細胞ではその発現量は低かった.未受精卵や受精卵で高発現するということからGlis1には大きな期待がもたれた.なぜなら,iPS細胞の作製には数週間という長い時間がかかり効率も非常に低いのに対し,卵への核移植では短時間に高効率に初期化が起こるからである7,8).卵のもつ非常に高い初期化の能力をiPS細胞の作製において再現することができるかもしれないと考えた.
3.Glis1を用いたiPS細胞の誘導
Glis1はKlf4の代替となる転写因子としてみつかってきたが,Klf4の代わりに用いたときのiPS細胞の誘導効率は最初の4つの初期化因子に比較すると低かった.そこで,Glis1とKlf4の両方を用いてiPS細胞の誘導を行ってみたところ非常に驚きの結果が得られた.4つの初期化因子のうちc-Mycを除く3つの初期化因子といっしょに用いた場合,Glis1はc-Mycと同じ程度の数のiPS細胞を誘導した(図2).また,c-Mycを含む4つの初期化因子といっしょに用いた場合,Glis1とc-MycはiPS細胞の数を相乗的に増加させた.
一方,iPS細胞を誘導すると,初期化の完全な細胞すなわちiPS細胞とともに,初期化の不完全な細胞すなわちiPS細胞になりそこねた細胞も現われる.これら初期化の不完全な細胞は生体に移植した場合に腫瘍化するという報告がある.マウスの細胞の場合,c-Mycを用いてiPS細胞の誘導を行うと初期化の完全な細胞を上回る数の初期化の不完全な細胞が現われるが,c-Mycの代わりにGlis1を用いるとほとんどの細胞が初期化の完全な細胞となった(図2).また,Glis1をc-Mycを含む4つの初期化因子といっしょに用いた場合もほとんどの細胞が初期化の完全な細胞となり,Glis1は非常にすぐれた能力をもつことがわかった.また,ヒト細胞を用いた場合もGlis1はよく似た効果を示すことがわかった.
さらに,Glis1を用いて作製したiPS細胞について詳細な解析を行った.c-Mycの代わりにGlis1を用いて作製したマウスのiPS細胞からはキメラマウスを作製することができ,さらに生殖系譜への寄与も確認された.ヒトの細胞についてもGlis1を用いて作製したiPS細胞は試験管内および生体内のどちらにおいても三胚葉へと分化できることを確認した.よって,c-Mycの代わりにGlis1を用いて作製したiPS細胞はマウスとヒトどちらの細胞においても分化能には問題のないことがわかった.
4.安全なiPS細胞の作製法
さきに述べたように,ES細胞ではGlis1はほとんど発現していない.そこで,未分化な細胞におけるGlis1の機能を調べてみようと,マウスのES細胞にGlis1を強制発現してみた.その結果,意外にもGlis1を強制発現したES細胞はほとんど増殖できないことがわかった.このことから,iPS細胞の誘導過程において細胞に導入された初期化因子の発現が抑制されないような初期化の不完全な細胞では,Glis1の発現が継続することにより細胞の増殖が抑制されていることが示唆された.つまり,増殖している細胞は完全に初期化されたiPS細胞であることを示していた.Glis1を用いたiPS細胞の作製法は完全に初期化されたiPS細胞のみを増殖させることから,より安全な方法といえるだろう.今後は,ヒト細胞でも同様の検討が必要である.
5.iPS細胞の誘導過程におけるGlis1の機能
筆者らのグループは,p53経路を抑制するとiPS細胞の作製効率の向上することを報告している9).そこで,p53経路とGlis1との関係を調べてみることにした.まず,p53ノックアウト細胞にGlis1を導入してiPS細胞を誘導した.もし,野生型の細胞で観察されたようなGlis1の効果がp53ノックアウト細胞において観察されなかったのなら,Glis1とp53は初期化において同じ経路で機能している,つまり,Glis1はp53を抑制することで初期化を促進していることになる.しかしながら,実際にはp53ノックアウト細胞においても野生型の細胞と同様にGlis1は効果を発揮していた.よって,Glis1はp53経路とは別の分子機構で初期化を促進していることが示唆された.
つぎに,初期化の早期における遺伝子発現の解析を行うことでGlis1の機能を調べることにした.マウスの野生型の細胞に最初の4つの初期化因子のうちc-Mycを除く3つの初期化因子,または,それにくわえGlis1を導入したところ,5日目において初期化されていた細胞はそれぞれ約0.2%,または,約0.7%であった.マウスのp53ノックアウト細胞の場合は,c-Mycを除く3つの初期化因子を導入した場合は約2%,それにくわえGlis1を導入した場合は約17%であった.よって,解析に十分な細胞数を得るためp53ノックアウト細胞にc-Mycを除く3つの初期化因子またはそれにくわえGlis1を導入し,初期化された細胞のみを回収して遺伝子発現の解析を行った.その結果,Glis1は初期化の誘導に寄与することが報告されている複数の遺伝子の発現を上昇させていることがわかった.つまり,Glis1は複数の初期化の経路を活性化させることによりiPS細胞の作製を促進していることが明らかになった.
おわりに
Glis1の発見は少なくとも基礎研究においては非常に画期的なものであった.さきに述べたiPS細胞の作製に残された3つの問題点のうち,1つ目と2つ目については,Glis1がc-Mycと同じ程度に効率よく,さらに初期化の不完全な細胞の増殖を抑えて完全に初期化された安全な細胞のみを増殖させるという効果でほぼ解決できる.また,3つ目の初期化の分子機構については,ここで述べたGlis1の機能解析にくわえ,網羅的な探索においてみつかってきたGlis1以外の新規の初期化因子の情報を活用することで,今後,ますます明らかにされていくであろう.初期化因子の網羅的な探索そしてGlis1の発見が基礎研究から応用研究へと進展し,iPS細胞が臨床現場で活躍する日の1日でも早く実現することを願っている.
文 献
- Takahashi, K. & Yamanaka, S.: Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell, 126, 663-676 (2006)[PubMed]
- Takahashi, K., Tanabe, K., Ohnuki, M. et al.: Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell, 131, 861-872 (2007)[PubMed]
- Nakagawa, M., Koyanagi, M., Tanabe, K. et al.: Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat. Biotechnol., 26, 101-106 (2008)[PubMed]
- Goshima, N., Kawamura, Y., Fukumoto, A. et al.: Human protein factory for converting the transcriptome into an in vitro-expressed proteome. Nat. Methods, 5, 1011-1017 (2008)[PubMed]
- Okita, K., Ichisaka, T. & Yamanaka, S.: Generation of germ-line competent induced pluripotent stem cells. Nature, 448, 313-317 (2007)[PubMed]
- Kim, Y. S., Lewandoski, M., Perantoni, A. O. et al.: Identification of Glis1, a novel Gli-related, Kruppel-like zinc finger protein containing transactivation and repressor functions. J. Biol. Chem., 277, 30901-30913 (2002)[PubMed]
- Wilmut, I., Schnieke, A. E., McWhir, J. et al.: Viable offspring derived from fetal and adult mammalian cells. Nature, 385, 810-813 (1997)[PubMed]
- Egli, D., Rosains, J., Birkhoff, G. et al.: Developmental reprogramming after chromosome transfer into mitotic mouse zygotes. Nature, 447, 679-685 (2007)[PubMed]
- Hong, H., Takahashi, K., Ichisaka, T. et al.: Suppression of induced pluripotent stem cell generation by the p53-p21 pathway. Nature, 460, 1132-1135 (2009)[PubMed]
著者プロフィール
略歴:2007年 京都大学大学院生命科学研究科博士課程 修了,同年 同 再生医科学研究所 研究員,2008年 同 物質-細胞統合システム拠点 特任研究員,2010年 同iPS細胞研究所 特任研究員を経て,2011年より同 ウイルス研究所 助教.
研究テーマ:幹細胞の分化過程における対称あるいは非対称の分裂.
抱負:マイペースに楽しく研究.
山中 伸弥(Shinya Yamanaka)
京都大学iPS細胞研究所 教授.
研究室URL:http://www.cira.kyoto-u.ac.jp/yamanaka_group/
© 2011 前川桃子・山中伸弥 Licensed under CC 表示 2.1 日本
