毛包幹細胞と色素幹細胞との協調的なWntシグナルの活性化により色素をともなう毛の再生が起こる
武尾 真・伊藤真由美
(米国New York大学School of Medicine,Department of Dermatology and Cell Biology)
email:武尾 真
DOI: 10.7875/first.author.2011.104
Coordinated activation of Wnt in epithelial and melanocyte stem cells initiates pigmented hair regeneration.
Piul Rabbani, Makoto Takeo, WeiChin Chou, Peggy Myung, Marcus Bosenberg, Lynda Chin, M. Mark Taketo, Mayumi Ito
Cell, 145, 941-955 (2011)
哺乳類の毛髪には寿命があり,毛包バルジで毛包幹細胞と色素幹細胞とが同時に活性化することにより色のついた毛髪がくり返し再生する.しかし,2つの異なる幹細胞が同時に活性化し協調的に毛髪を再生する分子機構はわかっていなかった.今回,筆者らは,Wntシグナル経路に注目し,細胞特異的にWntシグナル経路を変更することによりその分子機構の解明を試みた.その結果,毛髪の再生が開始するとき毛包幹細胞からWntリガンドが分泌され,毛包幹細胞と色素幹細胞とでWntシグナル経路が同時に活性化することが明らかになった.これらの結果は,毛包の再生過程においてWntシグナルが毛包幹細胞と色素幹細胞との協調的な挙動を可能にする分子機構としてはたらいていることを示すばかりでなく,多くの細胞種が関係する哺乳類の器官再生に関する新たな知見をあたえてくれる.
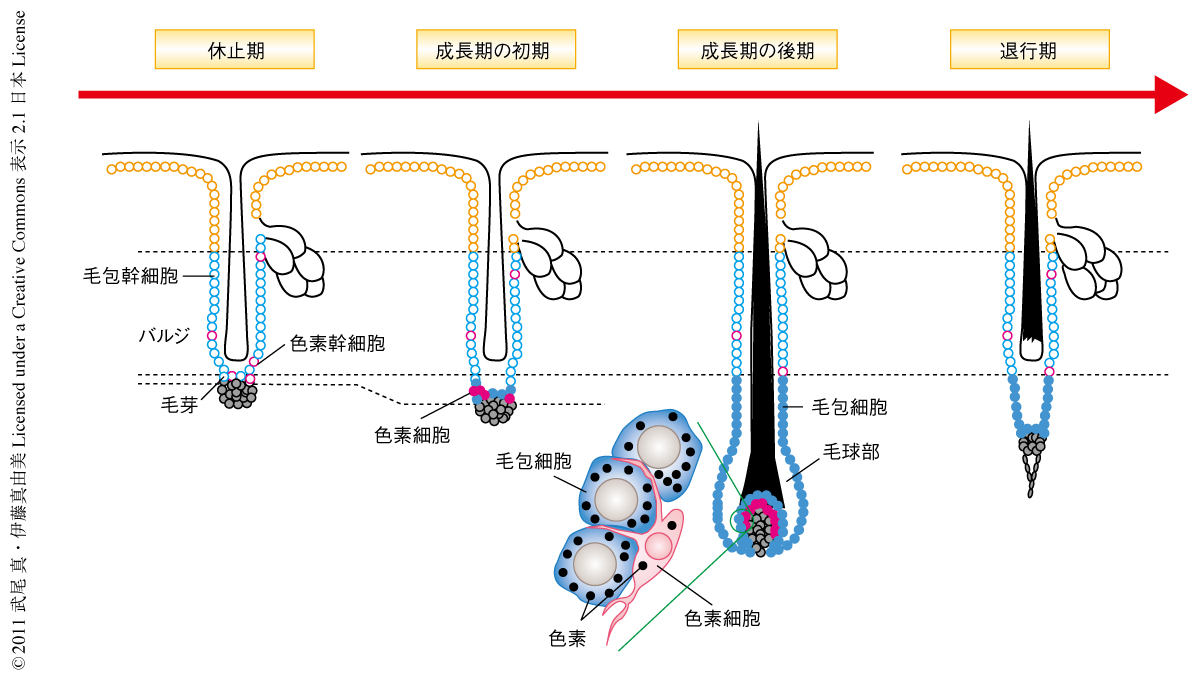
哺乳類の毛髪には寿命があり生涯をつうじて再生と脱毛とをくり返す(毛周期).1回の毛周期は,毛包の下部が再生し毛髪がつくりだされる成長期,毛髪の伸長が止まり毛包下部が収縮する退行期,および,休止期の3つの時期に分けられる(図1).毛包のバルジ領域および毛芽領域には毛包幹細胞と色素幹細胞がある1,2).成長期にこの2つの幹細胞が同時に活性化し,毛包幹細胞の子孫細胞により毛包下部が再生され毛球部がつくられる.同時に,色素幹細胞の子孫細胞が毛球部へと移動し色素細胞へと分化する2).毛球部では毛包幹細胞の子孫細胞が毛髪に分化する過程で色素細胞によりつくられた色素を取り込み色のついた毛髪が再生する.このように,成長期の毛包の再生過程において毛包幹細胞と色素幹細胞とが同時に活性化し協調的に色のついた毛髪を再生させることは古くから知られていたが,その分子機構はわかっていなかった.
これまでに,毛包の再生過程において毛包幹細胞の活性化にかかわる分子機構の研究が精力的に行われ,Wntシグナル経路が重要な機構のひとつであることが示されている3,4).また,色素細胞の発生にはWntシグナルが必要であること5,6),培養細胞ではWntシグナルが色素細胞の分化を誘導することから7),色素幹細胞の分化にもWntシグナルが重要な役割をはたしているものと考えられる.しかしながら,成体の色素幹細胞に関しては幹細胞の維持に注目した研究は行われているが8),活性化の機構はほとんどわかっていない.そのため,成体の色素幹細胞におけるWntシグナルのはたらきはわかっていない.そこで筆者らは,毛包幹細胞または色素幹細胞に特異的な遺伝子プロモーターを使い,細胞特異的にWntシグナルを抑制または恒常的に活性化させ,成体の色素幹細胞の活性化にWntシグナルが必要であるかどうかを明らかにしようと試みた.また,毛包が再生されるときにどのようにして細胞系譜の異なる2種類の幹細胞が同時に活性化して分裂および分化するのか,その分子機構の解明も試みた.この研究により得られた結果は,単に毛包幹細胞と色素幹細胞とが協調的にはたらくための分子機構を明らかにするだけでなく,哺乳類の成体における多くの細胞種を含む複雑な器官の再生を可能にするヒントをあたえてくれるものと期待される.
毛包が再生するとき,色素幹細胞ではWntシグナル経路が活性化しているのだろうか? Wntシグナル経路が活性化するとβカテニンが核へと移行する.この核に局在したβカテニンをWntシグナル経路の活性化の指標とし,マウスの毛包において成長期に色素幹細胞ではWntシグナル経路が活性化しているのかどうか調べた.休止期には毛包幹細胞でも色素幹細胞でも核におけるβカテニンの発現はみられなかったが,成長期がはじまると毛芽領域において毛包幹細胞と色素幹細胞の両方でWntシグナル経路の活性化が同時に観察された.成長期が進んで毛球部がつくられるとWntシグナル経路の活性化は毛球部の毛包細胞と色素細胞だけにみられるようになった.毛周期をつうじてバルジ領域の色素幹細胞においてWntシグナル経路の活性化することはなかった.この結果から,毛包が再生するときに毛芽領域の色素幹細胞においてWntシグナルが活性化することが明らかになった.また,成長期でのWntシグナル経路の活性化がヒトの頭髪でもみられたことから,マウスとヒトの毛包の再生では同じ分子機構のはたらいていることが考えられた.
Wntシグナル経路の活性化が色素細胞の分化に必要かどうかを調べるため,休止期の色素幹細胞を含むすべての色素細胞においてWntシグナル経路を活性化しつづけるトランスジェニックマウスを作製した.このトランスジェニックマウスでは通常はWntシグナル経路の活性化していないバルジ領域の色素幹細胞でも色素細胞の分化マーカーが発現し,異所的に色素もつくっていた.一方で,普段からWntシグナル経路の活性化している毛球部の色素細胞では分化マーカーの発現と色素の産生について対照マウスとの違いはみられなかった.このことから,Wntシグナルの活性化は色素幹細胞の分化を誘導するのに十分であることが明らかになった.
おもしろいことに,このトランスジェニックマウスでは毛周期が進むにつれて白髪が増加し,4回目の毛周期ではすべてのトランスジェニックマウスで全体の20%以上が白髪になった.一方,対照マウスでは白髪の増加はみられなかった.これまで,バルジ領域の色素幹細胞が活性化することにより自己複製能力が低下し,最終的には色素幹細胞がなくなり白髪になることが報告されている9).そこで,このトランスジェニックマウスでも同じ理由で白髪が増加しているのかどうか調べた.その結果,トランスジェニックマウスでは毛周期が進むにつれてバルジ領域の色素幹細胞が減少していた.これらの結果から,恒常的なWntシグナル経路の活性化は色素幹細胞の分化をひき起こし,自己複製能を低下させ色素幹細胞の減少とそれにともなう白髪をひき起こすことが明らかになった.
Wntシグナル経路の恒常的な活性化は色素幹細胞の分化の誘導に十分であることはわかったが,それでは,通常の毛包の再生はどうだろうか? 通常の毛包の再生におけるWntシグナルの役割を調べるため別のトランスジェニックマウスを作製した.このトランスジェニックマウスではタモキシフェンを投与することで色素細胞に特異的にWntシグナル経路の構成タンパク質のひとつであるβカテニンをコードする遺伝子をゲノムから取り除くことができる.βカテニンのなくなった細胞ではWntシグナル経路の活性化は起こらない.このトランスジェニックマウスにおいてβカテニンを取り除くと細胞周期の進行にともない白髪が有意に増加した.また,白髪の毛包には毛球部の色素細胞の数が少なく,色素細胞での分化マーカーの発現もみられなかった.一方,バルジ領域では色素幹細胞の数と分化状態に違いはみられなかった.
このトランスジェニックマウスの毛球部において色素細胞が減少する原因として,色素細胞が分裂できない可能性と色素細胞が死滅する可能性の2つが考えられた.これらの可能性を確かめるため,成長期での細胞分裂の頻度をBrdUの取り込み率を指標に比較したところ,トランスジェニックマウスではBrdUを取り込む色素細胞の割合が対照マウスに比べ約半分にまで減少していた.一方,細胞死のマーカーである活性化型カスパーゼ3はトランスジェニックマウスと対照マウスの両方において検出できなかった.これらの結果から,通常の毛包の再生においても色素幹細胞の増殖と分化にWntシグナル経路の活性化の必要であることが明らかになった.さきに述べたように,毛包幹細胞の増殖と分化にもWntシグナル経路がかかわっていた.また,毛包が再生するとき毛芽領域では毛包幹細胞と色素幹細胞で同時にWntシグナル経路が活性化した.これらのことから,Wntシグナル経路が2つの幹細胞が協調的に毛髪を再生するための分子機構としてはたらいているではないかと予想された.
Wntシグナル経路はWntリガンドが受容体に結合することで活性化する.色素幹細胞を活性化するWntリガンドについて調べるため,神経堤から分化したすべての色素細胞がYFPを発現するトランスジェニックマウスから蛍光セルソーターにより色素幹細胞を単離し,すべてのWntリガンドの発現量の変化を調べた.しかしながら,成長期を誘導する前後でWntリガンドの発現の上昇は検出されなかった.
自らWntリガンドをつくりださずに,色素幹細胞はいったいどのようにWntシグナル経路を活性化しているのだろうか? 幹細胞説によると,幹細胞は内因的な要因とニッチからの外因的な要因により挙動が制御されている.そこで,バルジ領域および毛芽領域において色素幹細胞のとなりに存在する毛包幹細胞に注目した.まず,毛包幹細胞においてGFPを発現するトランスジェニックマウスから毛包幹細胞を単離し,リアルタイムPCR法によりWntリガンドの発現量を調べた.この結果,成長期を誘導した毛包幹細胞においていくつかのWntリガンドの発現量の増加することが確認された.また,in situハイブリダイゼーション法によりWntリガンドを発現している細胞は毛芽領域の毛包幹細胞であることも確認された.
つぎに,上皮細胞から分泌されたWntリガンドにより色素細胞のWntシグナル経路が活性化するかどうかを調べるため,Wnt10bの発現ベクターを導入した上皮細胞と色素細胞との共培養実験を行ったところ,色素細胞のWntシグナル経路の活性が約5倍まで増加した.さらに,生体においても外部からのWntリガンドにより色素幹細胞は活性化するかどうかを調べるためWntリガンドの皮内注入実験を行った.この実験には毛包幹細胞のWntシグナル経路の活性化による2次的な効果を排除するため毛包細胞に特異的にβカテニンを欠失させたトランスジェニックマウスを使った.このトランスジェニックマウスではWnt7aの注入により色素幹細胞においてWntシグナル経路の活性化がみられた.また,色素細胞の分化マーカーの発現も確認された.これらの結果から,毛包が再生するとき毛包幹細胞から分泌されるWntリガンドにより,毛包幹細胞と色素幹細胞とでWntシグナル経路が同時に活性化されることが示唆された.
毛包の再生過程において毛包幹細胞と色素幹細胞でWntシグナル経路が同時に活性化することにはどんな意味があるのだろうか? 毛包幹細胞でのWntシグナル経路の活性化が色素幹細胞の挙動にどのような影響をあたえるのか調べるため,バルジ領域を含むすべての毛包幹細胞においてWntシグナル経路を活性化しつづけるトランスジェニックマウスを作製した.このトランスジェニックマウスではバルジ領域にWntシグナル経路の活性化した肥大部が形成され,その領域で多くの色素細胞が観察された.また,バルジ領域の色素細胞の数も有意に増加していた.同様の結果は,すべての上皮基底細胞においてWntシグナル経路の活性化しつづける別のトランスジェニックマウスでも観察された.通常,マウスの皮膚に色素細胞はないが,おもしろいことに,このトランスジェニックマウスではバルジ領域だけではなく表皮にも多数の色素細胞がみられた.これらの結果は,毛包細胞でのWntシグナルの活性化は色素細胞の分裂を促進することを示していた.
毛包幹細胞でのWntシグナル経路の活性化がどのようにして色素細胞の増殖を促進するのか調べるため,バルジ領域を含むすべての毛包幹細胞においてWntシグナル経路を活性化しつづけるさきのトランスジェニックマウスと対照マウスとで発現している遺伝子を比較した.全層皮膚を使ったマイクロアレイ法による解析の結果,Wntシグナルの直接のターゲットであり色素細胞増殖因子と考えられるEndothelin1(Edn1)の発現がトランスジェニックマウスにおいて有意に増加していることが明らかになった.
全層皮膚には毛包幹細胞と色素幹細胞のほかにも多くの細胞が含まれる.そこで,このトランスジェニックマウスから蛍光セルソーターにより単離した上皮幹細胞と色素幹細胞を使いリアルタイムPCR法による解析を行った.その結果,毛包の再生の初期に毛包幹細胞においてEdn1とEdn2の発現が,色素幹細胞においてすべてのEdnの受容体であるEdn受容体B(EdnrB)の発現が増加することが確認された.このことから,毛包幹細胞から分泌されるEdnにより色素細胞の増殖が促進されるものと考えられた.そこで,この仮説を確かめるため,このトランスジェニックマウスにEdnrBの拮抗阻害剤であるBQ788を投与したところ色素細胞の異所的な増殖が抑えられた.また,このトランスジェニックマウスの上皮細胞を使った共培養実験において,BQ788の培地への添加,または,色素細胞に対するsiRNAによるEdnrBのノックダウンにより,色素細胞の増殖が抑えられた.このことから,Wntシグナル経路が活性化すると毛包幹細胞からEdnが分泌され,これにより色素幹細胞の分裂が促進されることがわかった.
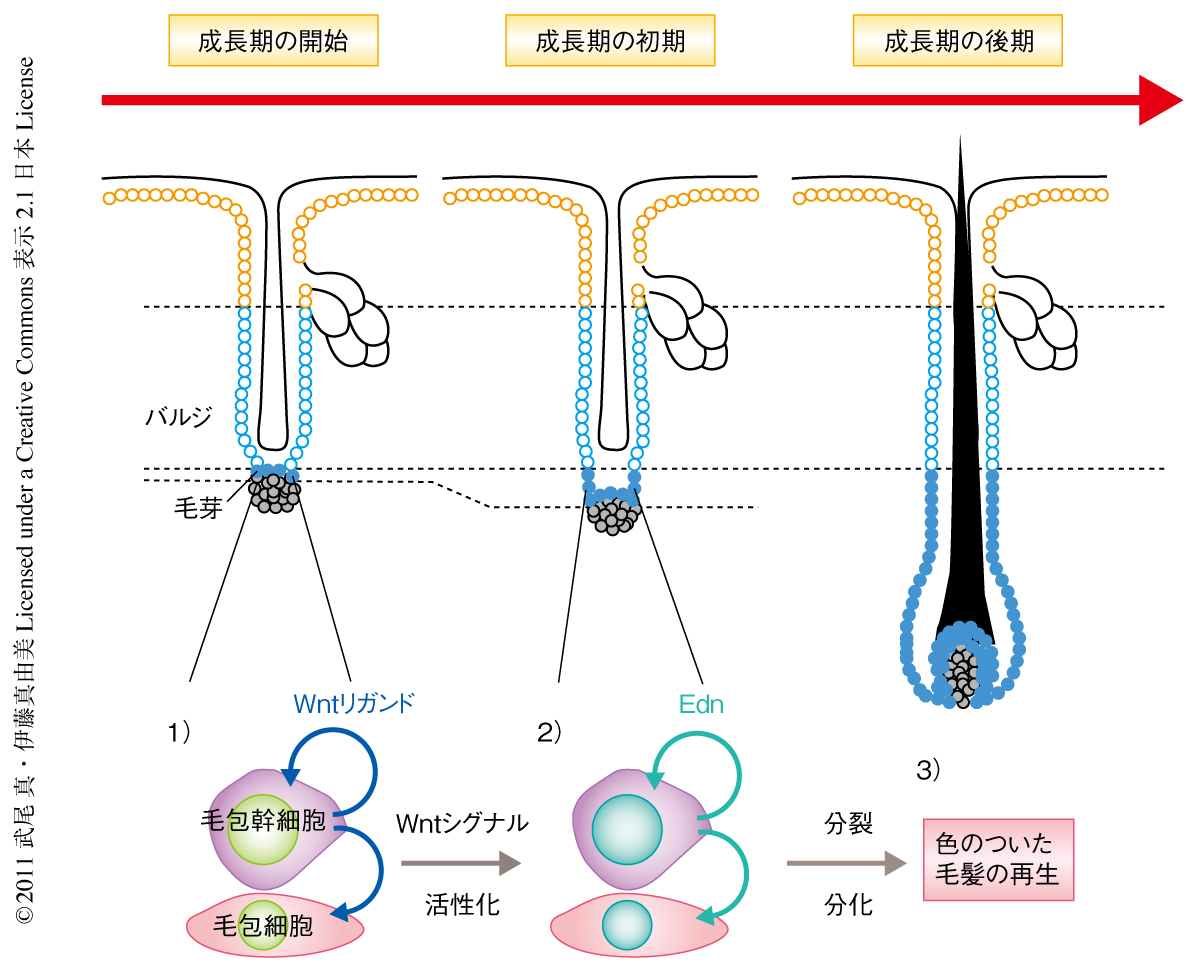
今回の研究により,1)毛包幹細胞からWntリガンドが分泌され,毛芽領域の毛包幹細胞と色素幹細胞とでWntシグナル経路が同時に活性化する,2)毛包幹細胞から分泌されたEdnにより色素幹細胞が増殖する,3)自らのWntシグナル経路の活性化により色素幹細胞が分化し毛包細胞から分化した毛髪に色素をわたす,という段階をへて色のついた毛髪が再生する分子機構が明らかになった(図2).機能的に完全な器官を再生するためには異なる種類の幹細胞や子孫細胞が時間的および空間的に協調してはたらくことが重要である.数種類の幹細胞を含み生涯をつうじて生理的な再生をくり返す哺乳類の毛包は,哺乳類の器官再生における幹細胞-幹細胞相互作用を研究するよいモデルである.今回,得られた知見および今後の研究により得られる結果が,哺乳類の器官再生のための新たな戦略と治療法の開発につながるものと期待される.
略歴:2008年 北海道大学大学院理学研究科 修了,同年より米国New York大学School of Medicine博士研究員.
研究テーマ:哺乳類の器官再生における(幹)細胞間の相互作用.
関心事:プラナリアからミミズをへてマウスやヒトにいたる再生現象一般.
伊藤 真由美(Mayumi Ito)
米国New York大学School of MedicineにてAssistant Professor.
© 2011 武尾 真・伊藤真由美 Licensed under CC 表示 2.1 日本
(米国New York大学School of Medicine,Department of Dermatology and Cell Biology)
email:武尾 真
DOI: 10.7875/first.author.2011.104
Coordinated activation of Wnt in epithelial and melanocyte stem cells initiates pigmented hair regeneration.
Piul Rabbani, Makoto Takeo, WeiChin Chou, Peggy Myung, Marcus Bosenberg, Lynda Chin, M. Mark Taketo, Mayumi Ito
Cell, 145, 941-955 (2011)
この論文に出現する遺伝子・タンパク質のUniprot ID
Wnt, βカテニン, カスパーゼ3(P70677), Wnt10b(P48614), Wnt7a(P24383), EdnrB(P48302), Endothelin1(P22387), Edn1(P22387), Edn2(P22387), Edn, Edn受容体B(P48302)
要 約
哺乳類の毛髪には寿命があり,毛包バルジで毛包幹細胞と色素幹細胞とが同時に活性化することにより色のついた毛髪がくり返し再生する.しかし,2つの異なる幹細胞が同時に活性化し協調的に毛髪を再生する分子機構はわかっていなかった.今回,筆者らは,Wntシグナル経路に注目し,細胞特異的にWntシグナル経路を変更することによりその分子機構の解明を試みた.その結果,毛髪の再生が開始するとき毛包幹細胞からWntリガンドが分泌され,毛包幹細胞と色素幹細胞とでWntシグナル経路が同時に活性化することが明らかになった.これらの結果は,毛包の再生過程においてWntシグナルが毛包幹細胞と色素幹細胞との協調的な挙動を可能にする分子機構としてはたらいていることを示すばかりでなく,多くの細胞種が関係する哺乳類の器官再生に関する新たな知見をあたえてくれる.
はじめに
哺乳類の毛髪には寿命があり生涯をつうじて再生と脱毛とをくり返す(毛周期).1回の毛周期は,毛包の下部が再生し毛髪がつくりだされる成長期,毛髪の伸長が止まり毛包下部が収縮する退行期,および,休止期の3つの時期に分けられる(図1).毛包のバルジ領域および毛芽領域には毛包幹細胞と色素幹細胞がある1,2).成長期にこの2つの幹細胞が同時に活性化し,毛包幹細胞の子孫細胞により毛包下部が再生され毛球部がつくられる.同時に,色素幹細胞の子孫細胞が毛球部へと移動し色素細胞へと分化する2).毛球部では毛包幹細胞の子孫細胞が毛髪に分化する過程で色素細胞によりつくられた色素を取り込み色のついた毛髪が再生する.このように,成長期の毛包の再生過程において毛包幹細胞と色素幹細胞とが同時に活性化し協調的に色のついた毛髪を再生させることは古くから知られていたが,その分子機構はわかっていなかった.
これまでに,毛包の再生過程において毛包幹細胞の活性化にかかわる分子機構の研究が精力的に行われ,Wntシグナル経路が重要な機構のひとつであることが示されている3,4).また,色素細胞の発生にはWntシグナルが必要であること5,6),培養細胞ではWntシグナルが色素細胞の分化を誘導することから7),色素幹細胞の分化にもWntシグナルが重要な役割をはたしているものと考えられる.しかしながら,成体の色素幹細胞に関しては幹細胞の維持に注目した研究は行われているが8),活性化の機構はほとんどわかっていない.そのため,成体の色素幹細胞におけるWntシグナルのはたらきはわかっていない.そこで筆者らは,毛包幹細胞または色素幹細胞に特異的な遺伝子プロモーターを使い,細胞特異的にWntシグナルを抑制または恒常的に活性化させ,成体の色素幹細胞の活性化にWntシグナルが必要であるかどうかを明らかにしようと試みた.また,毛包が再生されるときにどのようにして細胞系譜の異なる2種類の幹細胞が同時に活性化して分裂および分化するのか,その分子機構の解明も試みた.この研究により得られた結果は,単に毛包幹細胞と色素幹細胞とが協調的にはたらくための分子機構を明らかにするだけでなく,哺乳類の成体における多くの細胞種を含む複雑な器官の再生を可能にするヒントをあたえてくれるものと期待される.
1.毛包成長期の毛芽領域において毛包幹細胞と色素幹細胞とでWntが協調的に活性化する
毛包が再生するとき,色素幹細胞ではWntシグナル経路が活性化しているのだろうか? Wntシグナル経路が活性化するとβカテニンが核へと移行する.この核に局在したβカテニンをWntシグナル経路の活性化の指標とし,マウスの毛包において成長期に色素幹細胞ではWntシグナル経路が活性化しているのかどうか調べた.休止期には毛包幹細胞でも色素幹細胞でも核におけるβカテニンの発現はみられなかったが,成長期がはじまると毛芽領域において毛包幹細胞と色素幹細胞の両方でWntシグナル経路の活性化が同時に観察された.成長期が進んで毛球部がつくられるとWntシグナル経路の活性化は毛球部の毛包細胞と色素細胞だけにみられるようになった.毛周期をつうじてバルジ領域の色素幹細胞においてWntシグナル経路の活性化することはなかった.この結果から,毛包が再生するときに毛芽領域の色素幹細胞においてWntシグナルが活性化することが明らかになった.また,成長期でのWntシグナル経路の活性化がヒトの頭髪でもみられたことから,マウスとヒトの毛包の再生では同じ分子機構のはたらいていることが考えられた.
2.色素幹細胞におけるWntシグナルの恒常的な活性化は色素細胞の分化と早期の白髪をひき起こす
Wntシグナル経路の活性化が色素細胞の分化に必要かどうかを調べるため,休止期の色素幹細胞を含むすべての色素細胞においてWntシグナル経路を活性化しつづけるトランスジェニックマウスを作製した.このトランスジェニックマウスでは通常はWntシグナル経路の活性化していないバルジ領域の色素幹細胞でも色素細胞の分化マーカーが発現し,異所的に色素もつくっていた.一方で,普段からWntシグナル経路の活性化している毛球部の色素細胞では分化マーカーの発現と色素の産生について対照マウスとの違いはみられなかった.このことから,Wntシグナルの活性化は色素幹細胞の分化を誘導するのに十分であることが明らかになった.
おもしろいことに,このトランスジェニックマウスでは毛周期が進むにつれて白髪が増加し,4回目の毛周期ではすべてのトランスジェニックマウスで全体の20%以上が白髪になった.一方,対照マウスでは白髪の増加はみられなかった.これまで,バルジ領域の色素幹細胞が活性化することにより自己複製能力が低下し,最終的には色素幹細胞がなくなり白髪になることが報告されている9).そこで,このトランスジェニックマウスでも同じ理由で白髪が増加しているのかどうか調べた.その結果,トランスジェニックマウスでは毛周期が進むにつれてバルジ領域の色素幹細胞が減少していた.これらの結果から,恒常的なWntシグナル経路の活性化は色素幹細胞の分化をひき起こし,自己複製能を低下させ色素幹細胞の減少とそれにともなう白髪をひき起こすことが明らかになった.
3.βカテニンは色素幹細胞の分化に必要である
Wntシグナル経路の恒常的な活性化は色素幹細胞の分化の誘導に十分であることはわかったが,それでは,通常の毛包の再生はどうだろうか? 通常の毛包の再生におけるWntシグナルの役割を調べるため別のトランスジェニックマウスを作製した.このトランスジェニックマウスではタモキシフェンを投与することで色素細胞に特異的にWntシグナル経路の構成タンパク質のひとつであるβカテニンをコードする遺伝子をゲノムから取り除くことができる.βカテニンのなくなった細胞ではWntシグナル経路の活性化は起こらない.このトランスジェニックマウスにおいてβカテニンを取り除くと細胞周期の進行にともない白髪が有意に増加した.また,白髪の毛包には毛球部の色素細胞の数が少なく,色素細胞での分化マーカーの発現もみられなかった.一方,バルジ領域では色素幹細胞の数と分化状態に違いはみられなかった.
このトランスジェニックマウスの毛球部において色素細胞が減少する原因として,色素細胞が分裂できない可能性と色素細胞が死滅する可能性の2つが考えられた.これらの可能性を確かめるため,成長期での細胞分裂の頻度をBrdUの取り込み率を指標に比較したところ,トランスジェニックマウスではBrdUを取り込む色素細胞の割合が対照マウスに比べ約半分にまで減少していた.一方,細胞死のマーカーである活性化型カスパーゼ3はトランスジェニックマウスと対照マウスの両方において検出できなかった.これらの結果から,通常の毛包の再生においても色素幹細胞の増殖と分化にWntシグナル経路の活性化の必要であることが明らかになった.さきに述べたように,毛包幹細胞の増殖と分化にもWntシグナル経路がかかわっていた.また,毛包が再生するとき毛芽領域では毛包幹細胞と色素幹細胞で同時にWntシグナル経路が活性化した.これらのことから,Wntシグナル経路が2つの幹細胞が協調的に毛髪を再生するための分子機構としてはたらいているではないかと予想された.
4.毛包幹細胞から分泌されるWntリガンドは色素幹細胞のWntを活性化しうる
Wntシグナル経路はWntリガンドが受容体に結合することで活性化する.色素幹細胞を活性化するWntリガンドについて調べるため,神経堤から分化したすべての色素細胞がYFPを発現するトランスジェニックマウスから蛍光セルソーターにより色素幹細胞を単離し,すべてのWntリガンドの発現量の変化を調べた.しかしながら,成長期を誘導する前後でWntリガンドの発現の上昇は検出されなかった.
自らWntリガンドをつくりださずに,色素幹細胞はいったいどのようにWntシグナル経路を活性化しているのだろうか? 幹細胞説によると,幹細胞は内因的な要因とニッチからの外因的な要因により挙動が制御されている.そこで,バルジ領域および毛芽領域において色素幹細胞のとなりに存在する毛包幹細胞に注目した.まず,毛包幹細胞においてGFPを発現するトランスジェニックマウスから毛包幹細胞を単離し,リアルタイムPCR法によりWntリガンドの発現量を調べた.この結果,成長期を誘導した毛包幹細胞においていくつかのWntリガンドの発現量の増加することが確認された.また,in situハイブリダイゼーション法によりWntリガンドを発現している細胞は毛芽領域の毛包幹細胞であることも確認された.
つぎに,上皮細胞から分泌されたWntリガンドにより色素細胞のWntシグナル経路が活性化するかどうかを調べるため,Wnt10bの発現ベクターを導入した上皮細胞と色素細胞との共培養実験を行ったところ,色素細胞のWntシグナル経路の活性が約5倍まで増加した.さらに,生体においても外部からのWntリガンドにより色素幹細胞は活性化するかどうかを調べるためWntリガンドの皮内注入実験を行った.この実験には毛包幹細胞のWntシグナル経路の活性化による2次的な効果を排除するため毛包細胞に特異的にβカテニンを欠失させたトランスジェニックマウスを使った.このトランスジェニックマウスではWnt7aの注入により色素幹細胞においてWntシグナル経路の活性化がみられた.また,色素細胞の分化マーカーの発現も確認された.これらの結果から,毛包が再生するとき毛包幹細胞から分泌されるWntリガンドにより,毛包幹細胞と色素幹細胞とでWntシグナル経路が同時に活性化されることが示唆された.
5.上皮幹細胞での異所的なWntシグナルの活性化は色素細胞の増殖を促進する
毛包の再生過程において毛包幹細胞と色素幹細胞でWntシグナル経路が同時に活性化することにはどんな意味があるのだろうか? 毛包幹細胞でのWntシグナル経路の活性化が色素幹細胞の挙動にどのような影響をあたえるのか調べるため,バルジ領域を含むすべての毛包幹細胞においてWntシグナル経路を活性化しつづけるトランスジェニックマウスを作製した.このトランスジェニックマウスではバルジ領域にWntシグナル経路の活性化した肥大部が形成され,その領域で多くの色素細胞が観察された.また,バルジ領域の色素細胞の数も有意に増加していた.同様の結果は,すべての上皮基底細胞においてWntシグナル経路の活性化しつづける別のトランスジェニックマウスでも観察された.通常,マウスの皮膚に色素細胞はないが,おもしろいことに,このトランスジェニックマウスではバルジ領域だけではなく表皮にも多数の色素細胞がみられた.これらの結果は,毛包細胞でのWntシグナルの活性化は色素細胞の分裂を促進することを示していた.
6.色素細胞の増殖はEdnrBシグナルを介して行われる
毛包幹細胞でのWntシグナル経路の活性化がどのようにして色素細胞の増殖を促進するのか調べるため,バルジ領域を含むすべての毛包幹細胞においてWntシグナル経路を活性化しつづけるさきのトランスジェニックマウスと対照マウスとで発現している遺伝子を比較した.全層皮膚を使ったマイクロアレイ法による解析の結果,Wntシグナルの直接のターゲットであり色素細胞増殖因子と考えられるEndothelin1(Edn1)の発現がトランスジェニックマウスにおいて有意に増加していることが明らかになった.
全層皮膚には毛包幹細胞と色素幹細胞のほかにも多くの細胞が含まれる.そこで,このトランスジェニックマウスから蛍光セルソーターにより単離した上皮幹細胞と色素幹細胞を使いリアルタイムPCR法による解析を行った.その結果,毛包の再生の初期に毛包幹細胞においてEdn1とEdn2の発現が,色素幹細胞においてすべてのEdnの受容体であるEdn受容体B(EdnrB)の発現が増加することが確認された.このことから,毛包幹細胞から分泌されるEdnにより色素細胞の増殖が促進されるものと考えられた.そこで,この仮説を確かめるため,このトランスジェニックマウスにEdnrBの拮抗阻害剤であるBQ788を投与したところ色素細胞の異所的な増殖が抑えられた.また,このトランスジェニックマウスの上皮細胞を使った共培養実験において,BQ788の培地への添加,または,色素細胞に対するsiRNAによるEdnrBのノックダウンにより,色素細胞の増殖が抑えられた.このことから,Wntシグナル経路が活性化すると毛包幹細胞からEdnが分泌され,これにより色素幹細胞の分裂が促進されることがわかった.
おわりに
今回の研究により,1)毛包幹細胞からWntリガンドが分泌され,毛芽領域の毛包幹細胞と色素幹細胞とでWntシグナル経路が同時に活性化する,2)毛包幹細胞から分泌されたEdnにより色素幹細胞が増殖する,3)自らのWntシグナル経路の活性化により色素幹細胞が分化し毛包細胞から分化した毛髪に色素をわたす,という段階をへて色のついた毛髪が再生する分子機構が明らかになった(図2).機能的に完全な器官を再生するためには異なる種類の幹細胞や子孫細胞が時間的および空間的に協調してはたらくことが重要である.数種類の幹細胞を含み生涯をつうじて生理的な再生をくり返す哺乳類の毛包は,哺乳類の器官再生における幹細胞-幹細胞相互作用を研究するよいモデルである.今回,得られた知見および今後の研究により得られる結果が,哺乳類の器官再生のための新たな戦略と治療法の開発につながるものと期待される.
文 献
- Liu, Y., Lyle, S., Yang, Z. et al.: Keratin 15 promoter targets putative epithelial stem cells in the HF bulge. J. Invest. Dermatol., 121, 963-968 (2003)[PubMed]
- Nishimura, E. K., Jordan, S. A., Oshima, H. et al.: Dominant role of the niche in melanocyte stem-cell fate determination. Nature, 416, 854-860 (2002)[PubMed]
- Lowry, W. E., Blanpain, C., Nowak, J. A. et al.: Defining the impact of β-catenin/Tcf transactivation on epithelial stem cells. Genes Dev., 19, 1596-1611 (2005)[PubMed]
- Van Mater, D., Kolligs, F. T., Dlugosz, A. A. et al.: Transient activation ofβ-catenin signaling in cutaneous keratinocytes is sufficient to trigger the active growth phase of the hair cycle in mice. Genes Dev., 17, 1219-1224 (2003)[PubMed]
- Ikeya, M., Lee, S. M., Johnson, J. E. et al.: Wnt signalling required for expansion of neural crest and CNS progenitors. Nature, 389, 966-970 (1997)[PubMed]
- Hari, L., Brault, V., Kleber, M. et al.: Lineage-specific requirements of β-catenin in neural crest development. J. Cell Biol., 159, 867-880 (2002)[PubMed]
- Takeda, K., Yasumoto, K., Takada, R. et al.: Induction of melanocyte-specific microphthalmia-associated transcription factor by Wnt-3a. J. Biol. Chem., 275, 14013-14016 (2000)[PubMed]
- Nishimura, E. K., Suzuki, M., Igras, V. et al.: Key roles for transforming growth factor β in melanocyte stem cell maintenance. Cell Stem Cell, 6, 130-140 (2010)[PubMed]
- Nishimura, E. K., Granter, S. R. & Fisher, D. E.: Mechanisms of hair graying: incomplete melanocyte stem cell maintenance in the niche. Science, 307, 720-724 (2005)[PubMed]
著者プロフィール
略歴:2008年 北海道大学大学院理学研究科 修了,同年より米国New York大学School of Medicine博士研究員.
研究テーマ:哺乳類の器官再生における(幹)細胞間の相互作用.
関心事:プラナリアからミミズをへてマウスやヒトにいたる再生現象一般.
伊藤 真由美(Mayumi Ito)
米国New York大学School of MedicineにてAssistant Professor.
© 2011 武尾 真・伊藤真由美 Licensed under CC 表示 2.1 日本
