siRNAはストレスをうけた植物においてトランスポゾンの子孫への転移を抑制する
伊藤 秀臣
(北海道大学大学院理学研究院 生物科学部門形態機能学)
email:伊藤秀臣
DOI: 10.7875/first.author.2011.059
An siRNA pathway prevents transgenerational retrotransposition in plants subjected to stress.
Hidetaka Ito, Hervé Gaubert, Etienne Bucher, Marie Mirouze, Isabelle Vaillant, Jerzy Paszkowski
Nature, 472, 115-119 (2011)
環境の変化は生物の進化にとって受動的なものなのか,それとも,能動的なものなのか,長年のあいだ討論されてきた.筆者らは,環境の変化が植物の適応進化に積極的な影響力をもつことをはじめて実証した.多くの生物のゲノムにはさまざまなトランスポゾンが存在している.しかしながら,そのほとんどはすでに転移機能を喪失しているか活性が抑制された状態にある.筆者らは,植物に環境ストレスである高温ストレスをあたえるとあるグループのトランスポゾンが活性化することを発見した.このトランスポゾンには転写活性のみられるのみならず,RNA干渉を担う遺伝子の変異体では子孫へ転移および増殖の起こることがわかった.さらに,このトランスポゾンの転移先の近傍の遺伝子の発現量が変化することもみつけた.これらのことは,トランスポゾンの転移によりひき起こされる遺伝子の発現変異集団のなかからストレス耐性植物の誕生する可能性を示唆した.
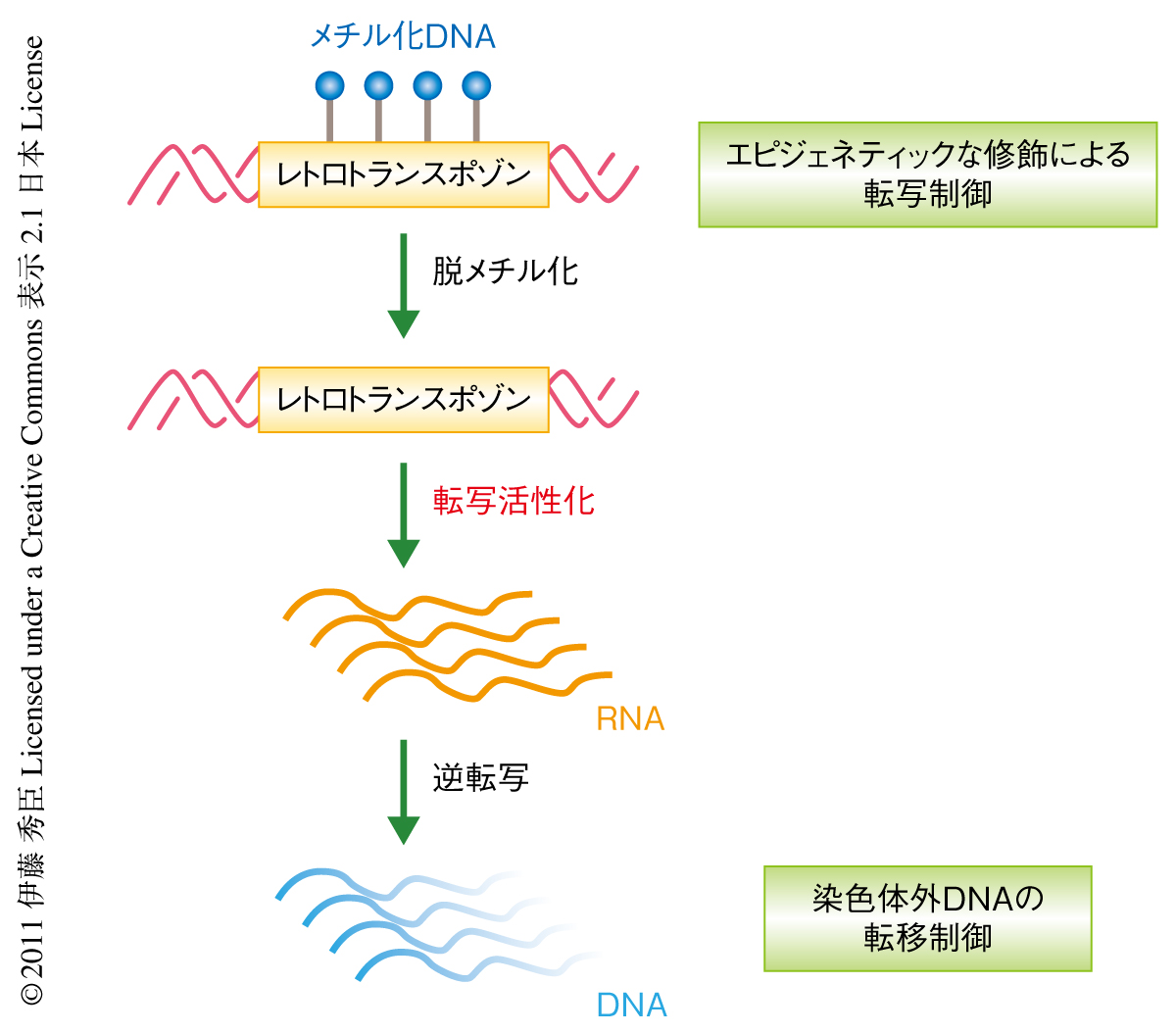
近年,シロイヌナズナにおけるトランスポゾンの制御にはDNAのメチル化などのエピジェネティックな機構のかかわっていることが報告されてきた1,2).また,植物には植物特異的な24塩基のsiRNA(small interfering RNA)の形成にかかわる遺伝子が備わっており,これはウイルスなど外来性の因子からの防御機構として進化してきたと考えられているRNA干渉(RNA interference:RNAi)を担っている.しかしながら近年まで,siRNAの形成を担う遺伝子の変異体において転移のみられるトランスポゾンの報告はなく,シロイヌナズナにおいてRNA干渉とトランスポゾンの転移制御との直接の関係は明らかにされていなかった.2009年,筆者らの所属していた研究グループは,Ty1/copia型のレトロトランスポゾンEvadeのDNA配列が低メチル化状態で転写活性を示した場合,植物に特異的なRNAポリメラーゼであるRNAポリメラーゼIV/Vの変異体と組み合わせることで転移の起こることをみつけた3).これにより,レトロトランスポゾンの制御機構として,転写活性を抑えるため宿主が獲得したDNAのメチル化機構と転写後制御としてのRNAポリメラーゼIV/Vの役割が示唆されていた(図1).
今回の筆者らの研究結果は,siRNAがトランスポゾンの転写後制御に重要な役割をはたしていることをあらためてサポートすることとなった.筆者らの研究グループは,温度の変化(21℃から37℃へ)に対応して変化する遺伝子をマイクロアレイにより網羅的に解析した.その結果,ほとんどの遺伝子の発現量は一時的に変化するものの,処理ののち24時間以内にはもとの状態にもどることがわかった4).しかし例外的に,あるグループに属するTy1/copia型のレトロトランスポゾンは処理2日後でも高い発現量を持続していた.このグループには8コピーのトランスポゾン全長配列が含まれ,そのうち3コピーはLTR(long terminal repeat,長い末端反復配列)の塩基配列が100%の相同性を示した.筆者らは,植物にとって高温ストレスとなる37℃の処理で活性化するこのトランスポゾンを“ONSEN”と命名した.
現在までに報告されている代表的なトランスポゾンのエピジェネティックな制御機構としてDNAのメチル化があげられる5,6).ONSENの転写活性制御に対するDNAのメチル化の関与を調べるため,DNAメチル化阻害剤を用いた処理とメチル化が低下するシロイヌナズナ変異体ddm1を用いた解析を行った結果,予想に反してONSENの活性化はみられなかった.このことから,DNAの脱メチルだけではONSENの活性化に十分ではないことがわかった.外来性の因子やトランスポゾンの制御機構として,近年,RNA干渉に関する研究がさかんに行われている.とくに,植物には24塩基からなるsiRNAを介した制御機構が備わっており,その経路にはたらく植物に特異的なRNAポリメラーゼIV/Vが存在している7,8).そこで,シロイヌナズナのsiRNAの形成に関与する遺伝子として,RNAポリメラーゼIV/V,RNA依存性RNAポリメラーゼ2,Dicer-like 3の3つの変異体を用いた転写活性の解析を行った.結果は,どの変異体を用いた場合でもONSENは不活性なままであった.このことから,ONSENの転写活性には高温(37℃)というストレス条件が必要なようであり,ONSEN遺伝子の発現を直接に制御している役者はいまだ同定されていない.興味深い点は,高温処理ののちのONSENの転写量は野生型に比べsiRNAの形成に関与する遺伝子の変異体で増加していたことである.このことから,ONSENの転写にはsiRNAを介した制御が行われているものと考えられた.
レトロトランスポゾンの複製的な転移では,逆転写により自己の配列を転写したRNAから染色体外DNAコピーがつくられる.そのため,この染色体外DNAコピーを検出することで実際にトランスポゾンに転移活性があるのかどうか調べることができる.発芽後1週間のシロイヌナズナの種苗に高温処理を施し,ONSENの染色体外DNAコピーの存在をサザンブロット法により調べてみると鎖状と環状の2つの形状のものと思われるDNAが検出された.鎖状のDNAは転移能力をもつと考えられ,環状のものはレトロトランスポゾンの複製にみられる副産物と考えられた.また,高温処理によるONSENの発現量と同様に,野生型に比べsiRNAの形成に関与する遺伝子の変異体ではより多くの染色体外DNAコピーのつくられていることが明らかになった.リアルタイムPCR法を用いて染色体外DNAコピーの蓄積を時系列をおって観察した結果,ONSENの染色体外DNAコピーの量は高温処理6時間あたりから増加しはじめ,高温処理の終了ののちはしだいに減少し,20日後には高温処理をはじめるまえの量と変わらないまでにもどった.高温処理ののちに一時的ではあるがONSENの染色体外DNAコピーの生産されたことから体細胞への転移が期待された.そのため,トランスポゾンディスプレイ法やサザンブロット法を用いて体細胞での転移あるいは挿入を調べたが新しい転移は確認できなかった.このことは,体細胞への挿入によるコピー数の増加はほとんどみられない,もしくは,ごく一部の体細胞組織でのみ転移の起こることを意味し,転移頻度はかなり低いことを示唆した.
ここまでの結果では,ONSENの転写量および染色体外DNAコピーの量は高温処理ののち時間とともに減少し,その世代での転移もほとんど起こらないようであったことから,ONSENに焦点をあてた研究へのモチベーションは下がってしまっていた.しかしながら,環境ストレスがトランスポゾンの転移に影響をあたえるという仮説を検証するためには転移の観察が必要不可欠であった.そこで,高温処理を施した植物の子孫での転移を調べてみることにした.以前から,トランスポゾンの次世代への転移は配偶子の形成時期にsiRNAにより抑制されるという仮説があったが9,10),実際にこれを証明した例はなかった.そこで,野生型およびsiRNAの形成に関与する遺伝子の変異体を用いてその子孫での転移を解析した.その結果,驚くべきことに,siRNAの形成に関与する遺伝子の変異体に高温処理を施した個体の子孫でONSENの転移が観察された(図2).
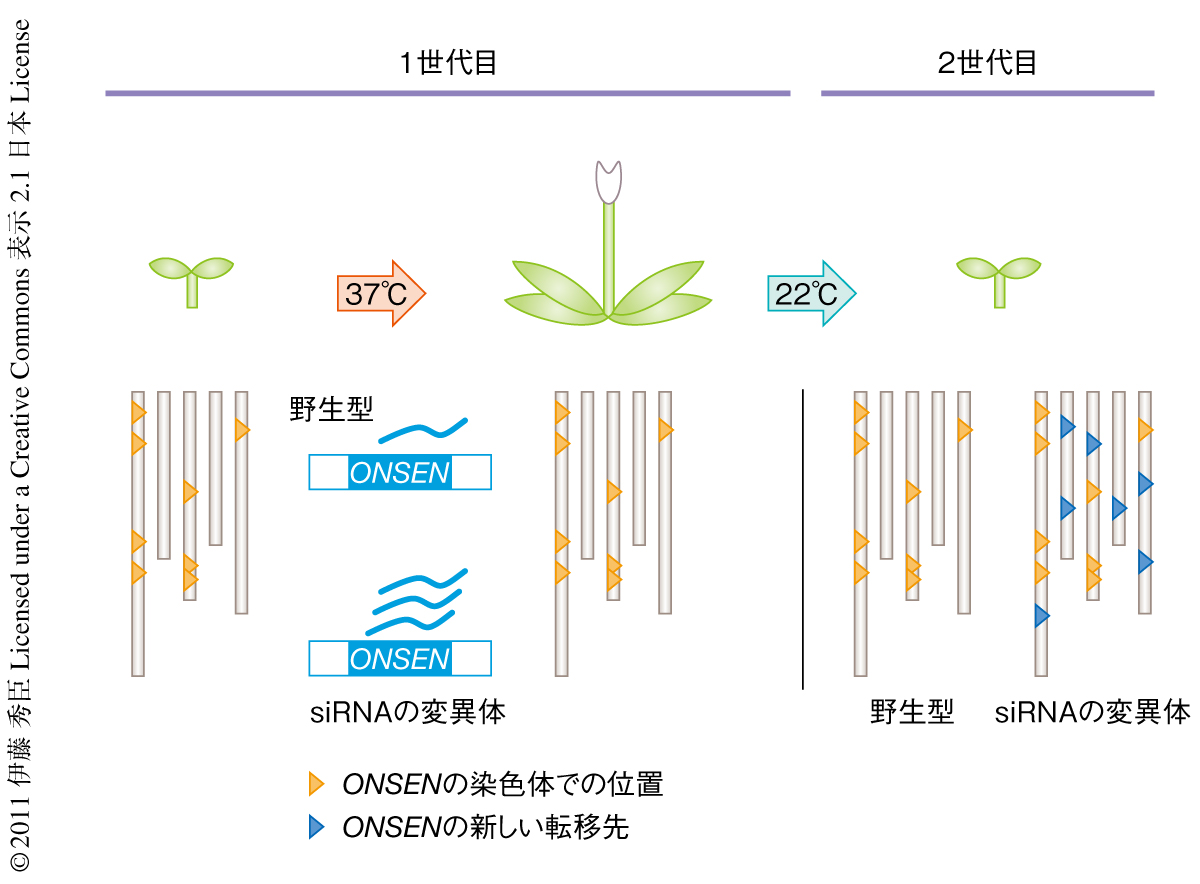
さらに興味深い点は,子の個体それぞれの挿入パターンが異なっていたことであった.この観察結果からいくつかの可能性が考えられた.1つ目はONSENの転移が配偶子の形成まえの花組織の体細胞で起こったこと,2つ目は配偶子で起こったこと,3つ目は受精ののちに起こったこと,である.これらの仮説を検証すべく,ひとつの個体に咲く別々の花から採取した種子のあいだでの挿入パターンを検証した.その結果,別々の花から採取した子孫のあいだでは転移先に共通性はなく,また,同じ花から採取した子孫のあいだでは共通なパターンをもつものと個体間で差のあるものの両方を含むという結果になった.つまり,ONSENの転移は親植物が配偶子を形成する以前の花組織にある体細胞で起こったことになった.このことは,以前に考えられていたsiRNAが配偶子の形成のときにトランスポゾンの転移を抑制するという仮説をくつがえすものであり,体細胞組織においてもsiRNAが転移制御に重要なはたらきをしていることを示していた.
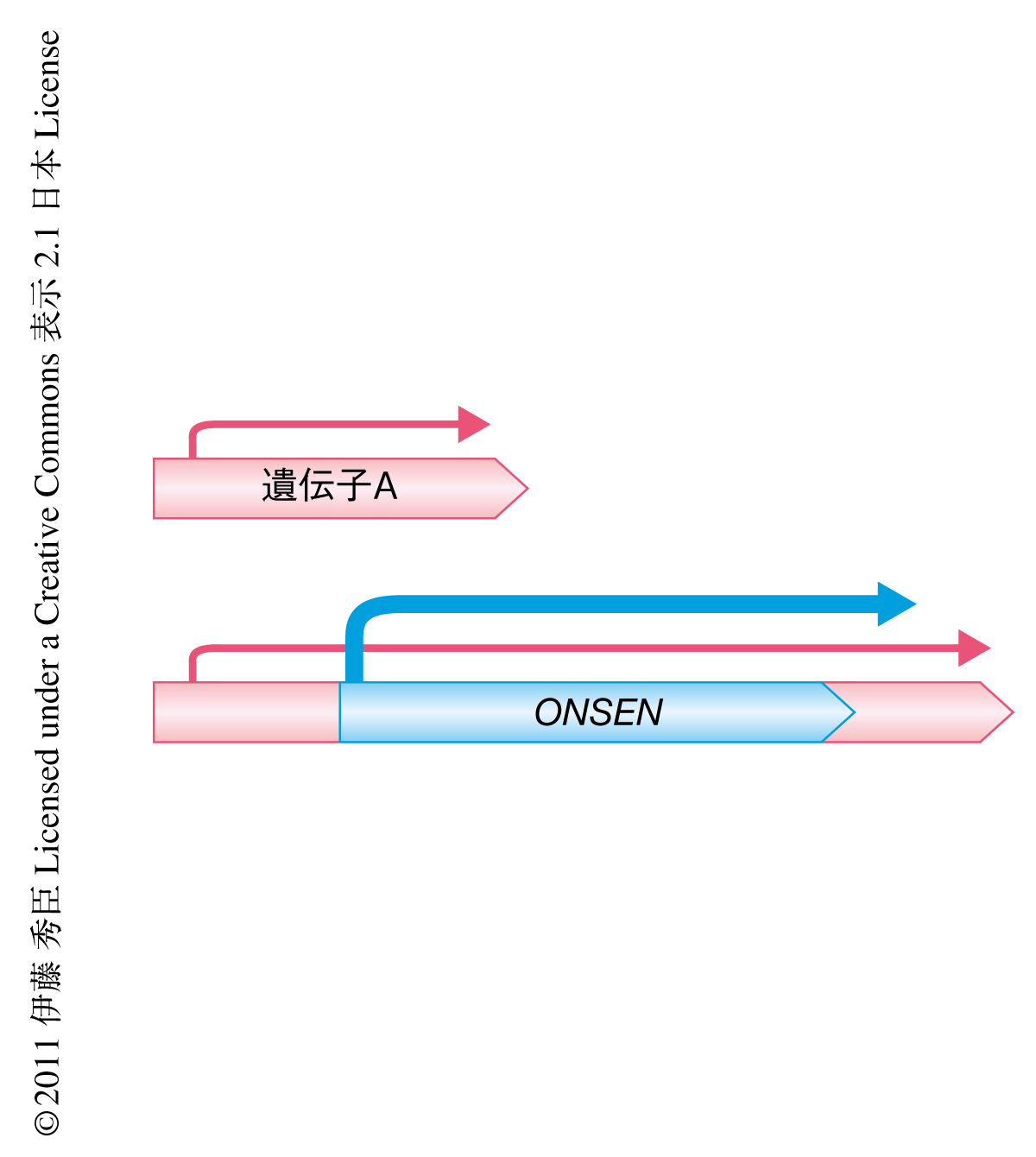
さまざまな生物種に存在するトランスポゾンは染色体のヘテロクロマチンに局在するものや遺伝子領域の近傍のユークロマチンに存在するものなど多様である.ONSENの転移先には何か共通性があるのか調べるため,転移先の領域のDNA配列を解読した.11個の挿入領域を調べた結果,興味深いことに,すべてが遺伝子領域に挿入されており,しかも,そのうち10個はエクソン領域に飛び込んでいた.このことは,ONSENは選り好んで遺伝子に飛び込み,その結果,転移先の遺伝子の発現に影響のでる可能性の高いことを意味していた(図3).
それでは,現存するシロイヌナズナのゲノムではONSENによる遺伝子の発現制御が起こっているのだろうか? それを調べるため近交系統を解析した.シロイヌナズナには世界各地に分布しそれぞれの環境に適応した近交系統が確立されている.そのなかで標準系統として用いられているColumbiaではONSENが熱感受性遺伝子の内部に挿入されていた.その遺伝子にONSENの含まれていない系統(Zurich)と比較すると,Columbia系統に高温処理を施したもののほうが遺伝子の発現量は上昇していた.このことは,ONSENの挿入により宿主ゲノムの遺伝子発現量の制御されていることを意味していた.つぎに,実際にsiRNAの形成に関与する遺伝子の変異体に高温処理を施しONSENの転移が観察された子孫を用いて転移先の領域の遺伝子発現量を解析した.その結果,期待どおり,ONSEN遺伝子自体の活性化と同時に転移先の遺伝子の発現量も上昇していることがわかった.
今回,発見したトランスポゾンONSENは高温により活性化し宿主子孫の遺伝子発現に影響をあたえることがわかった.トランスポゾンは宿主のゲノム構造を変える非常に有力なものであるがゆえ,宿主としてはこれを抑え込む必要がある.その戦略のひとつとしてエピジェネティックな修飾による転写活性の制御機構を獲得したのであり,また,もうひとつの戦略としてRNA干渉による転移制御があるのであろう.この二重のロックを解除することで高頻度の転移が起こり宿主のゲノム構造の変化および遺伝子発現の変化が起こる.宿主にとって有害とも思われるこれらの変化であるが,一方では,植物がストレスにさらされた際に環境適応形質を獲得するひとつのしくみであると考えることもできるのではないだろうか.
略歴:2004年 京都大学大学院農学研究科 修了,同年 国立遺伝学研究所 研究員,2007年 スイスGeneva大学 博士研究員を経て,2010年より北海道大学大学院理学研究院 助教.
研究テーマ:環境ストレスとゲノム進化.
抱負:自分の研究成果が将来の科学の礎となれればと思います.
© 2011 伊藤 秀臣 Licensed under CC 表示 2.1 日本
(北海道大学大学院理学研究院 生物科学部門形態機能学)
email:伊藤秀臣
DOI: 10.7875/first.author.2011.059
An siRNA pathway prevents transgenerational retrotransposition in plants subjected to stress.
Hidetaka Ito, Hervé Gaubert, Etienne Bucher, Marie Mirouze, Isabelle Vaillant, Jerzy Paszkowski
Nature, 472, 115-119 (2011)
要 約
環境の変化は生物の進化にとって受動的なものなのか,それとも,能動的なものなのか,長年のあいだ討論されてきた.筆者らは,環境の変化が植物の適応進化に積極的な影響力をもつことをはじめて実証した.多くの生物のゲノムにはさまざまなトランスポゾンが存在している.しかしながら,そのほとんどはすでに転移機能を喪失しているか活性が抑制された状態にある.筆者らは,植物に環境ストレスである高温ストレスをあたえるとあるグループのトランスポゾンが活性化することを発見した.このトランスポゾンには転写活性のみられるのみならず,RNA干渉を担う遺伝子の変異体では子孫へ転移および増殖の起こることがわかった.さらに,このトランスポゾンの転移先の近傍の遺伝子の発現量が変化することもみつけた.これらのことは,トランスポゾンの転移によりひき起こされる遺伝子の発現変異集団のなかからストレス耐性植物の誕生する可能性を示唆した.
はじめに
近年,シロイヌナズナにおけるトランスポゾンの制御にはDNAのメチル化などのエピジェネティックな機構のかかわっていることが報告されてきた1,2).また,植物には植物特異的な24塩基のsiRNA(small interfering RNA)の形成にかかわる遺伝子が備わっており,これはウイルスなど外来性の因子からの防御機構として進化してきたと考えられているRNA干渉(RNA interference:RNAi)を担っている.しかしながら近年まで,siRNAの形成を担う遺伝子の変異体において転移のみられるトランスポゾンの報告はなく,シロイヌナズナにおいてRNA干渉とトランスポゾンの転移制御との直接の関係は明らかにされていなかった.2009年,筆者らの所属していた研究グループは,Ty1/copia型のレトロトランスポゾンEvadeのDNA配列が低メチル化状態で転写活性を示した場合,植物に特異的なRNAポリメラーゼであるRNAポリメラーゼIV/Vの変異体と組み合わせることで転移の起こることをみつけた3).これにより,レトロトランスポゾンの制御機構として,転写活性を抑えるため宿主が獲得したDNAのメチル化機構と転写後制御としてのRNAポリメラーゼIV/Vの役割が示唆されていた(図1).
今回の筆者らの研究結果は,siRNAがトランスポゾンの転写後制御に重要な役割をはたしていることをあらためてサポートすることとなった.筆者らの研究グループは,温度の変化(21℃から37℃へ)に対応して変化する遺伝子をマイクロアレイにより網羅的に解析した.その結果,ほとんどの遺伝子の発現量は一時的に変化するものの,処理ののち24時間以内にはもとの状態にもどることがわかった4).しかし例外的に,あるグループに属するTy1/copia型のレトロトランスポゾンは処理2日後でも高い発現量を持続していた.このグループには8コピーのトランスポゾン全長配列が含まれ,そのうち3コピーはLTR(long terminal repeat,長い末端反復配列)の塩基配列が100%の相同性を示した.筆者らは,植物にとって高温ストレスとなる37℃の処理で活性化するこのトランスポゾンを“ONSEN”と命名した.
1.ONSENの転写活性
現在までに報告されている代表的なトランスポゾンのエピジェネティックな制御機構としてDNAのメチル化があげられる5,6).ONSENの転写活性制御に対するDNAのメチル化の関与を調べるため,DNAメチル化阻害剤を用いた処理とメチル化が低下するシロイヌナズナ変異体ddm1を用いた解析を行った結果,予想に反してONSENの活性化はみられなかった.このことから,DNAの脱メチルだけではONSENの活性化に十分ではないことがわかった.外来性の因子やトランスポゾンの制御機構として,近年,RNA干渉に関する研究がさかんに行われている.とくに,植物には24塩基からなるsiRNAを介した制御機構が備わっており,その経路にはたらく植物に特異的なRNAポリメラーゼIV/Vが存在している7,8).そこで,シロイヌナズナのsiRNAの形成に関与する遺伝子として,RNAポリメラーゼIV/V,RNA依存性RNAポリメラーゼ2,Dicer-like 3の3つの変異体を用いた転写活性の解析を行った.結果は,どの変異体を用いた場合でもONSENは不活性なままであった.このことから,ONSENの転写活性には高温(37℃)というストレス条件が必要なようであり,ONSEN遺伝子の発現を直接に制御している役者はいまだ同定されていない.興味深い点は,高温処理ののちのONSENの転写量は野生型に比べsiRNAの形成に関与する遺伝子の変異体で増加していたことである.このことから,ONSENの転写にはsiRNAを介した制御が行われているものと考えられた.
2.染色体外DNAコピーの合成と蓄積
レトロトランスポゾンの複製的な転移では,逆転写により自己の配列を転写したRNAから染色体外DNAコピーがつくられる.そのため,この染色体外DNAコピーを検出することで実際にトランスポゾンに転移活性があるのかどうか調べることができる.発芽後1週間のシロイヌナズナの種苗に高温処理を施し,ONSENの染色体外DNAコピーの存在をサザンブロット法により調べてみると鎖状と環状の2つの形状のものと思われるDNAが検出された.鎖状のDNAは転移能力をもつと考えられ,環状のものはレトロトランスポゾンの複製にみられる副産物と考えられた.また,高温処理によるONSENの発現量と同様に,野生型に比べsiRNAの形成に関与する遺伝子の変異体ではより多くの染色体外DNAコピーのつくられていることが明らかになった.リアルタイムPCR法を用いて染色体外DNAコピーの蓄積を時系列をおって観察した結果,ONSENの染色体外DNAコピーの量は高温処理6時間あたりから増加しはじめ,高温処理の終了ののちはしだいに減少し,20日後には高温処理をはじめるまえの量と変わらないまでにもどった.高温処理ののちに一時的ではあるがONSENの染色体外DNAコピーの生産されたことから体細胞への転移が期待された.そのため,トランスポゾンディスプレイ法やサザンブロット法を用いて体細胞での転移あるいは挿入を調べたが新しい転移は確認できなかった.このことは,体細胞への挿入によるコピー数の増加はほとんどみられない,もしくは,ごく一部の体細胞組織でのみ転移の起こることを意味し,転移頻度はかなり低いことを示唆した.
3.世代をこえた転移
ここまでの結果では,ONSENの転写量および染色体外DNAコピーの量は高温処理ののち時間とともに減少し,その世代での転移もほとんど起こらないようであったことから,ONSENに焦点をあてた研究へのモチベーションは下がってしまっていた.しかしながら,環境ストレスがトランスポゾンの転移に影響をあたえるという仮説を検証するためには転移の観察が必要不可欠であった.そこで,高温処理を施した植物の子孫での転移を調べてみることにした.以前から,トランスポゾンの次世代への転移は配偶子の形成時期にsiRNAにより抑制されるという仮説があったが9,10),実際にこれを証明した例はなかった.そこで,野生型およびsiRNAの形成に関与する遺伝子の変異体を用いてその子孫での転移を解析した.その結果,驚くべきことに,siRNAの形成に関与する遺伝子の変異体に高温処理を施した個体の子孫でONSENの転移が観察された(図2).
さらに興味深い点は,子の個体それぞれの挿入パターンが異なっていたことであった.この観察結果からいくつかの可能性が考えられた.1つ目はONSENの転移が配偶子の形成まえの花組織の体細胞で起こったこと,2つ目は配偶子で起こったこと,3つ目は受精ののちに起こったこと,である.これらの仮説を検証すべく,ひとつの個体に咲く別々の花から採取した種子のあいだでの挿入パターンを検証した.その結果,別々の花から採取した子孫のあいだでは転移先に共通性はなく,また,同じ花から採取した子孫のあいだでは共通なパターンをもつものと個体間で差のあるものの両方を含むという結果になった.つまり,ONSENの転移は親植物が配偶子を形成する以前の花組織にある体細胞で起こったことになった.このことは,以前に考えられていたsiRNAが配偶子の形成のときにトランスポゾンの転移を抑制するという仮説をくつがえすものであり,体細胞組織においてもsiRNAが転移制御に重要なはたらきをしていることを示していた.
4.転移先の特徴
さまざまな生物種に存在するトランスポゾンは染色体のヘテロクロマチンに局在するものや遺伝子領域の近傍のユークロマチンに存在するものなど多様である.ONSENの転移先には何か共通性があるのか調べるため,転移先の領域のDNA配列を解読した.11個の挿入領域を調べた結果,興味深いことに,すべてが遺伝子領域に挿入されており,しかも,そのうち10個はエクソン領域に飛び込んでいた.このことは,ONSENは選り好んで遺伝子に飛び込み,その結果,転移先の遺伝子の発現に影響のでる可能性の高いことを意味していた(図3).
それでは,現存するシロイヌナズナのゲノムではONSENによる遺伝子の発現制御が起こっているのだろうか? それを調べるため近交系統を解析した.シロイヌナズナには世界各地に分布しそれぞれの環境に適応した近交系統が確立されている.そのなかで標準系統として用いられているColumbiaではONSENが熱感受性遺伝子の内部に挿入されていた.その遺伝子にONSENの含まれていない系統(Zurich)と比較すると,Columbia系統に高温処理を施したもののほうが遺伝子の発現量は上昇していた.このことは,ONSENの挿入により宿主ゲノムの遺伝子発現量の制御されていることを意味していた.つぎに,実際にsiRNAの形成に関与する遺伝子の変異体に高温処理を施しONSENの転移が観察された子孫を用いて転移先の領域の遺伝子発現量を解析した.その結果,期待どおり,ONSEN遺伝子自体の活性化と同時に転移先の遺伝子の発現量も上昇していることがわかった.
おわりに
今回,発見したトランスポゾンONSENは高温により活性化し宿主子孫の遺伝子発現に影響をあたえることがわかった.トランスポゾンは宿主のゲノム構造を変える非常に有力なものであるがゆえ,宿主としてはこれを抑え込む必要がある.その戦略のひとつとしてエピジェネティックな修飾による転写活性の制御機構を獲得したのであり,また,もうひとつの戦略としてRNA干渉による転移制御があるのであろう.この二重のロックを解除することで高頻度の転移が起こり宿主のゲノム構造の変化および遺伝子発現の変化が起こる.宿主にとって有害とも思われるこれらの変化であるが,一方では,植物がストレスにさらされた際に環境適応形質を獲得するひとつのしくみであると考えることもできるのではないだろうか.
文 献
- Slotkin, R. K. & Martienssen, R.: Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet., 8, 272-285 (2007)[PubMed]
- Lisch, D.: Epigenetic regulation of transposable elements in plants. Annu. Rev. Plant Biol., 60, 43-66 (2008)[PubMed]
- Mirouze, M., Reinders, J., Bucher, E. et al.: Selective epigenetic control of retrotransposition in Arabidopsis. Nature, 461, 427-430 (2009)[PubMed]
- Tittel-Elmer, M., Bucher, E., Broger, L. et al.: Stress-induced activation of heterochromatic transcription. PLoS Genet., 6, e1001175 (2010)[PubMed]
- Miura, A., Yonebayashi, S., Watanabe, K. et al.: Mobilization of transposons by a mutation abolishing full DNA methylation in Arabidopsis. Nature, 411, 212-214 (2001)[PubMed]
- Tsukahara, S., Kobayashi, A., Kawabe, A. et al.: Bursts of retrotransposition reproduced in Arabidopsis. Nature, 461, 423-426 (2009)[PubMed]
- Herr, A. J., Jensen, M. B., Dalmay, T. et al.: RNA polymerase IV directs silencing of endogenous DNA. Science, 308, 118-120 (2005)[PubMed]
- Mosher, R. A., Schwach, F., Studholme, D. et al.: PolIVb influences RNA-directed DNA methylation independently of its role in siRNA biogenesis. Proc. Natl Acad. Sci. USA, 105, 3145-3150 (2008)[PubMed]
- Slotkin, R. K., Vaughn, M., Borges, F. et al.: Epigenetic reprogramming and small RNA silencing of transposable elements in pollen. Cell, 136, 461-472 (2009)[PubMed]
- Law, J. A. & Jacobsen, S. E.: Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet., 11, 204-220 (2010)[PubMed]
著者プロフィール
略歴:2004年 京都大学大学院農学研究科 修了,同年 国立遺伝学研究所 研究員,2007年 スイスGeneva大学 博士研究員を経て,2010年より北海道大学大学院理学研究院 助教.
研究テーマ:環境ストレスとゲノム進化.
抱負:自分の研究成果が将来の科学の礎となれればと思います.
© 2011 伊藤 秀臣 Licensed under CC 表示 2.1 日本
