アストロサイトからニューロンへの乳酸の供給は海馬に依存的な長期記憶の形成に必須である
鈴木章円・Cristina M. Alberini
(米国Mount Sinai School of Medicine,Department of Neuroscience)
email:鈴木章円
DOI: 10.7875/first.author.2011.056
Astrocyte-neuron lactate transport is required for long-term memory formation.
Akinobu Suzuki, Sarah A. Stern, Ozlem Bozdagi, George W. Huntley, Ruth H. Walker, Pierre J. Magistretti, Cristina M. Alberini
Cell, 144, 810-823 (2011)
脳神経系はニューロンとグリアから構成されているが,グリアの記憶形成に対する役割はほとんど明らかにされていない.この論文において筆者らは,記憶形成のときにグリアのひとつであるアストロサイトにおいてグリコーゲンから産生された乳酸のニューロンへの供給が長期記憶の形成に必須であることを明らかにした.グリコーゲン分解酵素阻害剤の海馬への直接の投与は長期記憶の形成,長期増強,さらに,記憶形成に関与するタンパク質の発現やリン酸化を阻害した.また,これらの阻害は乳酸の投与により回復することも発見された.さらに,乳酸のアストロサイトからの放出に関与するトランスポーターであるMCT1およびMCT4の発現を抑制した場合にも長期記憶の形成阻害が観察され,この阻害は乳酸の投与により回復した.一方,ニューロンへの乳酸の取り込みに関与するトランスポーターMCT2の発現を抑制した場合にも同様に記憶形成の阻害が観察されたが,乳酸の投与による回復は観察されなかったことから,アストロサイトからニューロンへの乳酸の輸送が記憶形成に必須であることが示された.
これまで,グリアのひとつであるアストロサイトはニューロンをサポートする役割をもつものと考えられていた1).しかしながら近年,アストロサイトが神経突起の伸長やニューロンのあいだのシグナル伝達に関与すること2),さらに,記憶の細胞モデルである長期増強(long-term potentiation:LTP)の制御にも関与することが報告され3),アストロサイトがより積極的にニューロンの活動を制御する可能性が示唆されている.さらに,記憶形成ののちの海馬においてアストロサイトの数が増加することや4),アストロサイトの機能を抑制すると記憶形成が阻害されるなど5),記憶の形成に対するアストロサイトの役割に関する研究も行われはじめている.したがって,記憶など脳の高次機能はニューロンとアストロサイトとのあいだの相互作用を介して制御されることが示唆されている.ニューロンとアストロサイトとの相互作用のひとつの分子機構としてニューロンへのエネルギーの供給があげられる.グルコースは脳活動を維持するためのエネルギー源のひとつであり,グルコーストランスポーターを介してニューロンやアストロサイトに輸送されグルタミン酸などの神経伝達物質の生合成に使われる6).一方で,ニューロンとは異なり,アストロサイトにはグリコーゲンが貯蔵され7),ニューロンのエネルギー源として利用されることが明らかになっている(図1)が8),記憶形成への役割は明らかになっていなかった.この研究において,筆者らは,アストロサイトにおけるグリコーゲン分解系やアストロサイトからニューロンへの乳酸の供給が長期記憶の形成に必須であることを見い出した.

まず,アストロサイトにおけるグリコーゲン分解系が記憶の形成に影響をあたえるかどうかを評価するため,グリコーゲン分解酵素の阻害剤であるDAB 9)(1,4-ジジオキシ-1,4-イミノ-d-アラビニトール)をラットの脳,記憶形成に重要な役割を担っている海馬に直接に投与し,受動的回避学習課題により短期および長期の記憶形成の能力を調べた.受動的回避学習課題とはラットの暗い場所を好む習性を利用して記憶能力を評価する行動学的な解析のひとつである.この解析において,明暗の2箱に分けた実験箱の明箱側にラットを入れると,ラットは暗いところを好むためすみやかに暗箱に入る.その直後にラットに電気ショックをあたえることで暗箱という“場所”と電気ショックによる“痛み”を関連づけて学習させる(トレーニング).一定時間の経過ののち同一の実験箱の明箱側にふたたびラットを入れると,以前の電気ショックによる体験と強い嫌悪感を想起し暗箱には入らなくなる.そこで,ラットが明箱から暗箱に移動するまでの時間を記録し(テスト),その反応時間の長短から記憶能力を評価する.暗箱に入るまでの時間が長いほど強く記憶していることになる.
トレーニングの15分前に海馬にグリコーゲン分解酵素阻害剤を投与しトレーニングの1時間後に記憶能力を評価したところ,グリコーゲン分解酵素阻害剤の投与群と溶媒を投与した対照群とのあいだに有意な差は観察されず,グリコーゲン分解酵素阻害剤の投与は短期記憶の形成には影響をあたえなかった.しかしながら,24時間後に記憶能力を評価した結果では投与群は対照群に比べ暗箱に入るまでの時間が有意に短く,長期記憶の形成能力の障害が観察されその阻害はトレーニングから1週間後でも観察された.また,トレーニングの直後にグリコーゲン分解酵素阻害剤を投与した場合も同様に記憶の形成阻害が観察されたが,トレーニングの24時間後に投与しさらに24時間後に記憶能力を評価した場合には記憶阻害は観察されなかった.以上の結果から,アストロサイトにおけるグリコーゲンの分解系は短期記憶の形成には必要とされないが,より多くのエネルギーを必要とするタンパク質合成をともなう長期記憶の形成には必須であること,さらに,トレーニング前後の比較的短い期間においてグリコーゲンの分解が必要であることが示唆された.
つぎに,グリコーゲン分解系によって産生されたどの基質が長期記憶の形成に関与しているのかを調べた.過去の培養細胞を用いた研究から,細胞外からの刺激によりグリコーゲンの分解が促されると乳酸が合成され,この乳酸がアストロサイトから細胞外へと放出されることが報告されていた10).そこで,トレーニングののちにおいてもグリコーゲンから乳酸が合成されこれが記憶形成に関与するのかどうかを調べるため,トレーニング前後のラットの海馬における細胞外の乳酸の濃度をin vivoマイクロダイアリシス法により測定した.その結果,トレーニングののちすみやかに乳酸の濃度は上昇し,さらに,トレーニングによる乳酸の濃度上昇はグリコーゲン分解酵素阻害剤の海馬への投与により抑制された.このことから,観察された乳酸の濃度上昇はグリコーゲンの分解によりもたらされていることが示唆された.そこで,乳酸の海馬への投与によりグリコーゲン分解酵素阻害剤による記憶障害は回復するかどうかを調べるため,グリコーゲン分解酵素阻害剤と乳酸とを海馬へ同時に投与し記憶能力を評価した.受動的回避学習課題により記憶形成への影響を観察したところ,グリコーゲン分解酵素阻害剤と乳酸(実際には,乳酸のナトリウム塩であるL-乳酸ナトリウムを用いた)を海馬に投与されたラットはグリコーゲン分解酵素阻害剤のみを投与されたラットに比べ高い長期記憶の能力を示した.つまり,乳酸の投与はグリコーゲン分解酵素阻害剤による長期記憶の形成能力の障害を回復させたのである.
また,電気生理学的な手法を用いた解析から,グリコーゲン分解酵素阻害剤の海馬への投与は,シナプス伝達の指標であり前シナプスの機能を示していると考えられているpaired-pulse facilitation(PPF),後シナプスの機能を示していると考えられているbasal transmission,さらに,長期可塑性の指標とされている長期増強の誘導には影響はなかった.しかしながら,グリコーゲン分解酵素阻害剤の投与は長期増強の保持を抑制し,この抑制は乳酸の投与により回復することが発見された.さらに,ウェスタンブロット法によりシナプスの構造変化に関連するタンパク質であるARCの発現量やコフィリンのリン酸化量,さらに,長期記憶の形成に必須の転写因子であるCREBのリン酸化量を調べた.その結果,トレーニングによりこれらタンパク質の発現量やリン酸化量は増加し,その増加はグリコーゲン分解酵素阻害剤の投与により阻害された.さらに,グリコーゲン分解酵素阻害剤による阻害は乳酸の投与により回復することも発見された.以上の行動学的,電気生理学的,生化学的な解析から,アストロサイトからのグリコーゲンに由来する乳酸の放出は長期記憶の形成に重要であることが示唆された.
これまで,海馬への乳酸の投与によりグリコーゲン分解酵素阻害剤によりひき起こされる記憶障害が回復することを述べたが,その詳細な分子機構はわかっていない.1994年,アストロサイト-ニューロン乳酸シャトル仮説が提唱されている11).この仮説によると,アストロサイトやニューロンには乳酸を輸送するための特異的なトランスポーター(モノカルボン酸トランスポーター,monocarboxylate transporters:MCT)が発現していて,このトランスポーターを介したアストロサイトからニューロンへのエネルギー輸送機構の存在が示されている.このモノカルボン酸トランスポーターにはこれまで14のアイソフォームが同定され,このうち脳にはMCT1,MCT2,MCT4の3つのアイソフォームが発現している12).これまでの研究から,MCT1はアストロサイトのほか毛細血管の内皮細胞およびオリゴデンドロサイトに,MCT4はおもにアストロサイトに発現しており12),一方,MCT2はニューロンに発現していることが示されている.さらに,MCT1とMCT4はアストロサイトからの乳酸の放出を制御し,MCT2はニューロンへの乳酸の取り込みを制御していることが報告されている12).しかしながら,このアストロサイトからニューロンへの乳酸の輸送機構を示した仮説の記憶形成に対する必要性はわかっていない.そこで,モノカルボン酸トランスポーターの発現を特異的に抑制するアンチセンス鎖を用いて,アストロサイト-ニューロン乳酸シャトルの記憶形成に対する必要性を検討した.
まず,アストロサイトに発現するMCT1およびMCT4に対する特異的なアンチセンス鎖を海馬へ投与し,MCT1およびMCT4の発現抑制による記憶形成への影響を調べた.アンチセンス鎖を海馬へ投与した結果,アンチセンス鎖と同じ塩基組成であるが配列の異なるスクランブル鎖を投与した対照群に比べ,有意に24時間後および1週間後の長期記憶の形成能力の障害が観察された.さらに,乳酸の投与はMCT1およびMCT4の発現抑制による長期記憶の形成能力の障害を回復させたことから,アストロサイトからの乳酸の放出は長期記憶の形成に必要であることが示唆された.
さらに,ニューロンに発現するMCT2に対する特異的なアンチセンス鎖を用いてMCT2の発現を抑制した結果でも同様に長期記憶の形成能力の障害が観察された.しかしながら,この長期記憶の障害は乳酸を投与しても回復しなかった.このことから,MCT2を介した乳酸のニューロンへの取り込みも長期記憶の形成に必要であることが示された.以上の結果から,アストロサイトからニューロンへの乳酸の輸送は長期記憶の形成に必須であることが示唆された.
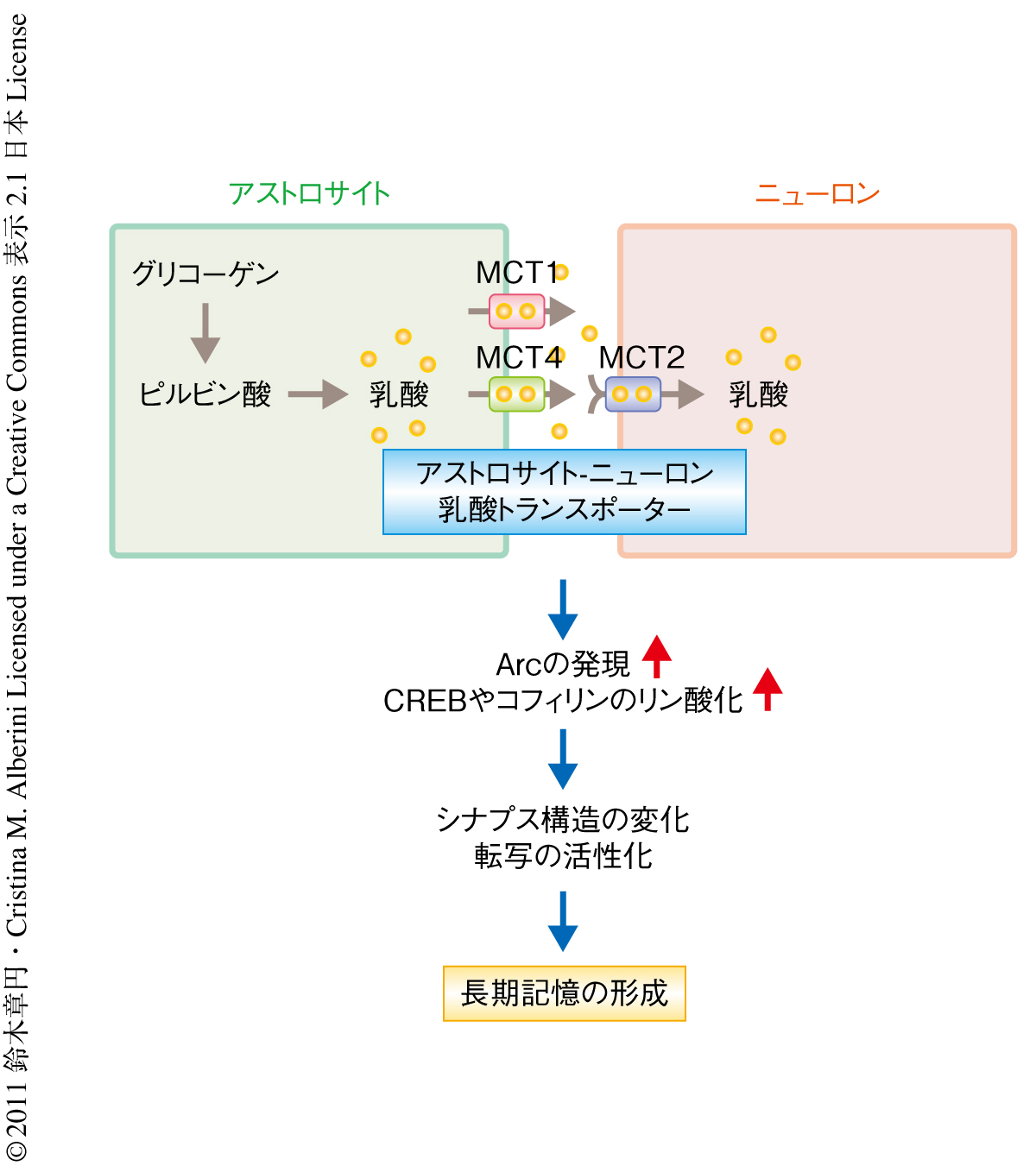
この研究によって,アストロサイトにおけるグリコーゲン分解系やアストロサイトからニューロンへの乳酸の供給がARCの発現,CREBやコフィリンのリン酸化を介した長期記憶の形成に必須であることが示された(図2).乳酸の供給は短期記憶の形成には必要ないが,長期記憶の形成に必須であることが観察された.短期記憶の形成とは異なり,長期記憶の形成には新規の遺伝子発現をともなった新規のタンパク質合成が必要となる.また,タンパク質の合成には多くのエネルギーが必要とされることから,長期記憶の形成のようなより多くのエネルギーが必要となるときにアストロサイトからのエネルギーの供給が行われるものと考えられた.しかし,アストロサイトからニューロンへ供給された乳酸はニューロンでエネルギー源としてだけ使われるのか,または,乳酸自体がある分子機構を制御することで記憶形成に関与しているのかは不明である.また,ARCやCREB,コフィリンはアストロサイトにも発現していることが報告されており,これらのタンパク質のアストロサイトにおける役割も明らかになっていない.今後の研究課題である.
略歴:2005年 東京農業大学大学院農学研究科博士後期課程 修了,同年 東京農業大学応用生物科学部 ポスドクを経て,2008年より米国Mount Sinai School of Medicineポスドク.
研究テーマ:アストロサイトとニューロンとのあいだの相互作用による記憶の制御.
© 2011 鈴木 章円 Licensed under CC 表示 2.1 日本
(米国Mount Sinai School of Medicine,Department of Neuroscience)
email:鈴木章円
DOI: 10.7875/first.author.2011.056
Astrocyte-neuron lactate transport is required for long-term memory formation.
Akinobu Suzuki, Sarah A. Stern, Ozlem Bozdagi, George W. Huntley, Ruth H. Walker, Pierre J. Magistretti, Cristina M. Alberini
Cell, 144, 810-823 (2011)
この論文に出現する遺伝子・タンパク質のUniprot ID
MCT1(P53987), MCT4(O35910), MCT2(Q63344), グルコーストランスポーター, ARC(Q63053), コフィリン, CREB, モノカルボン酸トランスポーター, monocarboxylate transporters, MCT, GLUT
要 約
脳神経系はニューロンとグリアから構成されているが,グリアの記憶形成に対する役割はほとんど明らかにされていない.この論文において筆者らは,記憶形成のときにグリアのひとつであるアストロサイトにおいてグリコーゲンから産生された乳酸のニューロンへの供給が長期記憶の形成に必須であることを明らかにした.グリコーゲン分解酵素阻害剤の海馬への直接の投与は長期記憶の形成,長期増強,さらに,記憶形成に関与するタンパク質の発現やリン酸化を阻害した.また,これらの阻害は乳酸の投与により回復することも発見された.さらに,乳酸のアストロサイトからの放出に関与するトランスポーターであるMCT1およびMCT4の発現を抑制した場合にも長期記憶の形成阻害が観察され,この阻害は乳酸の投与により回復した.一方,ニューロンへの乳酸の取り込みに関与するトランスポーターMCT2の発現を抑制した場合にも同様に記憶形成の阻害が観察されたが,乳酸の投与による回復は観察されなかったことから,アストロサイトからニューロンへの乳酸の輸送が記憶形成に必須であることが示された.
はじめに
これまで,グリアのひとつであるアストロサイトはニューロンをサポートする役割をもつものと考えられていた1).しかしながら近年,アストロサイトが神経突起の伸長やニューロンのあいだのシグナル伝達に関与すること2),さらに,記憶の細胞モデルである長期増強(long-term potentiation:LTP)の制御にも関与することが報告され3),アストロサイトがより積極的にニューロンの活動を制御する可能性が示唆されている.さらに,記憶形成ののちの海馬においてアストロサイトの数が増加することや4),アストロサイトの機能を抑制すると記憶形成が阻害されるなど5),記憶の形成に対するアストロサイトの役割に関する研究も行われはじめている.したがって,記憶など脳の高次機能はニューロンとアストロサイトとのあいだの相互作用を介して制御されることが示唆されている.ニューロンとアストロサイトとの相互作用のひとつの分子機構としてニューロンへのエネルギーの供給があげられる.グルコースは脳活動を維持するためのエネルギー源のひとつであり,グルコーストランスポーターを介してニューロンやアストロサイトに輸送されグルタミン酸などの神経伝達物質の生合成に使われる6).一方で,ニューロンとは異なり,アストロサイトにはグリコーゲンが貯蔵され7),ニューロンのエネルギー源として利用されることが明らかになっている(図1)が8),記憶形成への役割は明らかになっていなかった.この研究において,筆者らは,アストロサイトにおけるグリコーゲン分解系やアストロサイトからニューロンへの乳酸の供給が長期記憶の形成に必須であることを見い出した.
1.アストロサイトにおけるグリコーゲン代謝系の記憶形成への必要性
まず,アストロサイトにおけるグリコーゲン分解系が記憶の形成に影響をあたえるかどうかを評価するため,グリコーゲン分解酵素の阻害剤であるDAB 9)(1,4-ジジオキシ-1,4-イミノ-d-アラビニトール)をラットの脳,記憶形成に重要な役割を担っている海馬に直接に投与し,受動的回避学習課題により短期および長期の記憶形成の能力を調べた.受動的回避学習課題とはラットの暗い場所を好む習性を利用して記憶能力を評価する行動学的な解析のひとつである.この解析において,明暗の2箱に分けた実験箱の明箱側にラットを入れると,ラットは暗いところを好むためすみやかに暗箱に入る.その直後にラットに電気ショックをあたえることで暗箱という“場所”と電気ショックによる“痛み”を関連づけて学習させる(トレーニング).一定時間の経過ののち同一の実験箱の明箱側にふたたびラットを入れると,以前の電気ショックによる体験と強い嫌悪感を想起し暗箱には入らなくなる.そこで,ラットが明箱から暗箱に移動するまでの時間を記録し(テスト),その反応時間の長短から記憶能力を評価する.暗箱に入るまでの時間が長いほど強く記憶していることになる.
トレーニングの15分前に海馬にグリコーゲン分解酵素阻害剤を投与しトレーニングの1時間後に記憶能力を評価したところ,グリコーゲン分解酵素阻害剤の投与群と溶媒を投与した対照群とのあいだに有意な差は観察されず,グリコーゲン分解酵素阻害剤の投与は短期記憶の形成には影響をあたえなかった.しかしながら,24時間後に記憶能力を評価した結果では投与群は対照群に比べ暗箱に入るまでの時間が有意に短く,長期記憶の形成能力の障害が観察されその阻害はトレーニングから1週間後でも観察された.また,トレーニングの直後にグリコーゲン分解酵素阻害剤を投与した場合も同様に記憶の形成阻害が観察されたが,トレーニングの24時間後に投与しさらに24時間後に記憶能力を評価した場合には記憶阻害は観察されなかった.以上の結果から,アストロサイトにおけるグリコーゲンの分解系は短期記憶の形成には必要とされないが,より多くのエネルギーを必要とするタンパク質合成をともなう長期記憶の形成には必須であること,さらに,トレーニング前後の比較的短い期間においてグリコーゲンの分解が必要であることが示唆された.
2.グリコーゲン分解系に由来する乳酸は長期記憶の形成に必要である
つぎに,グリコーゲン分解系によって産生されたどの基質が長期記憶の形成に関与しているのかを調べた.過去の培養細胞を用いた研究から,細胞外からの刺激によりグリコーゲンの分解が促されると乳酸が合成され,この乳酸がアストロサイトから細胞外へと放出されることが報告されていた10).そこで,トレーニングののちにおいてもグリコーゲンから乳酸が合成されこれが記憶形成に関与するのかどうかを調べるため,トレーニング前後のラットの海馬における細胞外の乳酸の濃度をin vivoマイクロダイアリシス法により測定した.その結果,トレーニングののちすみやかに乳酸の濃度は上昇し,さらに,トレーニングによる乳酸の濃度上昇はグリコーゲン分解酵素阻害剤の海馬への投与により抑制された.このことから,観察された乳酸の濃度上昇はグリコーゲンの分解によりもたらされていることが示唆された.そこで,乳酸の海馬への投与によりグリコーゲン分解酵素阻害剤による記憶障害は回復するかどうかを調べるため,グリコーゲン分解酵素阻害剤と乳酸とを海馬へ同時に投与し記憶能力を評価した.受動的回避学習課題により記憶形成への影響を観察したところ,グリコーゲン分解酵素阻害剤と乳酸(実際には,乳酸のナトリウム塩であるL-乳酸ナトリウムを用いた)を海馬に投与されたラットはグリコーゲン分解酵素阻害剤のみを投与されたラットに比べ高い長期記憶の能力を示した.つまり,乳酸の投与はグリコーゲン分解酵素阻害剤による長期記憶の形成能力の障害を回復させたのである.
また,電気生理学的な手法を用いた解析から,グリコーゲン分解酵素阻害剤の海馬への投与は,シナプス伝達の指標であり前シナプスの機能を示していると考えられているpaired-pulse facilitation(PPF),後シナプスの機能を示していると考えられているbasal transmission,さらに,長期可塑性の指標とされている長期増強の誘導には影響はなかった.しかしながら,グリコーゲン分解酵素阻害剤の投与は長期増強の保持を抑制し,この抑制は乳酸の投与により回復することが発見された.さらに,ウェスタンブロット法によりシナプスの構造変化に関連するタンパク質であるARCの発現量やコフィリンのリン酸化量,さらに,長期記憶の形成に必須の転写因子であるCREBのリン酸化量を調べた.その結果,トレーニングによりこれらタンパク質の発現量やリン酸化量は増加し,その増加はグリコーゲン分解酵素阻害剤の投与により阻害された.さらに,グリコーゲン分解酵素阻害剤による阻害は乳酸の投与により回復することも発見された.以上の行動学的,電気生理学的,生化学的な解析から,アストロサイトからのグリコーゲンに由来する乳酸の放出は長期記憶の形成に重要であることが示唆された.
3.アストロサイトからニューロンへの乳酸の輸送は長期記憶の形成に必須である
これまで,海馬への乳酸の投与によりグリコーゲン分解酵素阻害剤によりひき起こされる記憶障害が回復することを述べたが,その詳細な分子機構はわかっていない.1994年,アストロサイト-ニューロン乳酸シャトル仮説が提唱されている11).この仮説によると,アストロサイトやニューロンには乳酸を輸送するための特異的なトランスポーター(モノカルボン酸トランスポーター,monocarboxylate transporters:MCT)が発現していて,このトランスポーターを介したアストロサイトからニューロンへのエネルギー輸送機構の存在が示されている.このモノカルボン酸トランスポーターにはこれまで14のアイソフォームが同定され,このうち脳にはMCT1,MCT2,MCT4の3つのアイソフォームが発現している12).これまでの研究から,MCT1はアストロサイトのほか毛細血管の内皮細胞およびオリゴデンドロサイトに,MCT4はおもにアストロサイトに発現しており12),一方,MCT2はニューロンに発現していることが示されている.さらに,MCT1とMCT4はアストロサイトからの乳酸の放出を制御し,MCT2はニューロンへの乳酸の取り込みを制御していることが報告されている12).しかしながら,このアストロサイトからニューロンへの乳酸の輸送機構を示した仮説の記憶形成に対する必要性はわかっていない.そこで,モノカルボン酸トランスポーターの発現を特異的に抑制するアンチセンス鎖を用いて,アストロサイト-ニューロン乳酸シャトルの記憶形成に対する必要性を検討した.
まず,アストロサイトに発現するMCT1およびMCT4に対する特異的なアンチセンス鎖を海馬へ投与し,MCT1およびMCT4の発現抑制による記憶形成への影響を調べた.アンチセンス鎖を海馬へ投与した結果,アンチセンス鎖と同じ塩基組成であるが配列の異なるスクランブル鎖を投与した対照群に比べ,有意に24時間後および1週間後の長期記憶の形成能力の障害が観察された.さらに,乳酸の投与はMCT1およびMCT4の発現抑制による長期記憶の形成能力の障害を回復させたことから,アストロサイトからの乳酸の放出は長期記憶の形成に必要であることが示唆された.
さらに,ニューロンに発現するMCT2に対する特異的なアンチセンス鎖を用いてMCT2の発現を抑制した結果でも同様に長期記憶の形成能力の障害が観察された.しかしながら,この長期記憶の障害は乳酸を投与しても回復しなかった.このことから,MCT2を介した乳酸のニューロンへの取り込みも長期記憶の形成に必要であることが示された.以上の結果から,アストロサイトからニューロンへの乳酸の輸送は長期記憶の形成に必須であることが示唆された.
おわりに
この研究によって,アストロサイトにおけるグリコーゲン分解系やアストロサイトからニューロンへの乳酸の供給がARCの発現,CREBやコフィリンのリン酸化を介した長期記憶の形成に必須であることが示された(図2).乳酸の供給は短期記憶の形成には必要ないが,長期記憶の形成に必須であることが観察された.短期記憶の形成とは異なり,長期記憶の形成には新規の遺伝子発現をともなった新規のタンパク質合成が必要となる.また,タンパク質の合成には多くのエネルギーが必要とされることから,長期記憶の形成のようなより多くのエネルギーが必要となるときにアストロサイトからのエネルギーの供給が行われるものと考えられた.しかし,アストロサイトからニューロンへ供給された乳酸はニューロンでエネルギー源としてだけ使われるのか,または,乳酸自体がある分子機構を制御することで記憶形成に関与しているのかは不明である.また,ARCやCREB,コフィリンはアストロサイトにも発現していることが報告されており,これらのタンパク質のアストロサイトにおける役割も明らかになっていない.今後の研究課題である.
文 献
- Kettenmann, H. & Ransom, B. R. (eds.): Neuroglia, 2nd ed. Oxford University Press, New York (2005)
- Haydon, P. G. & Carmignoto, G.: Astrocyte control of synaptic transmission and neurovascular coupling. Physiol. Rev., 86, 1009-1031 (2006)[PubMed]
- Henneberger, C., Papouin, T., Oliet, S. H. et al.: Long-term potentiation depends on release of D-serine from astrocytes. Nature, 463, 232-236 (2010)[PubMed]
- Jahanshahi, M., Sadeghi, Y., Hosseini, A. et al.: The effect of spatial learning on the number of astrocytes in the CA3 subfield of the rat hippocampus. Singapore Med. J., 49, 388-391 (2008)[PubMed]
- Gibbs, M. E., O'Dowd, B. S., Hertz, E. et al.: Astrocytic energy metabolism consolidates memory in young chicks. Neuroscience, 141, 9-13 (2006)[PubMed]
- Hamprecht, B., Verleysdonk, S. & Wiesinger, H.: Enzymes of carbohydrate and energy metabolism. in Neuroglia, 2nd ed. (Kettenmann, H. & Ransom, B. R. eds.), pp. 202-215, Oxford University Press, New York (2005)
- Brown, A. M., Baltan Tekkok, S. & Ransom, B. R.: Energy transfer from astrocytes to axons: the role of CNS glycogen. Neurochem. Int., 45, 529-536 (2004)[PubMed]
- Swanson, R. A.: Physiologic coupling of glial glycogen metabolism to neuronal activity in brain. Can. J. Physiol. Pharmacol., 70, 138-144 (1992)[PubMed]
- Walls, A. B., Sickmann, H. M., Brown, A. et al.: Characterization of 1,4-dideoxy-1,4-imino-d-arabinitol (DAB) as an inhibitor of brain glycogen shunt activity. J. Neurochem., 105, 1462-1470 (2008)[PubMed]
- Dringen, R., Gebhardt, R. & Hamprecht, B.: Glycogen in astrocytes: possible function as lactate supply for neighboring cells. Brain Res., 623, 208-214 (1993)[PubMed]
- Pellerin, L. & Magistretti, P. J.: Glutamate uptake into astrocytes stimulates aerobic glycolysis: a mechanism coupling neuronal activity to glucose utilization. Proc. Natl. Acad. Sci. USA, 91, 10625-10629 (1994)[PubMed]
- Pierre, K. & Pellerin, L.: Monocarboxylate transporters in the central nervous system: distribution, regulation and function. J. Neurochem., 94, 1-14 (2005)[PubMed]
著者プロフィール
略歴:2005年 東京農業大学大学院農学研究科博士後期課程 修了,同年 東京農業大学応用生物科学部 ポスドクを経て,2008年より米国Mount Sinai School of Medicineポスドク.
研究テーマ:アストロサイトとニューロンとのあいだの相互作用による記憶の制御.
© 2011 鈴木 章円 Licensed under CC 表示 2.1 日本
