神経系と中胚葉は共通の前駆体である体軸幹細胞から発生し転写因子Sox2とTbx6がその発生運命を決める
竹本龍也・近藤寿人
(大阪大学大学院生命機能研究科 形態形成研究室)
email:近藤寿人
DOI: 10.7875/first.author.2011.043
Tbx6-dependent Sox2 regulation determines neural or mesodermal fate in axial stem cells.
Tatsuya Takemoto, Masanori Uchikawa, Megumi Yoshida, Donald M. Bell, Robin Lovell-Badge, Virginia E. Papaioannou, Hisato Kondoh
Nature, 470, 394-398 (2011)
多くの教科書の記述では“胚の組織はまず外胚葉,中胚葉,内胚葉のいずれかに分かれ,それが胚の細胞の発生運命をせばめてゆく第一歩である”とされている.この通説では,神経系はひとたび“外胚葉”が成立してから表皮と分離して生じるものであり,中胚葉に属する体節(骨や筋肉のもと)などとはおよそ縁もゆかりもないということになる.しかし,筆者らの研究によって,それとはあいいれない新しい細胞系譜の分岐とその調節が示された.体の胴部では神経系と中胚葉とが共通の前駆体である“体軸幹細胞”から生まれること,そして,そのいずれになるのかは神経系の発生を指令するSox2と中胚葉への発生を指令するTbx6という2つの転写因子の力関係で決まることが示された.この研究成果は,三胚葉の意義についての旧来の考え方を根本からあらためるものであった.
筆者らのグループでは,神経系を生み出す重要な調節タンパク質であるSox2とそれをコードするSox2遺伝子について十年来にわたり研究してきた.Sox2は着床前の胚やその発生時期に対応するES細胞(embryonic stem cell,胚性幹細胞)およびiPS細胞(induced pluripotent stem cell,人工多能性幹細胞)において重要な調節機能を担っており,そのためSox2遺伝子はiPS細胞を作製するのに必要な4つの遺伝子のうちのひとつとなっている.しかし,着床後の2層状態の胚において,Sox2は胚盤葉(上層)の成立に中心的な機能をはたしたのち,2層の胚構造が3層に再編成される“原腸陥入”の時期からは神経系の原基である“神経板”をつくるための主要なタンパク質として機能し,実際にSox2遺伝子の発現は神経板の形成と対応することからSox2遺伝子の発現は“pan-neural marker”(神経系に普遍的なマーカー)とみなされることもある(図1).
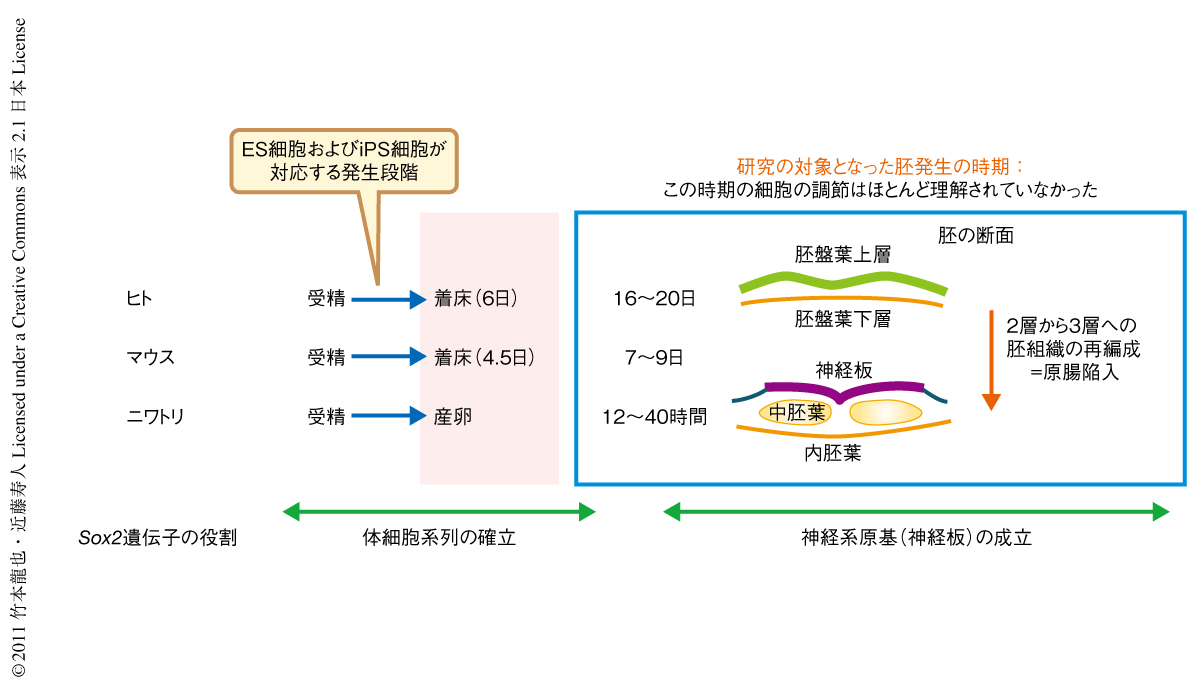
筆者らは,Sox2遺伝子の神経板における発現は神経板の発生段階と領域ごとに異なるエンハンサー(遺伝子の近傍のゲノム領域にある制御機能をもつDNA配列)により制御されていることを示し1,2),とくに,原腸陥入の開始直後の段階では頭部の神経板の形成にかかわるエンハンサーN2と胴部の神経板の形成にかかわるエンハンサーN1のみが活性をもつこと(図2),そして,エンハンサーN1とエンハンサーN2とがまったく異なった制御をうけることから3,4),頭部の神経板(将来の脳)と胴部の神経板(将来の脊髄など)とがまったく異なったしくみで生まれることを示した.
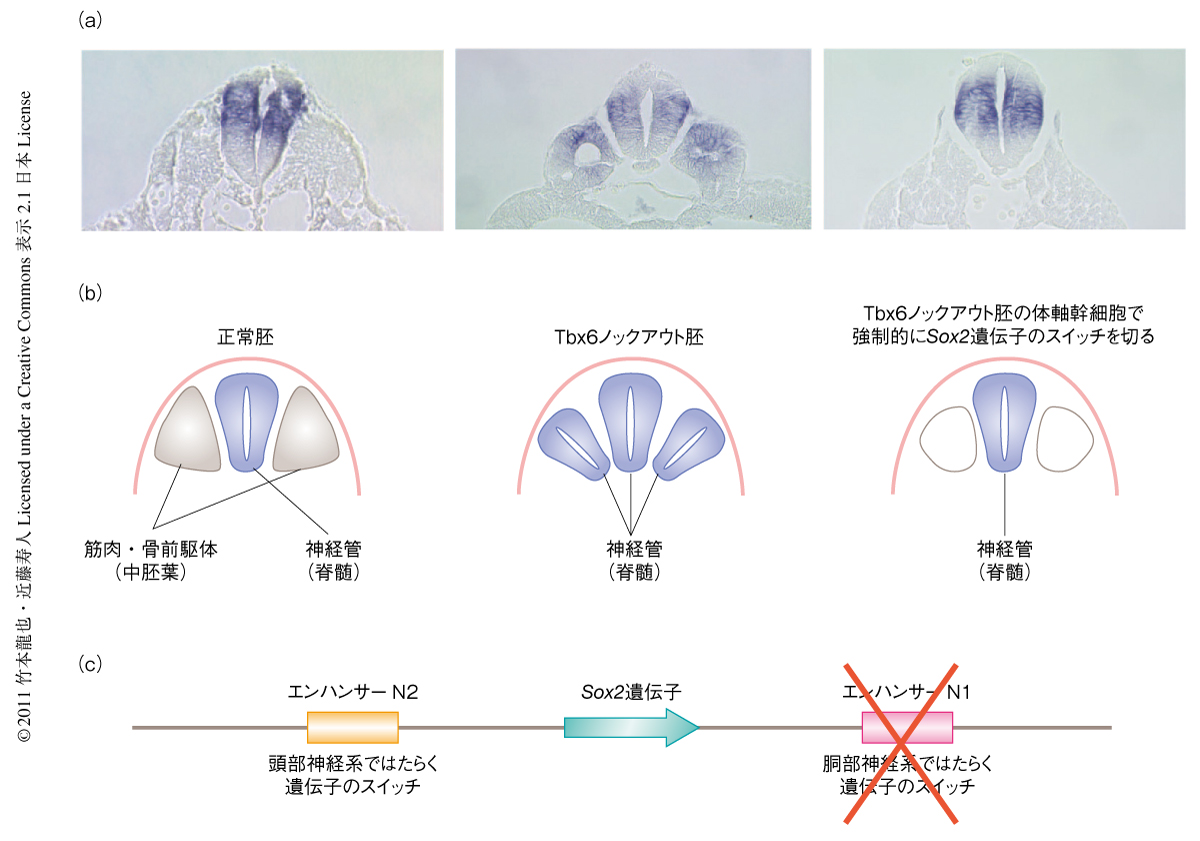
胴部神経系については,まず,中胚葉と神経系が体軸幹細胞というべき同じ前駆体に由来することが示唆された3).胴部神経板においてエンハンサーN1が活性をもつ細胞をエンハンサーN1のレポーター遺伝子などにより調べると,それは胚を2層から3層に変える細胞移動を行うまえの上層に位置する細胞であった.上層にとどまる細胞はエンハンサーN1の活性がオンのままSox2遺伝子を活性化して神経板になる.しかし,上層からすぐ下の中間層(骨や筋肉のもとになる中胚葉の層)に移動した細胞ではひとたび活性化されたエンハンサーN1が再度その活性を失う.このことはつまり,中胚葉をつくるにはSox2遺伝子のエンハンサーN1の活性を遮断する必要があるのではないかということを示唆し,さらには,もしSox2が中間層ではたらいてしまえば中胚葉はできず神経系になってしまうのではないかという可能性を示唆していた.
筆者らは,この2006年の研究をさらに発展させ,そして,つぎに述べる“Tbx6遺伝子のノックアウトマウスでみられる不思議な現象を解明する”という方法で,中胚葉と神経系とがその共通の前駆体である“体軸幹細胞”から生まれることを証明した.
転写因子をコードするTbx6遺伝子のノックアウトマウス胚では骨や筋肉のもとになる中胚葉の代わりに脊髄が発生してしまい,その結果,3本の脊髄をもつという不思議な現象が1998年に報告されていた5).この現象は,三胚葉に関する旧来のモデルでは説明できない.今回の研究で,Tbx6ノックアウト胚では神経系と中胚葉の共通の前駆体である“体軸幹細胞”において,中間層に移動したあともエンハンサーN1の活性が遮断されることなく持続してしまった結果,Sox2遺伝子が発現され異所的な脊髄をつくることが示された.つまり,Tbx6ノックアウトマウス胚にみられた不思議な現象の謎を解き明かすとともに,胴部では神経板と中胚葉組織とが共通の前駆体から直接に分かれて生じることを示した(図2).
今回の研究では,中間の細胞層に移動した細胞集団においてSox2遺伝子の転写の活性化を抑えるためエンハンサーN1のノックアウトマウスを用いることにより明快な結論を得ることができた.これは筆者らの十年来の研究のひとつの結実であった.
さらに,つぎの重要な結論を得ることもできた.1)Tbx6もSox2もはたらかなかった中間層の細胞は,神経系の性質も中胚葉の性質ももたなかった.したがって,Tbx6は中間層においてSox2遺伝子のエンハンサーN1の活性を遮断する作用をもつだけでなく,中胚葉自体を生み出すのにも必要である.2)野生型胚の中間層でSox2を強制発現することによりTbx6がはたらいている状況でもSox2の作用により中間層において神経系を生み出すことができた.したがって,Tbx6のもつエンハンサーN1の活性を抑制する作用は中胚葉を生み出すこと自体に必須である.3)Tbx6がエンハンサーN1を抑制するのは,Tbx6がその配列に直接に結合するためではなく間接的な作用によるものであった.エンハンサーN1はWntシグナルとFgfシグナルの双方が協同的にはたらくことにより活性化された.Tbx6ノックアウトマウス胚では中間層において発現が抑制されるはずのWnt3aが上層と同様に強く発現されるため,中間層でエンハンサーN1が活性をもつことになってしまった.
これらの結果から,神経系と中胚葉との共通の前駆体である“体軸幹細胞”は,Sox2とTbx6という2つの転写因子の活性のせめぎあいの結果により神経系にも中胚葉にもなりうるのであり,中間層に細胞が移動すること自体は細胞の将来を決める機構ではないことが示された.
最近になり,世界的には少数派ではあるが,“神経系は三胚葉の分離のあとに外胚葉から成立する”という旧来の説に異議をとなえる新しい細胞系譜の解析データを報告するグループがでてきた.2009年に発表された細胞系譜に関する論文では,神経板と中胚葉の双方に子孫細胞を残す細胞が存在すること,したがって,“体軸幹細胞”の存在を考えなければ細胞系譜のデータを説明できないことが示されていた6).今回の研究は,その新しい細胞系譜のデータを裏づけるとともに,具体的な調節機構を明らかにすることにより,発生生物学の教科書的な定説をつくがえす,神経系の成立についての新しい調節機構と細胞系譜を示したものであった(図3).

では,これまでの三胚葉モデル(教科書の定説)はどこに由来しているのだろうか? 現在の生物学の教科書が大きなよりどころとしているのは1920年代~1930年代の両生類胚の研究である.しかし,当時の発生生物学のパイオニアたちは大胆にイマジネーションを駆使しながらも結論には慎重であった.当時は作業仮説であった考え方がいつしか後継者たちにより教条化されたものと推定される.さまざまな新しい研究方法を駆使できる現代において90年前の発想が教条化されていた経緯には,科学のあり方として大いに反省すべき点がある.
実際に,1930年代から,今回の研究の結論を支持する(当時では説明が困難な)さまざまな実験結果が得られていた.そして,それに対する説明をあたえることが将来への課題であると当時の研究者により指摘されていた.一例は1938年に報告された実験である7,8).中胚葉になるはずの(予定中胚葉領域の)組織片を培養すると,神経系と中胚葉の双方の組織があたかも胚の胴部を展開したかのような配置をもって発生してきた.これは実は,神経板と中胚葉の共通の前駆体,すなわち,“体軸幹細胞”を培養していたのである.今回の研究は,古典期の発生生物学が説明できなかった課題を正しい発生過程の調節機構を示すことによって解決したものといえる.
現代の幹細胞科学の将来には大きな期待がよせられているが,ES細胞やiPS細胞などから特定の体の組織をつくるためにも,また,体の発生の中間段階の幹細胞を研究する場合にも,胚の段階でどのような前駆体からどのような細胞種が生まれるかを正しく知っておく必要がある.しかし,これまでの幹細胞研究は誤りともいうべき旧来の三胚葉説を前提として進められていることが多かった.筆者らの研究は,細胞系譜の分離とその調節についての新しくそして正しい情報を提供し,幹細胞の研究に新しい指針を示した.
略歴:2005年 大阪大学大学院理学研究科博士課程 修了,同年より大阪大学大学院生命機能研究科 特任助手(現 助教).
研究テーマ:神経系原基細胞と中胚葉系細胞を生みだす体軸幹細胞の制御.
抱負:胚発生の研究をつうじて,個体がうまくつくられるしくみを楽しみながら明らかにしたい.
近藤 寿人(Hisato Kondoh)
大阪大学大学院生命機能研究科 教授.
研究室URL:http://www.fbs.osaka-u.ac.jp/jpn/general/lab/06/
© 2011 竹本龍也・近藤寿人 Licensed under CC 表示 2.1 日本
(大阪大学大学院生命機能研究科 形態形成研究室)
email:近藤寿人
DOI: 10.7875/first.author.2011.043
Tbx6-dependent Sox2 regulation determines neural or mesodermal fate in axial stem cells.
Tatsuya Takemoto, Masanori Uchikawa, Megumi Yoshida, Donald M. Bell, Robin Lovell-Badge, Virginia E. Papaioannou, Hisato Kondoh
Nature, 470, 394-398 (2011)
要 約
多くの教科書の記述では“胚の組織はまず外胚葉,中胚葉,内胚葉のいずれかに分かれ,それが胚の細胞の発生運命をせばめてゆく第一歩である”とされている.この通説では,神経系はひとたび“外胚葉”が成立してから表皮と分離して生じるものであり,中胚葉に属する体節(骨や筋肉のもと)などとはおよそ縁もゆかりもないということになる.しかし,筆者らの研究によって,それとはあいいれない新しい細胞系譜の分岐とその調節が示された.体の胴部では神経系と中胚葉とが共通の前駆体である“体軸幹細胞”から生まれること,そして,そのいずれになるのかは神経系の発生を指令するSox2と中胚葉への発生を指令するTbx6という2つの転写因子の力関係で決まることが示された.この研究成果は,三胚葉の意義についての旧来の考え方を根本からあらためるものであった.
はじめに
筆者らのグループでは,神経系を生み出す重要な調節タンパク質であるSox2とそれをコードするSox2遺伝子について十年来にわたり研究してきた.Sox2は着床前の胚やその発生時期に対応するES細胞(embryonic stem cell,胚性幹細胞)およびiPS細胞(induced pluripotent stem cell,人工多能性幹細胞)において重要な調節機能を担っており,そのためSox2遺伝子はiPS細胞を作製するのに必要な4つの遺伝子のうちのひとつとなっている.しかし,着床後の2層状態の胚において,Sox2は胚盤葉(上層)の成立に中心的な機能をはたしたのち,2層の胚構造が3層に再編成される“原腸陥入”の時期からは神経系の原基である“神経板”をつくるための主要なタンパク質として機能し,実際にSox2遺伝子の発現は神経板の形成と対応することからSox2遺伝子の発現は“pan-neural marker”(神経系に普遍的なマーカー)とみなされることもある(図1).
筆者らは,Sox2遺伝子の神経板における発現は神経板の発生段階と領域ごとに異なるエンハンサー(遺伝子の近傍のゲノム領域にある制御機能をもつDNA配列)により制御されていることを示し1,2),とくに,原腸陥入の開始直後の段階では頭部の神経板の形成にかかわるエンハンサーN2と胴部の神経板の形成にかかわるエンハンサーN1のみが活性をもつこと(図2),そして,エンハンサーN1とエンハンサーN2とがまったく異なった制御をうけることから3,4),頭部の神経板(将来の脳)と胴部の神経板(将来の脊髄など)とがまったく異なったしくみで生まれることを示した.
胴部神経系については,まず,中胚葉と神経系が体軸幹細胞というべき同じ前駆体に由来することが示唆された3).胴部神経板においてエンハンサーN1が活性をもつ細胞をエンハンサーN1のレポーター遺伝子などにより調べると,それは胚を2層から3層に変える細胞移動を行うまえの上層に位置する細胞であった.上層にとどまる細胞はエンハンサーN1の活性がオンのままSox2遺伝子を活性化して神経板になる.しかし,上層からすぐ下の中間層(骨や筋肉のもとになる中胚葉の層)に移動した細胞ではひとたび活性化されたエンハンサーN1が再度その活性を失う.このことはつまり,中胚葉をつくるにはSox2遺伝子のエンハンサーN1の活性を遮断する必要があるのではないかということを示唆し,さらには,もしSox2が中間層ではたらいてしまえば中胚葉はできず神経系になってしまうのではないかという可能性を示唆していた.
1.Tbx6ノックアウトマウスの表現型
筆者らは,この2006年の研究をさらに発展させ,そして,つぎに述べる“Tbx6遺伝子のノックアウトマウスでみられる不思議な現象を解明する”という方法で,中胚葉と神経系とがその共通の前駆体である“体軸幹細胞”から生まれることを証明した.
転写因子をコードするTbx6遺伝子のノックアウトマウス胚では骨や筋肉のもとになる中胚葉の代わりに脊髄が発生してしまい,その結果,3本の脊髄をもつという不思議な現象が1998年に報告されていた5).この現象は,三胚葉に関する旧来のモデルでは説明できない.今回の研究で,Tbx6ノックアウト胚では神経系と中胚葉の共通の前駆体である“体軸幹細胞”において,中間層に移動したあともエンハンサーN1の活性が遮断されることなく持続してしまった結果,Sox2遺伝子が発現され異所的な脊髄をつくることが示された.つまり,Tbx6ノックアウトマウス胚にみられた不思議な現象の謎を解き明かすとともに,胴部では神経板と中胚葉組織とが共通の前駆体から直接に分かれて生じることを示した(図2).
今回の研究では,中間の細胞層に移動した細胞集団においてSox2遺伝子の転写の活性化を抑えるためエンハンサーN1のノックアウトマウスを用いることにより明快な結論を得ることができた.これは筆者らの十年来の研究のひとつの結実であった.
2.体軸幹細胞の制御
さらに,つぎの重要な結論を得ることもできた.1)Tbx6もSox2もはたらかなかった中間層の細胞は,神経系の性質も中胚葉の性質ももたなかった.したがって,Tbx6は中間層においてSox2遺伝子のエンハンサーN1の活性を遮断する作用をもつだけでなく,中胚葉自体を生み出すのにも必要である.2)野生型胚の中間層でSox2を強制発現することによりTbx6がはたらいている状況でもSox2の作用により中間層において神経系を生み出すことができた.したがって,Tbx6のもつエンハンサーN1の活性を抑制する作用は中胚葉を生み出すこと自体に必須である.3)Tbx6がエンハンサーN1を抑制するのは,Tbx6がその配列に直接に結合するためではなく間接的な作用によるものであった.エンハンサーN1はWntシグナルとFgfシグナルの双方が協同的にはたらくことにより活性化された.Tbx6ノックアウトマウス胚では中間層において発現が抑制されるはずのWnt3aが上層と同様に強く発現されるため,中間層でエンハンサーN1が活性をもつことになってしまった.
これらの結果から,神経系と中胚葉との共通の前駆体である“体軸幹細胞”は,Sox2とTbx6という2つの転写因子の活性のせめぎあいの結果により神経系にも中胚葉にもなりうるのであり,中間層に細胞が移動すること自体は細胞の将来を決める機構ではないことが示された.
3.細胞系譜との関連
最近になり,世界的には少数派ではあるが,“神経系は三胚葉の分離のあとに外胚葉から成立する”という旧来の説に異議をとなえる新しい細胞系譜の解析データを報告するグループがでてきた.2009年に発表された細胞系譜に関する論文では,神経板と中胚葉の双方に子孫細胞を残す細胞が存在すること,したがって,“体軸幹細胞”の存在を考えなければ細胞系譜のデータを説明できないことが示されていた6).今回の研究は,その新しい細胞系譜のデータを裏づけるとともに,具体的な調節機構を明らかにすることにより,発生生物学の教科書的な定説をつくがえす,神経系の成立についての新しい調節機構と細胞系譜を示したものであった(図3).
4.これまでの三胚葉モデルの由来と問題点
では,これまでの三胚葉モデル(教科書の定説)はどこに由来しているのだろうか? 現在の生物学の教科書が大きなよりどころとしているのは1920年代~1930年代の両生類胚の研究である.しかし,当時の発生生物学のパイオニアたちは大胆にイマジネーションを駆使しながらも結論には慎重であった.当時は作業仮説であった考え方がいつしか後継者たちにより教条化されたものと推定される.さまざまな新しい研究方法を駆使できる現代において90年前の発想が教条化されていた経緯には,科学のあり方として大いに反省すべき点がある.
実際に,1930年代から,今回の研究の結論を支持する(当時では説明が困難な)さまざまな実験結果が得られていた.そして,それに対する説明をあたえることが将来への課題であると当時の研究者により指摘されていた.一例は1938年に報告された実験である7,8).中胚葉になるはずの(予定中胚葉領域の)組織片を培養すると,神経系と中胚葉の双方の組織があたかも胚の胴部を展開したかのような配置をもって発生してきた.これは実は,神経板と中胚葉の共通の前駆体,すなわち,“体軸幹細胞”を培養していたのである.今回の研究は,古典期の発生生物学が説明できなかった課題を正しい発生過程の調節機構を示すことによって解決したものといえる.
おわりに
現代の幹細胞科学の将来には大きな期待がよせられているが,ES細胞やiPS細胞などから特定の体の組織をつくるためにも,また,体の発生の中間段階の幹細胞を研究する場合にも,胚の段階でどのような前駆体からどのような細胞種が生まれるかを正しく知っておく必要がある.しかし,これまでの幹細胞研究は誤りともいうべき旧来の三胚葉説を前提として進められていることが多かった.筆者らの研究は,細胞系譜の分離とその調節についての新しくそして正しい情報を提供し,幹細胞の研究に新しい指針を示した.
文 献
- Uchikawa, M., Ishida, Y., Takemoto, T. et al.: Functional analysis of chicken Sox2 enhancers highlights an array of diverse regulatory elements that are conserved in mammals. Dev. Cell, 4, 509-519 (2003)[PubMed]
- Kamachi, Y., Iwafuchi, M., Okuda, Y. et al.: Evolution of non-coding regulatory sequences involved in the developmental process: reflection of differential employment of paralogous genes as highlighted by Sox2 and group B1 Sox genes. Proc. Jpn. Acad. Ser. B, 85, 55-68 (2009)[PubMed]
- Takemoto, T., Uchikawa, M., Kamachi, Y. et al.: Convergence of Wnt and FGF signals in the genesis of posterior neural plate through activation of the Sox2 enhancer N-1. Development, 133, 297-306 (2006)[PubMed]
- Iwafuchi-Doi, M., Yoshida, Y., Onichtchouk, D. et al.: The Pou5f1/Pou3f-dependent but SoxB-independent regulation of conserved enhancer N2 initiates Sox2 expression during epiblast to neural plate stages in vertebrates. Dev Biol., 352, 354-366 (2011)[PubMed]
- Chapman, D. & Papaioannou, V.: Three neural tubes in mouse embryos with mutations in the T-box gene Tbx6. Nature, 391, 695-697 (1998)[PubMed]
- Tzouanacou, E., Wegener, A., Wymeersch, F. et al.: Redefining the progression of lineage segregations during mammalian embryogenesis by clonal analysis. Dev. Cell, 17, 365-376 (2009)[PubMed]
- Holtfreter, J.: Differenzierungspotenzen isolierter Teile der Urodelengastrula. [Differentiation potentials of isolated portions of urodele gastrula.] Roux' Arch. Entw. Mech., 138, 522-656 (1938)
- Holtfreter, J. & Hamburger, V.: Amphibians. in Analysis of Development (Miller, B. H., Weiss, P. A., Hamburger, V. eds.), pp.230-297, W. B. Saunders, Philadelphia (1955)
著者プロフィール
略歴:2005年 大阪大学大学院理学研究科博士課程 修了,同年より大阪大学大学院生命機能研究科 特任助手(現 助教).
研究テーマ:神経系原基細胞と中胚葉系細胞を生みだす体軸幹細胞の制御.
抱負:胚発生の研究をつうじて,個体がうまくつくられるしくみを楽しみながら明らかにしたい.
近藤 寿人(Hisato Kondoh)
大阪大学大学院生命機能研究科 教授.
研究室URL:http://www.fbs.osaka-u.ac.jp/jpn/general/lab/06/
© 2011 竹本龍也・近藤寿人 Licensed under CC 表示 2.1 日本
