ショウジョウバエの雄特異的な介在ニューロンは雌への接触により活性化され定型的な求愛行動の引き金をひく
古波津 創・小金澤雅之・山元大輔
(東北大学大学院生命科学研究科 脳機能遺伝分野)
email:山元大輔
DOI: 10.7875/first.author.2011.030
Female contact activates male-specific interneurons that trigger stereotypic courtship behavior in Drosophila.
Soh Kohatsu, Masayuki Koganezawa & Daisuke Yamamoto
Neuron, 69, 498-508 (2011)
ショウジョウバエのfruitless遺伝子は雄の性行動に必要な神経回路の主要部を組み立てるはたらきをもつ.高温になると開口するdTrpA1チャネルをfru発現ニューロンに限定して発現させ人工的にこのニューロンを興奮させたところ,雄は相手となる雌のいない容器内で定型的な求愛行動を実行した.少数の細胞にだけdTrpA1を発現させる実験から雄の性行動を開始させる能力のある細胞としてP1クラスターとP2bクラスターを同定した.拘束した条件で行動させ,同時に,ニューロンの活動をCa2+イメージングにより記録する実験により,雄が前脚で雌の腹部を触ってフェロモンを感知するとP1クラスターが一過性に興奮することが判明した.したがって,P1クラスターは定型的な求愛行動を開始させる中枢をなすものと考えられた.
動物の本能行動が神経系においていかにかたちづくられ,数あるレパートリーのなかから適切な行動が状況に応じどのように選択され実行されるのかという問題は,動物行動学が解決すべき主要なテーマのひとつである.Tinbergenは,さまざまな動物の行動観察から,行動のレパートリーごとに固有の中枢が存在し,それらのあいだでの協調的あるいは競合的な神経接続を介して行動の選択がなされると想定した1).行動中枢はより下位の中枢を階層的に活性化して,選択されたひとつのレパートリーを構成する行動要素のそれぞれをつくりだし,最後には定型的な運動出力を自動的に生み出すとされる.これが生得的解発機構(innate releasing mechanism:IRM)であり,これを起動させる特異的な感覚刺激が鍵刺激(releaser)である.
Wiersma & Ikedaは,ザリガニの遊泳肢の体節をこえた協調的な運動をささえる神経機構の分析を試み,腹髄のあるニューロン軸索に緊張性の発火が生じさえすればこの運動が完璧なかたちで実行されることを示し,そのニューロン軸索を司令線維とよんだ2).司令線維は行動中枢の決定を下位の運動中枢へと伝えるはたらきをしているものと考えられる.そののち,コオロギを用いた一連の研究から司令線維の発火頻度の違いにより異なる歌のレパートリーのひき起こされることが示されている3).しかし,司令線維を駆動する上位の行動中枢の実体は明確になっておらず,司令線維そのものの同定も多くの場合はあいまいなままであった.
行動の選択と制御の中枢機構を解明するためには,これらの司令的な機能をもつニューロンの同定と,その活動の人為的な操作が不可欠である.キイロショウジョウバエを用いると遺伝的な手法により個々のニューロンの標識と操作とが可能である.この昆虫の性行動はきわめて定型的で要素的な行動を明確に定義できることから,Tinbergenが想定した生得的解発機構の実体を解明する好個の素材となる.この研究の目的は,ショウジョウバエを用いて求愛の行動中枢を同定することにあった.
キイロショウジョウバエの性行動は以下の要素から成り立っている.まず,雄が雌にむかって定位(orientation),接近し,前脚で雌の腹部を叩いて(タッピング,tapping)追跡しながら,片方の翅を打ち振るわせてラブソングを奏でる(シンギング,singing).雌の動きが遅くなると雄は背後にまわり口吻で雌の交尾器をなめる(リッキング,licking).つづいて,腹部を内側に曲げて雌にマウントしようとする(交尾試行,attempted copulation).雌が性的に受け入れる“気持ち”になると膣口を開き翅を立てて交尾(copulation)を可能にする.交尾の受け入れ以外のこれらの行動はすべて雄に特異的であり,野生型の雌は決してこの一連の行動をとることはない.野生型の雄は,通常,雌に対して求愛行動をとり,求愛対象がいなければ決してこの行動はせず,また,雄に対しても持続的に求愛をすることはない.
しかし,ある遺伝子の機能が低下すると,雄が雄に求愛したり,雌にも雄にも求愛しなくなったりといった行動の変化が起こる.その遺伝子はfruitless(fru)の名で知られている4,5).fru遺伝子の1次転写産物は性によって異なるスプライシングをうけ,翻訳の起こるのは雄の神経系のみであり,雌の神経系はFruタンパク質を欠く.fru遺伝子の改変により雌雄いずれでもその翻訳の起こるfruM変異体が,相同組換えによるノックイン法を用いて作製されている.注目すべきことに,fruMホモ接合体の雌個体はほかの雌に対して雄の求愛行動を示す6).つまり,fruM遺伝子を発現するニューロンがFruタンパク質をもつ場合に雄の行動が生じることになる.このことは,Fruタンパク質が神経系の雄化因子であることを示唆している.ただし,fru遺伝子がおもにはたらくのは成虫の神経系のかたちづくられる蛹期であり,成虫となって実際に求愛する際にはもはや必要でないと考えられている.実際,fru発現ニューロンのなかには雄特異的に存在するものや性的二型を示すものがあるが7,8),そうした性差は蛹期に形成されるもので成虫羽化ののち性により変化することはないと思われる.
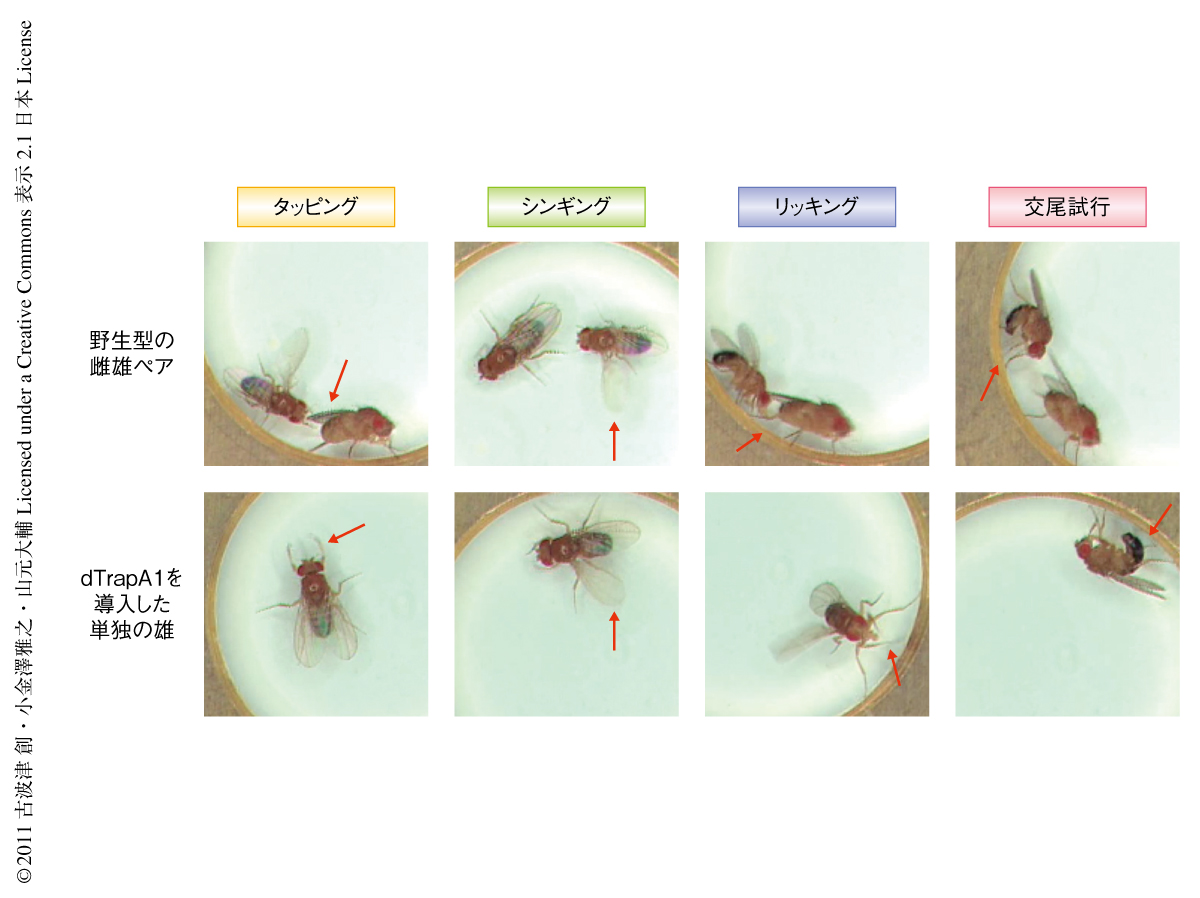
ショウジョウバエの脳には105個ほどのニューロンがあるとされるが,fru遺伝子を発現するのはそのうちの約2000個である.これまで述べた知見から,雄の求愛行動を制御するニューロンはこの2000個に含まれているものと期待された.この仮説を検証するため,雄の脳のfru発現ニューロンを強制的に活性化させることによって求愛行動を人工的にひき起こすことが可能かどうかを調べた.ニューロンの強制的な活性化を惹起するツールとして,温度感受性イオンチャネルのdTrpA1を用いた.dTrpA1は高温で開口して陽イオンの流入をひき起こしニューロンを興奮させる9).dTrpA1をfru発現ニューロンにのみ発現させる方策としてGal4-UAS系を活用した.Gal4は酵母の転写因子で,UASはその結合標的配列である.UAS-dTrpA1遺伝子を導入した個体とfru発現細胞でGal4を発現するfruNP21系統個体とを掛け合わせて得た個体では,Gal4タンパク質をもつ細胞,すなわち,fru発現ニューロンにおいてのみUAS-dTrpA1遺伝子が転写される.その結果,fru発現ニューロンはdTrpA1をもつことになり,高温条件ではその開口により興奮を起こす.このような雄を単独でチェンバーに入れ観察すると,20℃では普通の雄と同じく容器内を歩き回るばかりであるが,26~30℃に温度を上げると,驚くべきことに相手がいないにもかかわらず,一連の求愛行動をくり返し示すことがわかった(図1).
fruNP21系統ではfru発現細胞の約80%にGal4が発現する.このうちどの細胞が求愛行動の惹起に寄与するのかを明らかにするため,MARCM(mosaic analysis with a repressible cell marker)法10) によってdTrpA1をより少数のfru発現細胞(たとえば,数十個)に限定して発現させ強制活性化を行うことにした.MARCM法は発生の過程で一部の細胞に体細胞染色体組換えを誘導するもので,これを利用すると,組換えが起こった細胞やその子孫細胞にのみdTrpA1を発現させることができる.dTrpA1をもつ細胞は同時に可視マーカー(mCD8::GFP)を発現するよう仕組んでおき,行動実験のあと各個体から脳を摘出しマーカーを発現する細胞を抗GFP抗体により染色し同定する.高温で求愛行動を示す個体では求愛行動をひき起こす能力のあるニューロンにdTrpA1が発現しているはずであり,そのニューロンは可視マーカーで標識される.一方,高温にしても求愛行動を示さない個体においてあるニューロンがdTrpA1と可視マーカーとを発現していたとすれば,それは求愛行動を起こす能力をもたない細胞であると考えられる.すでに筆者らは,脳のfru発現ニューロンの多くを同定し約50のクラスターに分類している7,8).高温で求愛行動を示したグループと示さなかったグループとを比較したとき,前者で有意に高くdTrpA1を発現していたfru発現ニューロンこそが求愛行動を発動させる元締めであろう.
行動要素としてタッピングとシンギングに着目してこの解析を行ったところ,fru発現ニューロンのうちP1クラスターとP2bクラスターの2つのグループが両方の行動要素の誘起に共通して有意の相関を示した.P1クラスターは脳半球あたり20個の介在ニューロンで構成され,その神経突起は正中線をこえて反対側に伸び左右の外側原大脳の神経叢をつないでいる.すでに筆者らは,P1クラスターが雄だけにしかないニューロングループであること,および,P1クラスターを雌の脳に人為的につくりだすとその雌が雄の求愛行動を示すことを明らかにしている8).一方,P2bクラスターは,今回,はじめて性行動への関与が認められたものであった.P2bクラスターを構成する介在ニューロンは外側原大脳に発し腹髄に伸びる長い軸索をもち,胸部3体節の神経節に終末を形成している.P1クラスターとP2bクラスターのニューロンは外側原大脳において互いの突起分枝を同じ領域に広げており,シナプス接続を形成していることが推察された.これらの観察から,P1クラスターが求愛行動の引き金をひく中枢を構成し,P2bクラスターはP1クラスターの“決定”を胸部の運動中枢に伝える下行性の司令線維である可能性が浮上してきた.
ここに仮定したようにP1クラスターが求愛の引き金をひく中枢であるならば,このニューロングループは求愛を開始させる刺激により興奮し,求愛活動を低下させる刺激により抑制をうけるはずである.これを検証するには,雄の求愛行動をひき起こす刺激,つまり,雌そのものに対してP1クラスターのニューロンがどのように反応するかを調べる必要がある.この実験のためには相反する2つの条件がみたされなければならない.脳のニューロン活動を記録するためには,脳が動かないことが必要である.その一方で,雄は求愛という行動,つまり,動きを示すことを求められる.そこで,この2つの要求を同時にみたすパラダイムを開発した.
まず,雄を固定した状態で求愛行動をとらせるシステムでは,雄の胸部背板を銅線に貼り付けて固定した.脚には発泡スチロールの球を持たせ,下からの気流で球を安定させた.この拘束した雄の前脚に雌の腹部で触れると,雄は提示された雌の腹部を追跡しはじめ,片翅を振動させた(シンギング).ただし,実際には体の位置は一定であり雄は球を回しているだけである(図2).球の回転速度と方向から雄の“主観的な動き”(追っかけている“つもり”)を自動的に計測した.接触をさせず雌の腹部を動かしながら雄に提示した場合には求愛行動は起こらなかった.また,雄の腹部を触らせても求愛行動は起こらなかった.ショウジョウバエの雄に固有の求愛抑制フェロモンであるcis-バクセン酸アセテート11)(cVA)を処女雌に塗布しその腹部を触らせたときにも求愛行動はひき起こされなかった.これらの結果から,雄は雌の体表の化学成分をタッピングにより感知し,その受容によって求愛行動が開始されるものと考えられた.

つぎに,行動中にニューロンの活動を記録するシステムでは,フレームに貼ったプラスティックフィルムに胸部背板を固定し,頭部クチクラの背面に窓を開けて脳を“透視”できるよう改変した.そして,fru発現ニューロンにCa2+感受性タンパク質のYellow Cameleon(YC2.1)を強制発現させ,ニューロンのCa2+濃度の変化を蛍光イメージングにより記録した.その結果,雄が雌に触ると外側原大脳のニューロンが一過性のCa2+上昇で応答することが明らかとなった.雄やcis-バクセン酸アセテートを塗布した処女雌に触ったときにも外側原大脳はCa2+上昇を示すが,その応答は弱い.一方,ガラス棒で触ったときにはほとんど応答しないが,ガラス棒にショウジョウバエのヘキサン抽出物を塗布すると大きな応答を発生させることができた.このように,外側原大脳には求愛行動を開始させる中枢がもつと期待される特性を示すニューロンが存在していた.
このニューロンを同定するため,MARCM法によりYellow Cameleonの発現を少数のニューロンに限定し,雌の腹部で刺激したときの応答を記録したのち,Yellow Cameleonに対する抗体染色によりそのニューロンを同定した.その結果,雌への接触により一過性のCa2+上昇を示した6個体の事例がすべてP1クラスターの活動記録であることが判明した.こうして,P1クラスターが求愛行動の鍵刺激に応答すること,P1クラスターの強制活性化により雄に求愛行動を誘起できることが実証された.
この研究により,20個の雄特異的な介在ニューロンからなるP1クラスターが雄の性行動の引き金をひく中枢を構成することが明らかとなった.その活動記録から,これらのニューロンは実際には求愛行動をひき起こさない刺激に対しても応答を示すことが示された.おそらく,P1クラスターは活動電位の発生閾値の高いニューロンからなり,多種の感覚性入力の統合によってはじめて発火して求愛行動の開始の引き金をひくのであろう.その情報処理のしくみと意志決定のロジックを解明するため,今後は,行動中の個体で実際に生起するニューロンの活動を電気生理学的に記録し,シナプス統合の動態から行動を理解する研究の展開が望まれる.
東北大学大学院生命科学研究科 研究員.
小金澤 雅之(Masayuki Koganezawa)
東北大学大学院生命科学研究科 准教授.
山元 大輔(Daisuke Yamamoto)
略歴:1978年 東京農工大学大学院農学研究科修士課程 修了,1980年 三菱化学生命科学研究所 研究員,1981年 理学博士(北海道大学),1981~1983年 米国Northwestern大学School of Medicine博士研究員,1999年 早稲田大学人間科学部 教授を経て,2005年より東北大学大学院生命科学研究科 教授.
研究テーマ:ショウジョウバエ性行動の遺伝子および細胞機構と進化.
© 2011 古波津 創・小金澤雅之・山元大輔 Licensed under CC 表示 2.1 日本
(東北大学大学院生命科学研究科 脳機能遺伝分野)
email:山元大輔
DOI: 10.7875/first.author.2011.030
Female contact activates male-specific interneurons that trigger stereotypic courtship behavior in Drosophila.
Soh Kohatsu, Masayuki Koganezawa & Daisuke Yamamoto
Neuron, 69, 498-508 (2011)
要 約
ショウジョウバエのfruitless遺伝子は雄の性行動に必要な神経回路の主要部を組み立てるはたらきをもつ.高温になると開口するdTrpA1チャネルをfru発現ニューロンに限定して発現させ人工的にこのニューロンを興奮させたところ,雄は相手となる雌のいない容器内で定型的な求愛行動を実行した.少数の細胞にだけdTrpA1を発現させる実験から雄の性行動を開始させる能力のある細胞としてP1クラスターとP2bクラスターを同定した.拘束した条件で行動させ,同時に,ニューロンの活動をCa2+イメージングにより記録する実験により,雄が前脚で雌の腹部を触ってフェロモンを感知するとP1クラスターが一過性に興奮することが判明した.したがって,P1クラスターは定型的な求愛行動を開始させる中枢をなすものと考えられた.
はじめに
動物の本能行動が神経系においていかにかたちづくられ,数あるレパートリーのなかから適切な行動が状況に応じどのように選択され実行されるのかという問題は,動物行動学が解決すべき主要なテーマのひとつである.Tinbergenは,さまざまな動物の行動観察から,行動のレパートリーごとに固有の中枢が存在し,それらのあいだでの協調的あるいは競合的な神経接続を介して行動の選択がなされると想定した1).行動中枢はより下位の中枢を階層的に活性化して,選択されたひとつのレパートリーを構成する行動要素のそれぞれをつくりだし,最後には定型的な運動出力を自動的に生み出すとされる.これが生得的解発機構(innate releasing mechanism:IRM)であり,これを起動させる特異的な感覚刺激が鍵刺激(releaser)である.
Wiersma & Ikedaは,ザリガニの遊泳肢の体節をこえた協調的な運動をささえる神経機構の分析を試み,腹髄のあるニューロン軸索に緊張性の発火が生じさえすればこの運動が完璧なかたちで実行されることを示し,そのニューロン軸索を司令線維とよんだ2).司令線維は行動中枢の決定を下位の運動中枢へと伝えるはたらきをしているものと考えられる.そののち,コオロギを用いた一連の研究から司令線維の発火頻度の違いにより異なる歌のレパートリーのひき起こされることが示されている3).しかし,司令線維を駆動する上位の行動中枢の実体は明確になっておらず,司令線維そのものの同定も多くの場合はあいまいなままであった.
行動の選択と制御の中枢機構を解明するためには,これらの司令的な機能をもつニューロンの同定と,その活動の人為的な操作が不可欠である.キイロショウジョウバエを用いると遺伝的な手法により個々のニューロンの標識と操作とが可能である.この昆虫の性行動はきわめて定型的で要素的な行動を明確に定義できることから,Tinbergenが想定した生得的解発機構の実体を解明する好個の素材となる.この研究の目的は,ショウジョウバエを用いて求愛の行動中枢を同定することにあった.
1.性行動を制御する神経回路の形成を支配するfruitless遺伝子
キイロショウジョウバエの性行動は以下の要素から成り立っている.まず,雄が雌にむかって定位(orientation),接近し,前脚で雌の腹部を叩いて(タッピング,tapping)追跡しながら,片方の翅を打ち振るわせてラブソングを奏でる(シンギング,singing).雌の動きが遅くなると雄は背後にまわり口吻で雌の交尾器をなめる(リッキング,licking).つづいて,腹部を内側に曲げて雌にマウントしようとする(交尾試行,attempted copulation).雌が性的に受け入れる“気持ち”になると膣口を開き翅を立てて交尾(copulation)を可能にする.交尾の受け入れ以外のこれらの行動はすべて雄に特異的であり,野生型の雌は決してこの一連の行動をとることはない.野生型の雄は,通常,雌に対して求愛行動をとり,求愛対象がいなければ決してこの行動はせず,また,雄に対しても持続的に求愛をすることはない.
しかし,ある遺伝子の機能が低下すると,雄が雄に求愛したり,雌にも雄にも求愛しなくなったりといった行動の変化が起こる.その遺伝子はfruitless(fru)の名で知られている4,5).fru遺伝子の1次転写産物は性によって異なるスプライシングをうけ,翻訳の起こるのは雄の神経系のみであり,雌の神経系はFruタンパク質を欠く.fru遺伝子の改変により雌雄いずれでもその翻訳の起こるfruM変異体が,相同組換えによるノックイン法を用いて作製されている.注目すべきことに,fruMホモ接合体の雌個体はほかの雌に対して雄の求愛行動を示す6).つまり,fruM遺伝子を発現するニューロンがFruタンパク質をもつ場合に雄の行動が生じることになる.このことは,Fruタンパク質が神経系の雄化因子であることを示唆している.ただし,fru遺伝子がおもにはたらくのは成虫の神経系のかたちづくられる蛹期であり,成虫となって実際に求愛する際にはもはや必要でないと考えられている.実際,fru発現ニューロンのなかには雄特異的に存在するものや性的二型を示すものがあるが7,8),そうした性差は蛹期に形成されるもので成虫羽化ののち性により変化することはないと思われる.
2.ニューロンの活動の人為的な活性化による性行動の誘導
ショウジョウバエの脳には105個ほどのニューロンがあるとされるが,fru遺伝子を発現するのはそのうちの約2000個である.これまで述べた知見から,雄の求愛行動を制御するニューロンはこの2000個に含まれているものと期待された.この仮説を検証するため,雄の脳のfru発現ニューロンを強制的に活性化させることによって求愛行動を人工的にひき起こすことが可能かどうかを調べた.ニューロンの強制的な活性化を惹起するツールとして,温度感受性イオンチャネルのdTrpA1を用いた.dTrpA1は高温で開口して陽イオンの流入をひき起こしニューロンを興奮させる9).dTrpA1をfru発現ニューロンにのみ発現させる方策としてGal4-UAS系を活用した.Gal4は酵母の転写因子で,UASはその結合標的配列である.UAS-dTrpA1遺伝子を導入した個体とfru発現細胞でGal4を発現するfruNP21系統個体とを掛け合わせて得た個体では,Gal4タンパク質をもつ細胞,すなわち,fru発現ニューロンにおいてのみUAS-dTrpA1遺伝子が転写される.その結果,fru発現ニューロンはdTrpA1をもつことになり,高温条件ではその開口により興奮を起こす.このような雄を単独でチェンバーに入れ観察すると,20℃では普通の雄と同じく容器内を歩き回るばかりであるが,26~30℃に温度を上げると,驚くべきことに相手がいないにもかかわらず,一連の求愛行動をくり返し示すことがわかった(図1).
3.遺伝的モザイクを用いた行動制御ニューロンの同定
fruNP21系統ではfru発現細胞の約80%にGal4が発現する.このうちどの細胞が求愛行動の惹起に寄与するのかを明らかにするため,MARCM(mosaic analysis with a repressible cell marker)法10) によってdTrpA1をより少数のfru発現細胞(たとえば,数十個)に限定して発現させ強制活性化を行うことにした.MARCM法は発生の過程で一部の細胞に体細胞染色体組換えを誘導するもので,これを利用すると,組換えが起こった細胞やその子孫細胞にのみdTrpA1を発現させることができる.dTrpA1をもつ細胞は同時に可視マーカー(mCD8::GFP)を発現するよう仕組んでおき,行動実験のあと各個体から脳を摘出しマーカーを発現する細胞を抗GFP抗体により染色し同定する.高温で求愛行動を示す個体では求愛行動をひき起こす能力のあるニューロンにdTrpA1が発現しているはずであり,そのニューロンは可視マーカーで標識される.一方,高温にしても求愛行動を示さない個体においてあるニューロンがdTrpA1と可視マーカーとを発現していたとすれば,それは求愛行動を起こす能力をもたない細胞であると考えられる.すでに筆者らは,脳のfru発現ニューロンの多くを同定し約50のクラスターに分類している7,8).高温で求愛行動を示したグループと示さなかったグループとを比較したとき,前者で有意に高くdTrpA1を発現していたfru発現ニューロンこそが求愛行動を発動させる元締めであろう.
4.求愛行動を開始させるP1クラスターとP2bクラスター
行動要素としてタッピングとシンギングに着目してこの解析を行ったところ,fru発現ニューロンのうちP1クラスターとP2bクラスターの2つのグループが両方の行動要素の誘起に共通して有意の相関を示した.P1クラスターは脳半球あたり20個の介在ニューロンで構成され,その神経突起は正中線をこえて反対側に伸び左右の外側原大脳の神経叢をつないでいる.すでに筆者らは,P1クラスターが雄だけにしかないニューロングループであること,および,P1クラスターを雌の脳に人為的につくりだすとその雌が雄の求愛行動を示すことを明らかにしている8).一方,P2bクラスターは,今回,はじめて性行動への関与が認められたものであった.P2bクラスターを構成する介在ニューロンは外側原大脳に発し腹髄に伸びる長い軸索をもち,胸部3体節の神経節に終末を形成している.P1クラスターとP2bクラスターのニューロンは外側原大脳において互いの突起分枝を同じ領域に広げており,シナプス接続を形成していることが推察された.これらの観察から,P1クラスターが求愛行動の引き金をひく中枢を構成し,P2bクラスターはP1クラスターの“決定”を胸部の運動中枢に伝える下行性の司令線維である可能性が浮上してきた.
5.拘束した個体を用いた求愛行動の定量化
ここに仮定したようにP1クラスターが求愛の引き金をひく中枢であるならば,このニューロングループは求愛を開始させる刺激により興奮し,求愛活動を低下させる刺激により抑制をうけるはずである.これを検証するには,雄の求愛行動をひき起こす刺激,つまり,雌そのものに対してP1クラスターのニューロンがどのように反応するかを調べる必要がある.この実験のためには相反する2つの条件がみたされなければならない.脳のニューロン活動を記録するためには,脳が動かないことが必要である.その一方で,雄は求愛という行動,つまり,動きを示すことを求められる.そこで,この2つの要求を同時にみたすパラダイムを開発した.
まず,雄を固定した状態で求愛行動をとらせるシステムでは,雄の胸部背板を銅線に貼り付けて固定した.脚には発泡スチロールの球を持たせ,下からの気流で球を安定させた.この拘束した雄の前脚に雌の腹部で触れると,雄は提示された雌の腹部を追跡しはじめ,片翅を振動させた(シンギング).ただし,実際には体の位置は一定であり雄は球を回しているだけである(図2).球の回転速度と方向から雄の“主観的な動き”(追っかけている“つもり”)を自動的に計測した.接触をさせず雌の腹部を動かしながら雄に提示した場合には求愛行動は起こらなかった.また,雄の腹部を触らせても求愛行動は起こらなかった.ショウジョウバエの雄に固有の求愛抑制フェロモンであるcis-バクセン酸アセテート11)(cVA)を処女雌に塗布しその腹部を触らせたときにも求愛行動はひき起こされなかった.これらの結果から,雄は雌の体表の化学成分をタッピングにより感知し,その受容によって求愛行動が開始されるものと考えられた.
6.行動中の個体からニューロンの活動を記録する
つぎに,行動中にニューロンの活動を記録するシステムでは,フレームに貼ったプラスティックフィルムに胸部背板を固定し,頭部クチクラの背面に窓を開けて脳を“透視”できるよう改変した.そして,fru発現ニューロンにCa2+感受性タンパク質のYellow Cameleon(YC2.1)を強制発現させ,ニューロンのCa2+濃度の変化を蛍光イメージングにより記録した.その結果,雄が雌に触ると外側原大脳のニューロンが一過性のCa2+上昇で応答することが明らかとなった.雄やcis-バクセン酸アセテートを塗布した処女雌に触ったときにも外側原大脳はCa2+上昇を示すが,その応答は弱い.一方,ガラス棒で触ったときにはほとんど応答しないが,ガラス棒にショウジョウバエのヘキサン抽出物を塗布すると大きな応答を発生させることができた.このように,外側原大脳には求愛行動を開始させる中枢がもつと期待される特性を示すニューロンが存在していた.
このニューロンを同定するため,MARCM法によりYellow Cameleonの発現を少数のニューロンに限定し,雌の腹部で刺激したときの応答を記録したのち,Yellow Cameleonに対する抗体染色によりそのニューロンを同定した.その結果,雌への接触により一過性のCa2+上昇を示した6個体の事例がすべてP1クラスターの活動記録であることが判明した.こうして,P1クラスターが求愛行動の鍵刺激に応答すること,P1クラスターの強制活性化により雄に求愛行動を誘起できることが実証された.
おわりに
この研究により,20個の雄特異的な介在ニューロンからなるP1クラスターが雄の性行動の引き金をひく中枢を構成することが明らかとなった.その活動記録から,これらのニューロンは実際には求愛行動をひき起こさない刺激に対しても応答を示すことが示された.おそらく,P1クラスターは活動電位の発生閾値の高いニューロンからなり,多種の感覚性入力の統合によってはじめて発火して求愛行動の開始の引き金をひくのであろう.その情報処理のしくみと意志決定のロジックを解明するため,今後は,行動中の個体で実際に生起するニューロンの活動を電気生理学的に記録し,シナプス統合の動態から行動を理解する研究の展開が望まれる.
文 献
- Tinbergen, N.: The Study of Instinct. Clarendon Press, Oxford (1951) [邦訳] ニコラス・ティンバーゲン 著,永野為武 訳,本能の研究. 三共出版 (1975)
- Wiersma, C. A. & Ikeda, K.: Interneurons commanding swimmeret movements in the crayfish, Procambarus clarki (Girard). Comp. Biochem. Physiol., 12, 509-525 (1964)[PubMed]
- Bentley, D.: Control of cricket song patterns by descending interneurons. J. Comp. Physiol., 116, 19-38 (1977)
- Ito, H., Fujitani, K., Usui, K. et al.: Sexual orientation in Drosophila is altered by the satori mutation in the sex-determination gene fruitless that encodes a zinc finger protein with a BTB domain. Proc. Natl. Acad. Sci. USA, 93, 9687-9692 (1996)[PubMed]
- Ryner, L. C., Goodwin, S. F., Castrillon, D. H. et al.: Control of male sexual behavior and sexual orientation in Drosophila by the fruitless gene. Cell, 87, 1079-1089 (1996)[PubMed]
- Demir, E. & Dickson, B. J.: fruitless splicing specifies male courtship behavior in Drosophila. Cell, 121, 785-794 (2005)[PubMed]
- Kimura, K. -I., Ote, M., Tazawa, T. et al.: Fruitless specifies sexually dimorphic neural circuitry in the Drosophila brain. Nature, 438, 229-233 (2005)[PubMed]
- Kimura, K. -I., Hachiya, T., Koganezawa, M. et al.: Fruitless and doublesex coordinate to generate male-specific neurons that can initiate courtship. Neuron, 59, 759-769 (2008)[PubMed]
- Hamada, F. N., Rosenzweig, M., Kang, K. et al.: An internal thermal sensor controlling temperature preference in Drosophila. Nature, 454, 217-220 (2008)[PubMed]
- Lee, T. & Luo, L.: Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron, 22, 451-461 (1999)[PubMed]
- Jallon, J. -M., Antony, C. & Benamar, O.: Un anti-aphrodisiawue produit par les males Drosophila melanogaster et transfere aux femelles lors de la copulation. C R. Acad. Sci. Paris, 292, 1147-1149 (1981)
著者プロフィール
東北大学大学院生命科学研究科 研究員.
小金澤 雅之(Masayuki Koganezawa)
東北大学大学院生命科学研究科 准教授.
山元 大輔(Daisuke Yamamoto)
略歴:1978年 東京農工大学大学院農学研究科修士課程 修了,1980年 三菱化学生命科学研究所 研究員,1981年 理学博士(北海道大学),1981~1983年 米国Northwestern大学School of Medicine博士研究員,1999年 早稲田大学人間科学部 教授を経て,2005年より東北大学大学院生命科学研究科 教授.
研究テーマ:ショウジョウバエ性行動の遺伝子および細胞機構と進化.
© 2011 古波津 創・小金澤雅之・山元大輔 Licensed under CC 表示 2.1 日本
