ホヤの変態において上衣細胞が成体のニューロンをつくりだす
堀江健生・笹倉靖徳
(筑波大学下田臨海実験センター 発生遺伝学グループ)
email:堀江健生,笹倉靖徳
DOI: 10.7875/first.author.2011.020
Ependymal cells of chordate larvae are stem-like cells that form the adult nervous system.
Takeo Horie, Ryoko Shinki, Yosuke Ogura, Takehiro G. Kusakabe, Nori Satoh, Yasunori Sasakura
Nature, 469, 525-528 (2011)
ホヤは変態によって幼生から成体へと劇的に体制を変化させる.このとき,幼生の中枢神経系が制御している幼生の器官の多くは変態の過程で失われ,反対に新しく成体の器官が形成される.この変化に対応するため,成体の中枢神経系は幼生のものとは大きく構成を変える必要がある.ホヤ成体の中枢神経系が変態においてどのように再構成されるかについては,幼生の中枢神経系を構成する細胞のほぼすべてがいちど失われて,変態の過程で成体の中枢神経系が新規につくりだされるものと考えられてきた.しかしながら,変態の過程において幼生の中枢神経系を正確に追跡した実験はこれまで行われてこなかったため,その説の真偽が議論となっていた.そのことを明確にするため,筆者らは,変態の過程において幼生の中枢神経系の追跡実験を行った.その結果,ホヤ幼生の中枢神経系の大部分は変態ののちにも維持され成体の中枢神経系を構築することが明らかにされた.また,その際に幼生のニューロンのほとんどは従来の説のとおりに失われるが,グリア細胞である上衣細胞が主として残り成体の中枢神経系を再構築すること,その一部が成体のニューロンを生みだしていることを明らかにした.この結果は,上衣細胞は神経幹細胞様の性質を備えており,脊索動物の変態において成体の中枢神経系を再構築するために利用されていることを示唆している.
ホヤは脊椎動物と同じ脊索動物門に属する海産の無脊椎動物であり,古くから発生学や進化生物学の研究材料として用いられている1).幼生は頭部と尾部からなるオタマジャクシ型の形態をしており,背側に中枢神経系をもつなど脊椎動物と共通の体制を備えている.筆者らが研究材料に用いているカタユウレイボヤは受精後およそ18時間(18℃)で孵化しオタマジャクシ型幼生となる.幼生は遊泳生活をおくり,孵化後およそ10時間で岸壁などに固着し,数日のあいだに変態を行い大きく体制を変化させて固着性の成体になる.ホヤ幼生の中枢神経系は前方部がふくらんだ脳のような構造をもち,背側に神経管が位置するなど脊椎動物の脳神経系の基本設計が単純なかたちでみられるが,約100個のニューロンと約250個のグリア細胞という非常に少数の細胞から構成されている2).そのため,脊索動物におけるニューロンの機能やその分化機構を研究するのに適したモデルである3).
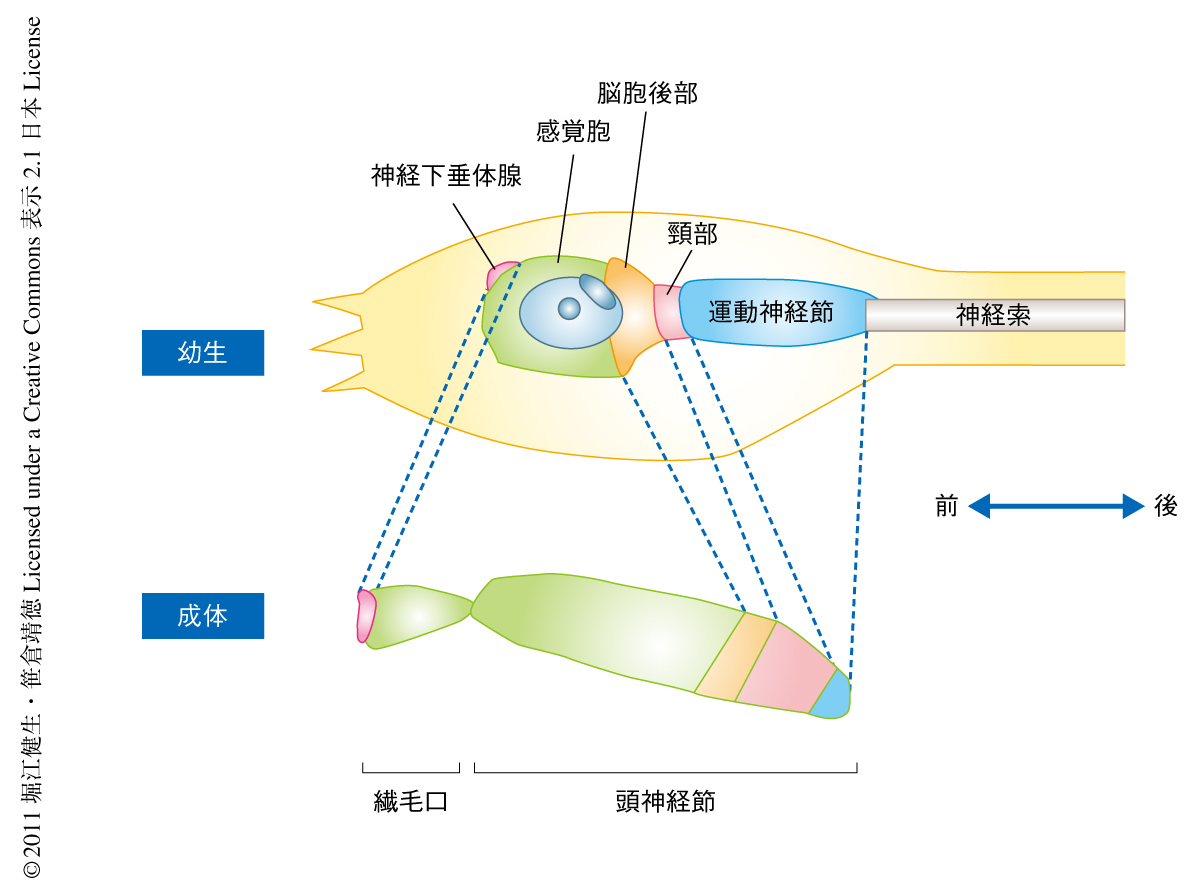
ホヤ幼生の中枢神経系は,前方部から感覚胞(sensory vesicle),脳胞後部(posterior brain),頸部(neck),運動神経節(visceral ganglion),神経索(nerve cord)と大きく5つの部分に分けることができる(図1).感覚胞は眼点および平衡器という2つの感覚器官をもっており,重力や光などを感知して遊泳運動を制御している.脳胞後部は感覚器官からの入力をうけその情報を運動ニューロンへと伝える感覚情報の統合中心として機能するものと考えられている.運動神経節には運動ニューロンが存在しており,尾部の筋肉の収縮の制御を介して運動を制御している.神経索は尾部に存在しており,頭部から伸びた2本の軸索が走行している.また,中枢神経系の全体はグリア細胞の一種である上衣細胞によりおおわれている.このように,幼生の神経系は遊泳運動に特化した構造と機能を備えている.一方,固着生活をする成体は当然ではあるが尾部をもたず遊泳もしない.成体の中枢神経系のおもな役割は餌を採取するエラの繊毛運動の調節や生殖の制御などであり,幼生の神経系の機能とは大きく異なっている.したがって,体全体の構造を変化させる変態の過程において,遊泳運動に特化した幼生の神経系はいちどその役割をおえて,新たに成体の神経系が再構築される必要がある.
この変態のときの中枢神経系の再構築の過程において,幼生の中枢神経系を構成するほぼすべての細胞は細胞死によって消失し,成体の中枢神経系は幼生の中枢神経系のごく一部である神経下垂体腺とよばれる領域から新たに構築されることが組織学的な観察結果から報告されていた4,5).その一方で,蛍光タンパク質YFPをレポーターとして幼生の神経系の別の領域の細胞に発現させてその運命を追跡した実験結果から,幼生の神経系のより多くの細胞が変態ののちにも維持されるという報告もあり6),幼生の神経系の細胞が残るのか失われるのかが大きな論点となっていた.その原因として,幼生の中枢神経系を全体にわたって正確に標識し直接に追跡する実験の行われていなかったことがあげられる.たとえば,さきに述べたYFPを使った実験では,YFPを発現する幼生の細胞が変態ののちにも本当に残っている可能性と,変態の途中でYFPを発現する細胞が新たに形成される可能性とを区別することはできない.
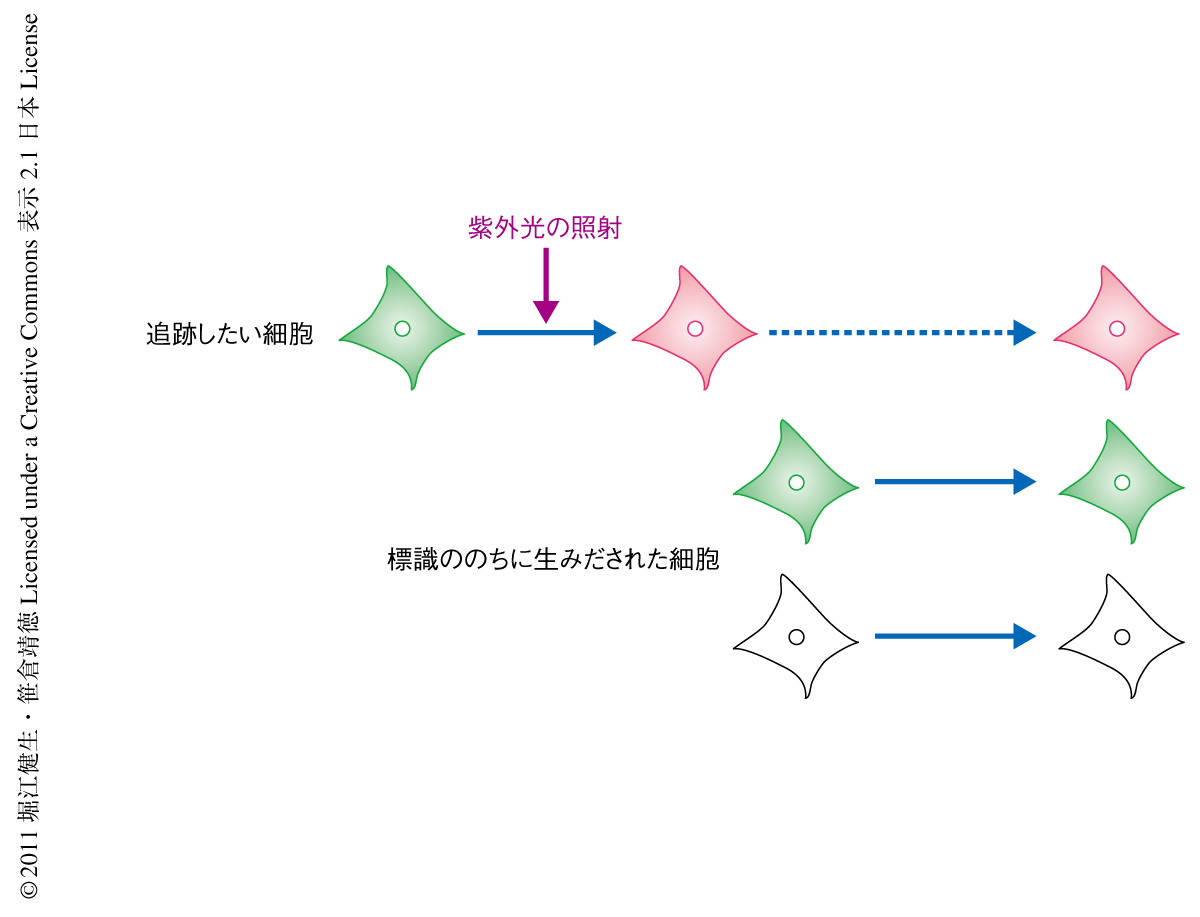
そこで筆者らは,光変換型蛍光タンパク質Kaedeを利用して幼生の中枢神経系の細胞のみを特異的に標識し,変態ののちの成体の神経系の構築に関与しているのかどうかについて調べることにした.Kaedeはサンゴから発見された蛍光タンパク質で,通常は緑色の蛍光を発しているが紫外光の照射により赤色へとその蛍光を変化させる7).この変化は不可逆的であるためKaedeを発現させた細胞に紫外光を照射することでその細胞を赤色蛍光で標識することができ,その細胞の運命を正確に追跡することが可能である(図2).この手法を用いて,変態の過程におけるホヤの中枢神経系の細胞の追跡実験を行った.
具体的な実験はつぎのとおりである.まず,幼生の中枢神経系のほぼすべての細胞でKaedeを発現するトランスジェニック系統を作製した.つづいて,この系統の幼生に紫外光を照射することにより幼生の中枢神経系に存在する細胞のみを赤色蛍光で標識し,変態を完了した成体において中枢神経系が幼生の中枢神経系に由来する赤色の細胞をもっているかどうかを調べた.その結果,幼生の神経系の細胞は失われるという従来の説とは異なり,赤色蛍光で標識された幼生の中枢神経系に由来する細胞は成体の中枢神経系の全体に分布していることが明らかになった.つぎに,幼生の中枢神経系のどの領域が成体の中枢神経系を形成するのかを局所的な標識により調べた.具体的には,Kaedeトランスジェニック系統の幼生に対して,中枢神経系のそれぞれの領域に局所的に紫外光を照射し,その領域に存在する細胞のみを特異的に標識して追跡した.その結果,尾部にある神経索を除く幼生の中枢神経系のほとんどの部分が変態ののちにも維持されて成体の中枢神経系を構築することが明らかになった.また,幼生の中枢神経系の前方領域から成体の中枢神経系の前方領域が,幼生の中枢神経系の後方領域から成体の中枢神経系の後方領域が形成されること,すなわち,変態ののちの成体の中枢神経系においても幼生の中枢神経系の前後軸パターンの反映されることが明らかになった(図1).
ホヤ幼生の中枢神経系は約100個のニューロンと約250個のグリア細胞から構成されている2).ホヤのニューロンはニューロンが利用する神経伝達物質により分類されており,中枢神経系にはおもにグルタミン酸作動性ニューロン,GABA/グリシン作動性ニューロン,コリン作動性ニューロンが存在している8).グリア細胞としては神経管を裏打ちする細胞である上衣細胞のみが存在していることが報告されている2).そこで,それらの細胞タイプのすべてが変態の過程で残るのか一部のみが残るのかを明らかにすることを試みた.まず,ニューロンについて追跡した.それぞれの神経伝達物質作動性ニューロンにおいて特異的にKaedeを発現するトランスジェニック系統を作製しどのニューロンが変態ののちにも維持されるのかについて追跡実験を行ったところ,一部のコリン作動性ニューロンとグルタミン酸作動性ニューロンが維持されたものの,幼生のニューロンの大部分は変態の途中に消失することが明らかになった.つづいて,グリア細胞である上衣細胞についても追跡実験を行ったところ,赤色蛍光で標識された幼生の上衣細胞の多くは変態ののちにも維持され,成体の中枢神経系の全体に存在していることが明らかになった.また,幼生の上衣細胞の一部は変態ののちに軸索を伸ばしており,変態の過程でニューロンへと分化している可能性が示唆された.
この結果から,幼生の上衣細胞から成体のニューロンの生みだされていることが示唆された.そこで,その可能性を検証するため,上衣細胞でKaedeを発現し,さらに,コリン作動性ニューロンで蛍光タンパク質CFPを発現するトランスジェニック個体を作製した.この個体に対して幼生期に紫外光を照射し幼生の上衣細胞を赤色蛍光で特異的に標識した.そして,赤色蛍光で標識された上衣細胞が変態ののちにCFPを発現するコリン作動性ニューロンに分化するかどうかを検討した.その結果,幼生の上衣細胞の一部が変態ののちCFPを発現していること,つまり,成体のコリン作動性ニューロンへと分化していることが明らかになった.成体のGABA/グリシン作動性ニューロンについても同様の実験を行い,幼生の上衣細胞から生みだされていることを確認することができた.これらの結果から,幼生の上衣細胞は少なくとも成体のコリン作動性ニューロンとGABA/グリシン作動性ニューロンを生みだすことのできることが明らかになり,幼生の上衣細胞が変態の過程でニューロンを生みだす神経幹細胞様の性質を備えている可能性が示唆された.
さきに述べたとおり,過去の組織学的な知見より幼生の神経下垂体腺から成体の中枢神経系の生みだされることが報告されていたことをうけて,幼生の神経下垂体腺が成体の中枢神経系を構成するかどうかについて調べた.幼生の神経下垂体腺で特異的に発現する遺伝子の発現調節領域を利用して神経下垂体腺でのみKaedeを発現するトランスジェニック個体を得た.この個体に紫外光を照射し特異的に標識することによって神経下垂体腺に存在する細胞を追跡した.その結果,神経下垂体腺に存在する細胞は変態ののちに成体の中枢神経系のごく一部分,繊毛口の前方領域を構成するが,成体の中枢神経系の全体は生みださないことが明らかになった(図1).
高等脊椎動物である哺乳類の成体の中枢神経系にも神経幹細胞が存在し多様なニューロンを生みだしていることが報告されている.その哺乳類の神経幹細胞はグリア細胞と共通した性質を備えていることも報告されている.とくに,上衣細胞は神経幹細胞であると報告されたこともあり,神経幹細胞を理解するうえで重要な細胞となっている9,10).今回,脊椎動物にもっとも近い無脊椎動物であるホヤの上衣細胞が神経幹細胞様の性質を備えていることが明らかにされた.このことにより脊椎動物の神経幹細胞やその進化的な起源について新たな知見をあたえることができたと考えている.今後,ホヤ幼生の上衣細胞が真に神経幹細胞としての形質と機能を備えているかどうかについての検証を行うとともに,変態の過程において上衣細胞からニューロンを生みだす分子機構を解明することで神経幹細胞からのニューロンを生みだす脊索動物の全体に普遍的な機構について解明したいと考えている.
略歴:2007年 兵庫県立大学大学院生命理学研究科 修了,同年より筑波大学下田臨海実験センター 研究員.
研究テーマ:ホヤ幼生の神経回路網の機能とその分化機構の解明.
抱負:単純なホヤの中枢神経系の研究から,複雑な脊椎動物の中枢神経系を理解するための基本原理を明らかにしたいと考えています.
笹倉 靖徳(Sasakura Yasunori)
筑波大学下田臨海実験センター 准教授.
研究室URL:http://www.shimoda.tsukuba.ac.jp/~sasakura/index.html
© 2011 堀江健生・笹倉靖徳 Licensed under CC 表示 2.1 日本
(筑波大学下田臨海実験センター 発生遺伝学グループ)
email:堀江健生,笹倉靖徳
DOI: 10.7875/first.author.2011.020
Ependymal cells of chordate larvae are stem-like cells that form the adult nervous system.
Takeo Horie, Ryoko Shinki, Yosuke Ogura, Takehiro G. Kusakabe, Nori Satoh, Yasunori Sasakura
Nature, 469, 525-528 (2011)
要 約
ホヤは変態によって幼生から成体へと劇的に体制を変化させる.このとき,幼生の中枢神経系が制御している幼生の器官の多くは変態の過程で失われ,反対に新しく成体の器官が形成される.この変化に対応するため,成体の中枢神経系は幼生のものとは大きく構成を変える必要がある.ホヤ成体の中枢神経系が変態においてどのように再構成されるかについては,幼生の中枢神経系を構成する細胞のほぼすべてがいちど失われて,変態の過程で成体の中枢神経系が新規につくりだされるものと考えられてきた.しかしながら,変態の過程において幼生の中枢神経系を正確に追跡した実験はこれまで行われてこなかったため,その説の真偽が議論となっていた.そのことを明確にするため,筆者らは,変態の過程において幼生の中枢神経系の追跡実験を行った.その結果,ホヤ幼生の中枢神経系の大部分は変態ののちにも維持され成体の中枢神経系を構築することが明らかにされた.また,その際に幼生のニューロンのほとんどは従来の説のとおりに失われるが,グリア細胞である上衣細胞が主として残り成体の中枢神経系を再構築すること,その一部が成体のニューロンを生みだしていることを明らかにした.この結果は,上衣細胞は神経幹細胞様の性質を備えており,脊索動物の変態において成体の中枢神経系を再構築するために利用されていることを示唆している.
はじめに
ホヤは脊椎動物と同じ脊索動物門に属する海産の無脊椎動物であり,古くから発生学や進化生物学の研究材料として用いられている1).幼生は頭部と尾部からなるオタマジャクシ型の形態をしており,背側に中枢神経系をもつなど脊椎動物と共通の体制を備えている.筆者らが研究材料に用いているカタユウレイボヤは受精後およそ18時間(18℃)で孵化しオタマジャクシ型幼生となる.幼生は遊泳生活をおくり,孵化後およそ10時間で岸壁などに固着し,数日のあいだに変態を行い大きく体制を変化させて固着性の成体になる.ホヤ幼生の中枢神経系は前方部がふくらんだ脳のような構造をもち,背側に神経管が位置するなど脊椎動物の脳神経系の基本設計が単純なかたちでみられるが,約100個のニューロンと約250個のグリア細胞という非常に少数の細胞から構成されている2).そのため,脊索動物におけるニューロンの機能やその分化機構を研究するのに適したモデルである3).
ホヤ幼生の中枢神経系は,前方部から感覚胞(sensory vesicle),脳胞後部(posterior brain),頸部(neck),運動神経節(visceral ganglion),神経索(nerve cord)と大きく5つの部分に分けることができる(図1).感覚胞は眼点および平衡器という2つの感覚器官をもっており,重力や光などを感知して遊泳運動を制御している.脳胞後部は感覚器官からの入力をうけその情報を運動ニューロンへと伝える感覚情報の統合中心として機能するものと考えられている.運動神経節には運動ニューロンが存在しており,尾部の筋肉の収縮の制御を介して運動を制御している.神経索は尾部に存在しており,頭部から伸びた2本の軸索が走行している.また,中枢神経系の全体はグリア細胞の一種である上衣細胞によりおおわれている.このように,幼生の神経系は遊泳運動に特化した構造と機能を備えている.一方,固着生活をする成体は当然ではあるが尾部をもたず遊泳もしない.成体の中枢神経系のおもな役割は餌を採取するエラの繊毛運動の調節や生殖の制御などであり,幼生の神経系の機能とは大きく異なっている.したがって,体全体の構造を変化させる変態の過程において,遊泳運動に特化した幼生の神経系はいちどその役割をおえて,新たに成体の神経系が再構築される必要がある.
この変態のときの中枢神経系の再構築の過程において,幼生の中枢神経系を構成するほぼすべての細胞は細胞死によって消失し,成体の中枢神経系は幼生の中枢神経系のごく一部である神経下垂体腺とよばれる領域から新たに構築されることが組織学的な観察結果から報告されていた4,5).その一方で,蛍光タンパク質YFPをレポーターとして幼生の神経系の別の領域の細胞に発現させてその運命を追跡した実験結果から,幼生の神経系のより多くの細胞が変態ののちにも維持されるという報告もあり6),幼生の神経系の細胞が残るのか失われるのかが大きな論点となっていた.その原因として,幼生の中枢神経系を全体にわたって正確に標識し直接に追跡する実験の行われていなかったことがあげられる.たとえば,さきに述べたYFPを使った実験では,YFPを発現する幼生の細胞が変態ののちにも本当に残っている可能性と,変態の途中でYFPを発現する細胞が新たに形成される可能性とを区別することはできない.
1.幼生の中枢神経系の細胞は変態ののちにも維持される
そこで筆者らは,光変換型蛍光タンパク質Kaedeを利用して幼生の中枢神経系の細胞のみを特異的に標識し,変態ののちの成体の神経系の構築に関与しているのかどうかについて調べることにした.Kaedeはサンゴから発見された蛍光タンパク質で,通常は緑色の蛍光を発しているが紫外光の照射により赤色へとその蛍光を変化させる7).この変化は不可逆的であるためKaedeを発現させた細胞に紫外光を照射することでその細胞を赤色蛍光で標識することができ,その細胞の運命を正確に追跡することが可能である(図2).この手法を用いて,変態の過程におけるホヤの中枢神経系の細胞の追跡実験を行った.
具体的な実験はつぎのとおりである.まず,幼生の中枢神経系のほぼすべての細胞でKaedeを発現するトランスジェニック系統を作製した.つづいて,この系統の幼生に紫外光を照射することにより幼生の中枢神経系に存在する細胞のみを赤色蛍光で標識し,変態を完了した成体において中枢神経系が幼生の中枢神経系に由来する赤色の細胞をもっているかどうかを調べた.その結果,幼生の神経系の細胞は失われるという従来の説とは異なり,赤色蛍光で標識された幼生の中枢神経系に由来する細胞は成体の中枢神経系の全体に分布していることが明らかになった.つぎに,幼生の中枢神経系のどの領域が成体の中枢神経系を形成するのかを局所的な標識により調べた.具体的には,Kaedeトランスジェニック系統の幼生に対して,中枢神経系のそれぞれの領域に局所的に紫外光を照射し,その領域に存在する細胞のみを特異的に標識して追跡した.その結果,尾部にある神経索を除く幼生の中枢神経系のほとんどの部分が変態ののちにも維持されて成体の中枢神経系を構築することが明らかになった.また,幼生の中枢神経系の前方領域から成体の中枢神経系の前方領域が,幼生の中枢神経系の後方領域から成体の中枢神経系の後方領域が形成されること,すなわち,変態ののちの成体の中枢神経系においても幼生の中枢神経系の前後軸パターンの反映されることが明らかになった(図1).
2.幼生のニューロンの大部分は変態において消失し成体の中枢神経系は幼生のグリア細胞から形成される
ホヤ幼生の中枢神経系は約100個のニューロンと約250個のグリア細胞から構成されている2).ホヤのニューロンはニューロンが利用する神経伝達物質により分類されており,中枢神経系にはおもにグルタミン酸作動性ニューロン,GABA/グリシン作動性ニューロン,コリン作動性ニューロンが存在している8).グリア細胞としては神経管を裏打ちする細胞である上衣細胞のみが存在していることが報告されている2).そこで,それらの細胞タイプのすべてが変態の過程で残るのか一部のみが残るのかを明らかにすることを試みた.まず,ニューロンについて追跡した.それぞれの神経伝達物質作動性ニューロンにおいて特異的にKaedeを発現するトランスジェニック系統を作製しどのニューロンが変態ののちにも維持されるのかについて追跡実験を行ったところ,一部のコリン作動性ニューロンとグルタミン酸作動性ニューロンが維持されたものの,幼生のニューロンの大部分は変態の途中に消失することが明らかになった.つづいて,グリア細胞である上衣細胞についても追跡実験を行ったところ,赤色蛍光で標識された幼生の上衣細胞の多くは変態ののちにも維持され,成体の中枢神経系の全体に存在していることが明らかになった.また,幼生の上衣細胞の一部は変態ののちに軸索を伸ばしており,変態の過程でニューロンへと分化している可能性が示唆された.
3.幼生の上衣細胞が成体のニューロンを生みだす
この結果から,幼生の上衣細胞から成体のニューロンの生みだされていることが示唆された.そこで,その可能性を検証するため,上衣細胞でKaedeを発現し,さらに,コリン作動性ニューロンで蛍光タンパク質CFPを発現するトランスジェニック個体を作製した.この個体に対して幼生期に紫外光を照射し幼生の上衣細胞を赤色蛍光で特異的に標識した.そして,赤色蛍光で標識された上衣細胞が変態ののちにCFPを発現するコリン作動性ニューロンに分化するかどうかを検討した.その結果,幼生の上衣細胞の一部が変態ののちCFPを発現していること,つまり,成体のコリン作動性ニューロンへと分化していることが明らかになった.成体のGABA/グリシン作動性ニューロンについても同様の実験を行い,幼生の上衣細胞から生みだされていることを確認することができた.これらの結果から,幼生の上衣細胞は少なくとも成体のコリン作動性ニューロンとGABA/グリシン作動性ニューロンを生みだすことのできることが明らかになり,幼生の上衣細胞が変態の過程でニューロンを生みだす神経幹細胞様の性質を備えている可能性が示唆された.
4.幼生の神経下垂体腺は成体のニューロンにはならない
さきに述べたとおり,過去の組織学的な知見より幼生の神経下垂体腺から成体の中枢神経系の生みだされることが報告されていたことをうけて,幼生の神経下垂体腺が成体の中枢神経系を構成するかどうかについて調べた.幼生の神経下垂体腺で特異的に発現する遺伝子の発現調節領域を利用して神経下垂体腺でのみKaedeを発現するトランスジェニック個体を得た.この個体に紫外光を照射し特異的に標識することによって神経下垂体腺に存在する細胞を追跡した.その結果,神経下垂体腺に存在する細胞は変態ののちに成体の中枢神経系のごく一部分,繊毛口の前方領域を構成するが,成体の中枢神経系の全体は生みださないことが明らかになった(図1).
おわりに
高等脊椎動物である哺乳類の成体の中枢神経系にも神経幹細胞が存在し多様なニューロンを生みだしていることが報告されている.その哺乳類の神経幹細胞はグリア細胞と共通した性質を備えていることも報告されている.とくに,上衣細胞は神経幹細胞であると報告されたこともあり,神経幹細胞を理解するうえで重要な細胞となっている9,10).今回,脊椎動物にもっとも近い無脊椎動物であるホヤの上衣細胞が神経幹細胞様の性質を備えていることが明らかにされた.このことにより脊椎動物の神経幹細胞やその進化的な起源について新たな知見をあたえることができたと考えている.今後,ホヤ幼生の上衣細胞が真に神経幹細胞としての形質と機能を備えているかどうかについての検証を行うとともに,変態の過程において上衣細胞からニューロンを生みだす分子機構を解明することで神経幹細胞からのニューロンを生みだす脊索動物の全体に普遍的な機構について解明したいと考えている.
文 献
- Satoh, N.: Developmental Biology of Ascidians. Cambridge University Press, Cambridge (1994)
- Nicol, D. & Meinertzhagen, I. A.: Cell counts and maps in the larval central nervous system of the ascidian Ciona intestinalis (L.). J. Comp. Neurol., 309, 415-429 (1991)[PubMed]
- Meinertzhagen, I. A., Lemaire, P. & Okamura, Y.: The neurobiology of the ascidian tadpole larva: recent developments in an ancient chordate. Annu. Rev. Neurosci., 27, 453-485 (2004)[PubMed]
- Manni, L., Lane, N. J., Joly, J. S. et al.: Neurogenic and non-neurogenic placodes in ascidians. J. Exp. Zool. B Mol. Dev. Evol., 302, 483-504 (2004)[PubMed]
- Manni, L., Agnoletto, A., Zaniolo, G. et al.: Stomodeal and neurohypophysial placodes in Ciona intestinalis: insights into the origin of the pituitary gland. J. Exp. Zool. B Mol. Dev. Evol., 304, 324-339 (2005)[PubMed]
- Dufour, H. D., Chettouh, Z., Deyts C. et al.: Precraniate origin of cranial motoneurons. Proc. Natl. Acad. Sci. USA, 103, 8727-8732 (2006)[PubMed]
- Ando, R., Hama, H., Yamamoto-Hino, M. et al.: An optical marker based on the UV-induced green-to-red photoconversion of a fluorescent protein. Proc. Natl. Acad. Sci. USA, 99, 12651-12656 (2002)[PubMed]
- Horie, T., Nakagawa, M., Sasakura, Y. et al.: Cell type and function of neurons in the ascidian nervous system. Dev. Growth Differ., 51, 207-220 (2009)[PubMed]
- Coskun, V., Wu, H., Blanchi, B. et al.: CD133+ neural stem cells in the ependyma of mammalian postnatal forebrain. Proc. Natl. Acad. Sci. USA, 105, 1026-1031(2008)[PubMed]
- Chojnacki, A. K., Mak, G. K. & Weiss, S.: Identity crisis for adult periventricular neural stem cells: subventricular zone astrocytes, ependymal cells or both? Nat. Rev. Neurosci., 10, 153-163 (2009)[PubMed]
著者プロフィール
略歴:2007年 兵庫県立大学大学院生命理学研究科 修了,同年より筑波大学下田臨海実験センター 研究員.
研究テーマ:ホヤ幼生の神経回路網の機能とその分化機構の解明.
抱負:単純なホヤの中枢神経系の研究から,複雑な脊椎動物の中枢神経系を理解するための基本原理を明らかにしたいと考えています.
笹倉 靖徳(Sasakura Yasunori)
筑波大学下田臨海実験センター 准教授.
研究室URL:http://www.shimoda.tsukuba.ac.jp/~sasakura/index.html
© 2011 堀江健生・笹倉靖徳 Licensed under CC 表示 2.1 日本
