小胞体関連分解にかかわるレクチンOS-9の糖鎖認識の構造基盤
佐藤匡史・山口芳樹
(理化学研究所基幹研究所 糖鎖構造生物学研究チーム)
email:佐藤匡史,山口芳樹
DOI: 10.7875/first.author.2011.014
Structural basis for oligosaccharide recognition of misfolded glycoproteins by OS-9 in ER-associated degradation.
Tadashi Satoh, Yang Chen, Dan Hu, Shinya Hanashima, Kazuo Yamamoto, Yoshiki Yamaguchi
Molecular Cell, 40, 905-916 (2010)
OS-9は小胞体関連分解にかかわるレクチンであり,小胞体で特定の糖鎖構造を目印としてフォールディングに失敗した異常糖タンパク質と結合しユビキチン-プロテアソーム系へと導く.今回,筆者らは,世界にさきがけてOS-9の糖鎖認識ドメインと異常型の糖鎖との複合体の結晶構造を2.10Å分解能で解明することに成功した.解析の結果,異常型の糖鎖と結合するOS-9のWWモチーフを発見し,OS-9がこのWWモチーフを介して異常型の糖鎖と選択的に結合するしくみを明らかにした.この研究の成果は小胞体の品質管理機構を解明していくうえで重要な構造的な基盤をあたえるものとなった.
分泌系タンパク質は小胞体において分子シャペロンの助けを借りて高次構造の形成を行う.小胞体で正しい立体構造を獲得したタンパク質はゴルジ体以降の分泌経路に進み,さまざまなオルガネラや細胞表面に輸送また分泌される.異常なタンパク質が細胞内に蓄積あるいは分泌されると生体にとって有害となるため,分泌経路の入り口である小胞体には高次構造の形成に失敗したタンパク質を選別し再生もしくは分解するための緻密な“品質管理機構”が備わっている.ミスフォールディングしたタンパク質は最終的には小胞体から細胞質に逆行輸送され,そこでユビキチン-プロテアソーム系により分解をうける.この機構は小胞体関連分解(ERAD:endoplasmic reticulum-associated degradation)とよばれている.小胞体で合成されるタンパク質の多くはN-結合型糖鎖をもっており,糖鎖を“シグナル”としてさまざまな糖加水分解酵素,糖転移酵素,レクチンにより品質管理が行われている1)(図1).
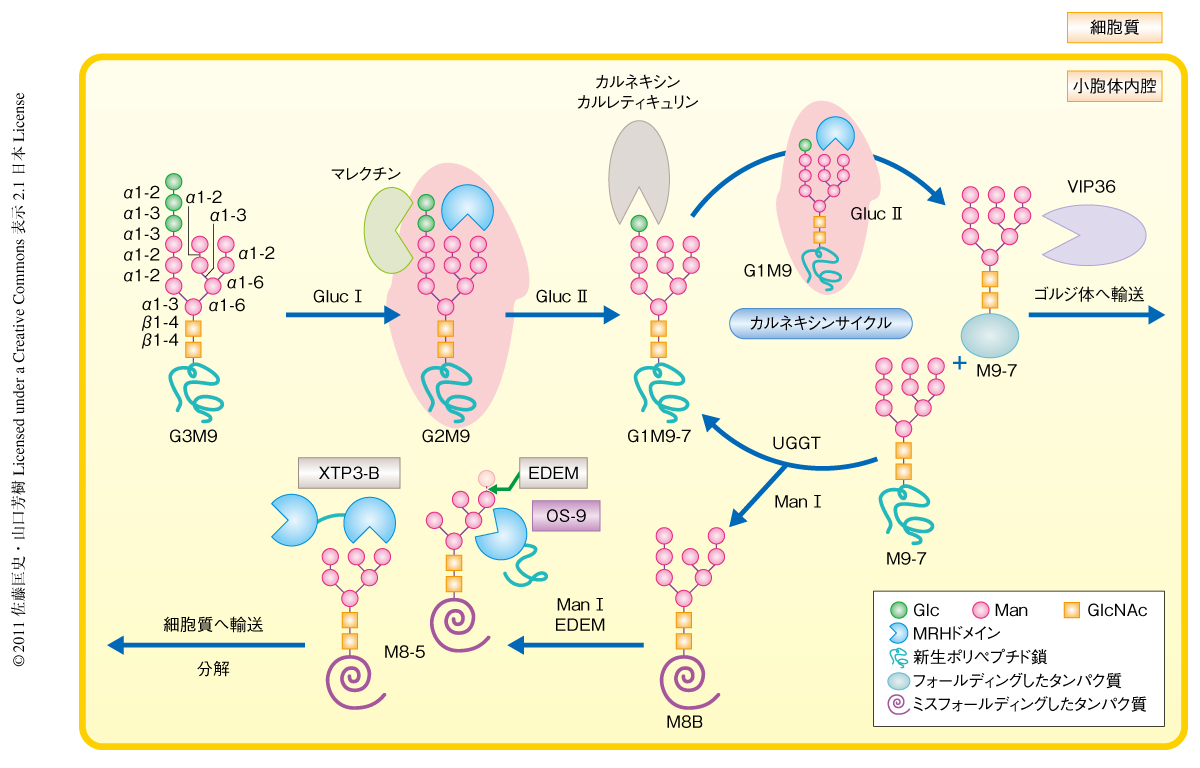
近年の研究により,小胞体関連分解はN-結合型糖タンパク質のマンノース残基の数(5~9個)により制御されていることが明らかになりつつある2).この制御にかかわっているのがマンノシダーゼ様タンパク質EDEMおよびレクチンOS-9である.出芽酵母ではそれぞれHtm1pおよびYos9pとよばれている.OS-9ファミリーはその重要性と新規性から大きな注目を集め国際的に熾烈な競争となり,1)小胞体関連分解における機能,2)ユビキチンリガーゼとの複合体の形成,3)EDEM/Htm1pの糖加水分解活性とOS-9/Yos9pの糖鎖結合の特異性との相関,を示したいくつかの論文がそれぞれほぼ同時期に報告された2).すなわち,OS-9およびYos9pはそれぞれHRD1-SEL1LユビキチンリガーゼおよびHrd1p-Hrd3pユビキチンリガーゼと小胞体膜で複合体を形成し,マンノース6-リン酸受容体の相同ドメインであるMRHドメインを介して小胞体関連分解の基質であるミスフォールディングした糖タンパク質と結合することで小胞体関連分解を促進すること,OS-9およびYos9pのMRHドメインはそれぞれEDEMおよびHtm1pのα1,2マンノシダーゼ活性により生成したマンノース1残基のトリミングされたMan-α1,6-Man結合を認識するという糖鎖特異性が明らかにされた.
筆者らは,小胞体で正しい立体構造を獲得したタンパク質をゴルジ体以降の分泌経路に運びMan8B-9を認識する輸送レクチンVIP36などの立体構造解析により,輸送レクチンによる糖鎖認識機構の構造的な基盤を明らかにしてきた3).一方,小胞体関連分解にかかわるMRHドメインをもつ細胞内レクチンの構造的な知見は得られておらず,“なぜマンノース残基のトリミングがOS-9の結合に必要なのか”,“どのようにOS-9はトリミングされた糖鎖を特異的に認識するのか”などのくわしい糖鎖認識機構は不明であった.このような研究背景のなか,筆者らは,ヒトOS-9のMRHドメインに注目し異常型の糖鎖との複合体の立体構造解析に挑んだ.
X線結晶構造解析にはmg量のタンパク質が必要であるが,ジスルフィド結合を多くもつタンパク質を大量に得ることは一般に困難であり構造生物学のボトルネックのひとつになっている.そこで筆者らは,ドメインの最適化,大腸菌で発現させる融合タンパク質の選択など,さまざまな発現系の設計とリフォールディング法の検討を重ね大量発現系を独自に構築した.この発現系を用いてジスルフィド結合を3個もつヒトOS-9のMRHドメインを1リットルの培養あたり4 mgも得ることができた(従来法と比較して約5倍量).OS-9のMRHドメインの結晶化は,α6マンノビオース,α6マンノトリオースなど種々のリガンドを用い,最終的にα3,α6マンノペンタオースを用いることにより良質な結晶を得ることに成功した.位相決定はセレノシステイン置換体結晶を用いた単波長異常分散法(single-wavelength anomalous dispersion:SAD)法により行い,2.85Åでの分解能のX線回折強度データを高エネルギー加速器研究機構フォトンファクトリーのAR NE3Aビームライン(波長は0.9790Å)で収集した.ネイティブ結晶のデータは台湾のNSRRC(National Synchrotron Radiation Research Center)の13B1ビームライン(波長は1.0000Å)で収集し,最終的に2.10Åでの分解能での構造解析に成功した(PDB ID:3AIH).
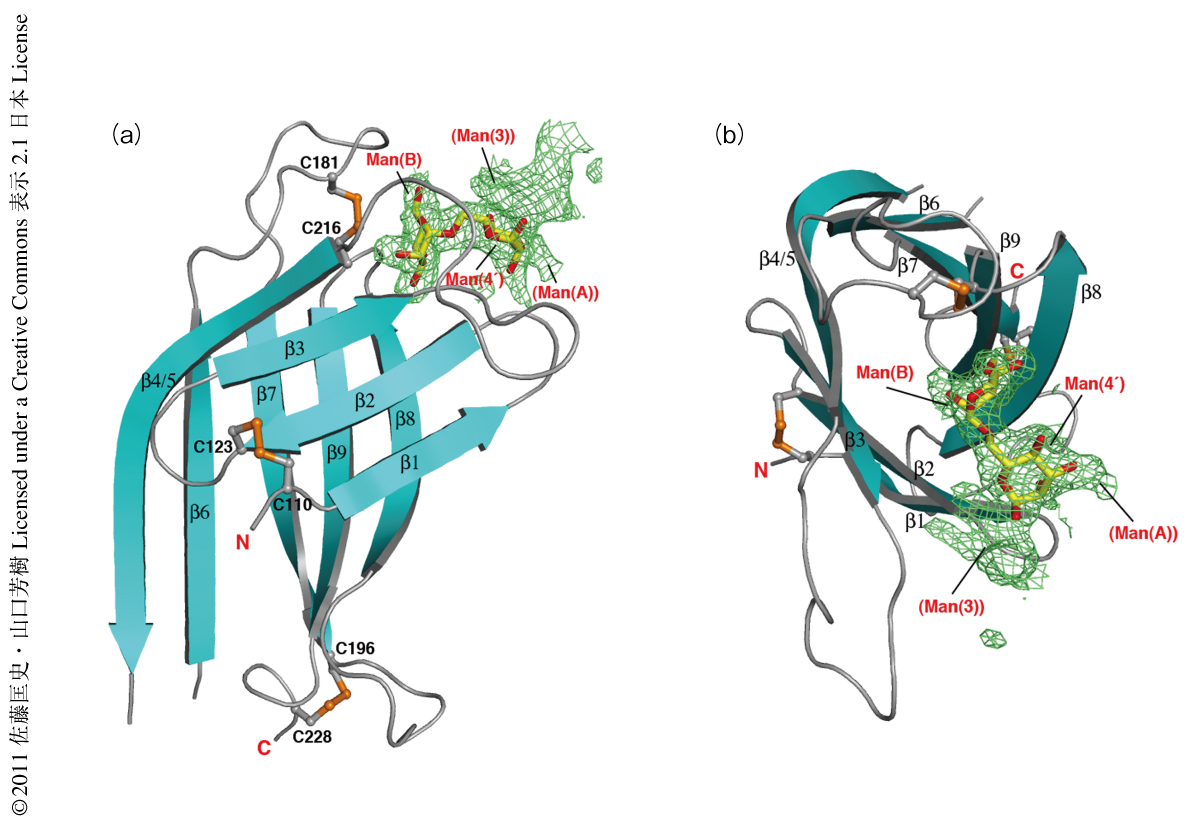
OS-9のMRHドメインは扁平でコンパクトなβバレル構造を形成しており,マンノース6-リン酸受容体と似た全体構造をとっていた4)(図2).このように基本的なフォールドは共通している一方で,OS-9のMRHドメインは特徴的なジスルフィド結合様式とβストランド構造をもっていた.すなわち,N末端から2番目と3番目のジスルフィド結合はマンノース6-リン酸受容体のものとほぼ一致していたが,1番目のものは大きく異なっていた.また,マンノース6-リン酸受容体ではβ4とβ5は別々に存在しているが,OS-9では長く連続したβ4/5によってマンノース6-リン酸受容体よりもさらにコンパクトなβバレル構造を形成していた.以上,OS-9のMRHドメインは特徴的なP型レクチンフォールドをもっているといえた.このP型レクチンとは,リン酸基を認識するマンノース6-リン酸受容体がもつドメインと相同なドメインMRHをもつレクチンファミリーの総称である.
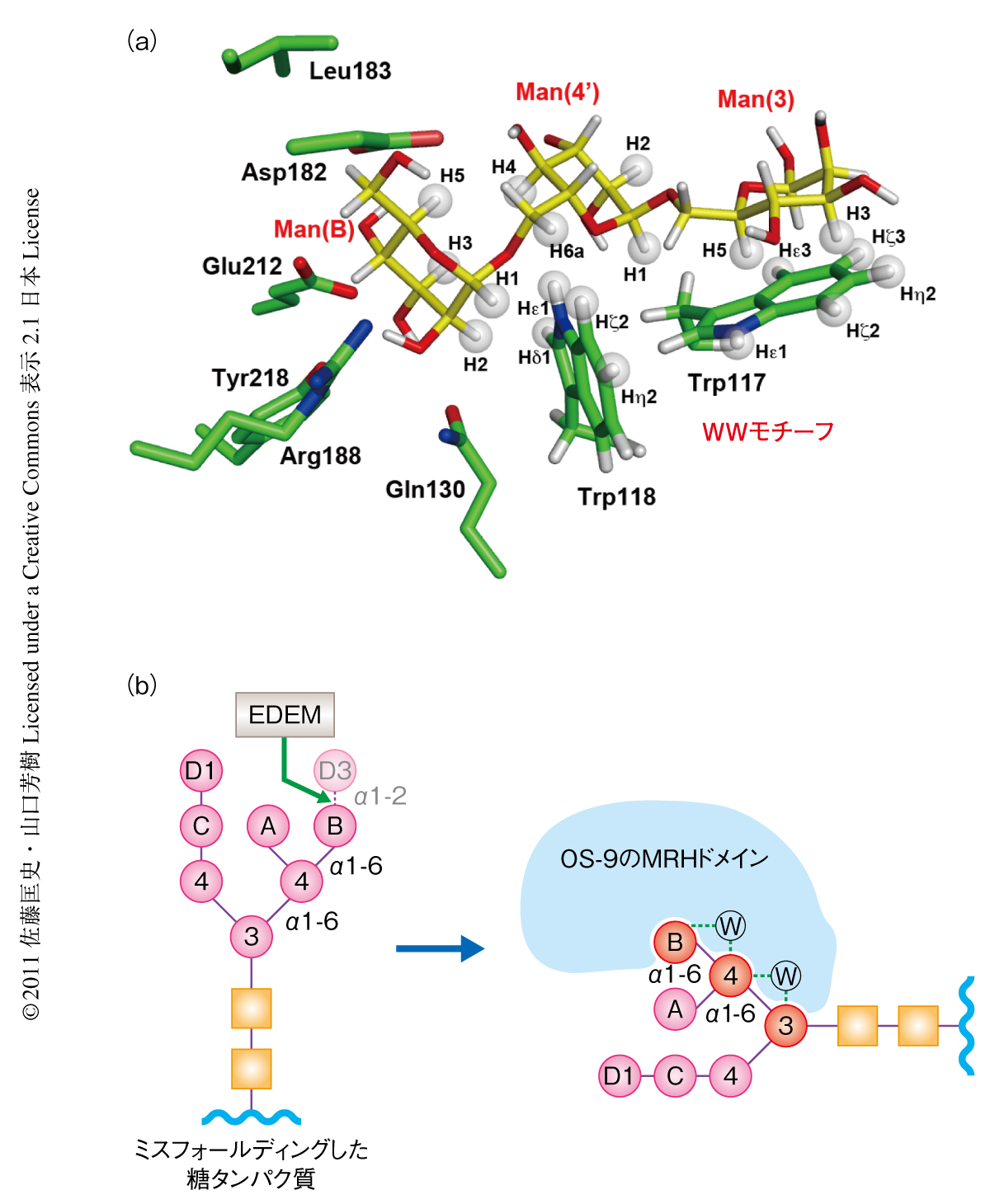
OS-9の糖鎖結合部位はマンノース6-リン酸受容体のマンノース6-リン酸との結合部位に相同なマンノース結合部位と,特徴的な連続する2つのトリプトファン残基から構成されたα6オリゴマンノース認識部位から構成されていた(図3a).この糖鎖認識部位を“WWモチーフ”と名づけた.また,NMRを用いてX線回折実験では明瞭に観測できなかったMan(3) 残基との相互作用を解析しその結合様式の全貌を明らかにした.すなわち,非還元末端のMan(B) はおもに水素結合により認識され,それにつづくMan(4’)-Man(3) はおもに疎水性相互作用により認識されていた.Trp117とMan(3) はタンパク質-糖鎖間の相互作用で頻繁にみられる疎水的なスタッキング5) を介して結合していた.一方,Man(4’) ではTrp118の側鎖がほぼ垂直に相互作用しており非常に珍しいものであった.非還元末端のMan(B) の2位の水酸基はOS-9に強く認識されていることから,EDEMによる高マンノース型糖鎖のα1,2結合したMan(D3) 残基のトリミングはOS-9のWWモチーフを介した相互作用に必須であることが明らかになった(図3b).このように,X線結晶構造解析とNMR解析を相補的に行うことでOS-9のMRHドメインがWWモチーフを介して高マンノース型糖鎖のManα1,6Manα1,6Man部分と特異的に結合することを示した.また,マンノース残基がトリミングされた高マンノース型糖鎖をOS-9が特異的に認識する構造基盤をはじめて明らかにした.
WWモチーフのそれぞれのトリプトファン残基の貢献度を調べるため部位特異的変異実験を行った.in vitroでの解析はレクチンテトラマー法にしたがい6),MRHドメインのC末端にビオチン化配列をコードしたcDNAを設計し大腸菌で発現させた組換えタンパク質を用いた.精製したタンパク質を酵素的にビオチン化しフィコエリスリン標識ストレプトアビジンと混合することにより蛍光標識した4量体プローブを作製し,細胞表面に発現する糖鎖に対してレクチンの結合をフローサイトメトリーにより定量化した.その結果,野生型OS-9はMan5GlcNAc2糖鎖をおもに発現したLec1細胞と強く結合したが,WWモチーフの1番目のトリプトファン残基をアラニン残基に置換した変異体では25倍ほど結合が減少し,2番目のトリプトファン残基をアラニン残基に置換した変異体では完全に結合能を失い,これらは構造解析の結果とよく合致していた.
小胞体関連分解の基質となる糖タンパク質とOS-9との相互作用解析は,HEK293T細胞を用い小胞体関連分解の基質のひとつである糖タンパク質ヒトα1アンチトリプシン変異体ヌルホンコンとFLAG標識したOS-9全長との免疫沈降法により行った.その結果,WWモチーフのトリプトファン残基それぞれの変異体は野生型と比較して顕著に結合が減少し,OS-9はMRHドメインのWWモチーフを介して小胞体関連分解の基質となる糖タンパク質と結合することが示された.その一方で,OS-9変異体とヒトα1アンチトリプシン変異体ヌルホンコンとの相互作用は完全には失われず,また,糖鎖付加しないヒトα1アンチトリプシン変異体ヌルホンコン変異体とOS-9との結合がみられることから,OS-9はミスフォールド糖タンパク質の“糖鎖”と“タンパク質”の両方を認識していることが示唆された.
OS-9のWWモチーフはカビからヒトまで,さらには,OS-9のホモログであり糖タンパク質の小胞体関連分解にかかわるXTP3-BのMRHドメインにおいても保存されている.出芽酵母Yos9pではWWモチーフの1番目のトリプトファン残基が同じ芳香族アミノ酸のフェニルアラニン残基に置換されているが,OS-9と同様に疎水的なスタッキングを介してα6マンノトリオース糖鎖と相互作用することが推測される.OS-9ファミリーにおいて高度に保存されたWWモチーフはOS-9のα6マンノトリオース構造を認識する基質結合の特異性を決定づけているといえる.一方,小胞体品質管理機構でカルネキシンサイクルにかかわるグルコシダーゼIIのβサブユニットや,リソソーム酵素の輸送などで機能するマンノース6-リン酸受容体のMRHドメインではWWモチーフはXxx-Tyrに置換されている.これら糖タンパク質の成熟経路および分泌経路にかかわるMRHドメインは,OS-9と異なりα1,2結合したオリゴマンノース(6-リン酸)構造を認識する結合特異性をもつことが知られている7-9).このように,この研究で明らかにしたMRHドメインの配列あるいは構造と機能との相関はMRHドメインをもつタンパク質の構造および機能解析を推進するうえで重要な知見であると考えられる.
この研究では,OS-9の異常型の糖鎖との複合体の結晶構造解析およびNMR解析から,OS-9による異常糖タンパク質の糖鎖認識機構をはじめて明らかにした.OS-9が結合する基質はミスフォールディングした糖タンパク質であり,OS-9による変性タンパク質の認識機構を解明することは今後の課題のひとつである.OS-9は小胞体シャペロンであるBiPやGRP94と複合体を形成することが知られており10),タンパク質部分の認識は間接的にBiPなどが担っている可能性もある.今後,これらのタンパク質の立体構造解析を進めることで,どのようにして異常なタンパク質をみわけて分解シグナルをOS-9に提示するのかなど,小胞体関連分解における分解シグナルの提示にまつわる疑問を立体構造的な観点から明らかにしていきたい.
略歴:2003年 日本大学大学院生物資源科学研究科博士課程 修了,同年 高エネルギー加速器研究機構物質構造科学研究所 博士研究員,2007年 米国National Cancer Institute博士研究員を経て,2008年より理化学研究所基幹研究所 研究員.
研究テーマ:ユビキチンおよび糖鎖修飾系の構造生物学.
抱負:立体構造解析から糖タンパク質の輸送機構および分解機構の新しい概念を打ち出したい.
山口 芳樹(Yoshiki Yamaguchi)
略歴:1998年 東京大学大学院薬学研究科博士課程 修了,2001年 同 助手,同年 名古屋市立大学大学院薬学研究科 講師を経て,2007年より理化学研究所基幹研究所 チームリーダー.
研究テーマ:糖タンパク質およびレクチンの構造生物学.
抱負:未解明な難問に挑み法則性を見い出したい.
研究室URL:http://www.riken.jp/r-world/research/lab/frontier-rs/sys-gly/stru/index.html
© 2011 佐藤匡史・山口芳樹 Licensed under CC 表示 2.1 日本
(理化学研究所基幹研究所 糖鎖構造生物学研究チーム)
email:佐藤匡史,山口芳樹
DOI: 10.7875/first.author.2011.014
Structural basis for oligosaccharide recognition of misfolded glycoproteins by OS-9 in ER-associated degradation.
Tadashi Satoh, Yang Chen, Dan Hu, Shinya Hanashima, Kazuo Yamamoto, Yoshiki Yamaguchi
Molecular Cell, 40, 905-916 (2010)
要 約
OS-9は小胞体関連分解にかかわるレクチンであり,小胞体で特定の糖鎖構造を目印としてフォールディングに失敗した異常糖タンパク質と結合しユビキチン-プロテアソーム系へと導く.今回,筆者らは,世界にさきがけてOS-9の糖鎖認識ドメインと異常型の糖鎖との複合体の結晶構造を2.10Å分解能で解明することに成功した.解析の結果,異常型の糖鎖と結合するOS-9のWWモチーフを発見し,OS-9がこのWWモチーフを介して異常型の糖鎖と選択的に結合するしくみを明らかにした.この研究の成果は小胞体の品質管理機構を解明していくうえで重要な構造的な基盤をあたえるものとなった.
はじめに
分泌系タンパク質は小胞体において分子シャペロンの助けを借りて高次構造の形成を行う.小胞体で正しい立体構造を獲得したタンパク質はゴルジ体以降の分泌経路に進み,さまざまなオルガネラや細胞表面に輸送また分泌される.異常なタンパク質が細胞内に蓄積あるいは分泌されると生体にとって有害となるため,分泌経路の入り口である小胞体には高次構造の形成に失敗したタンパク質を選別し再生もしくは分解するための緻密な“品質管理機構”が備わっている.ミスフォールディングしたタンパク質は最終的には小胞体から細胞質に逆行輸送され,そこでユビキチン-プロテアソーム系により分解をうける.この機構は小胞体関連分解(ERAD:endoplasmic reticulum-associated degradation)とよばれている.小胞体で合成されるタンパク質の多くはN-結合型糖鎖をもっており,糖鎖を“シグナル”としてさまざまな糖加水分解酵素,糖転移酵素,レクチンにより品質管理が行われている1)(図1).
近年の研究により,小胞体関連分解はN-結合型糖タンパク質のマンノース残基の数(5~9個)により制御されていることが明らかになりつつある2).この制御にかかわっているのがマンノシダーゼ様タンパク質EDEMおよびレクチンOS-9である.出芽酵母ではそれぞれHtm1pおよびYos9pとよばれている.OS-9ファミリーはその重要性と新規性から大きな注目を集め国際的に熾烈な競争となり,1)小胞体関連分解における機能,2)ユビキチンリガーゼとの複合体の形成,3)EDEM/Htm1pの糖加水分解活性とOS-9/Yos9pの糖鎖結合の特異性との相関,を示したいくつかの論文がそれぞれほぼ同時期に報告された2).すなわち,OS-9およびYos9pはそれぞれHRD1-SEL1LユビキチンリガーゼおよびHrd1p-Hrd3pユビキチンリガーゼと小胞体膜で複合体を形成し,マンノース6-リン酸受容体の相同ドメインであるMRHドメインを介して小胞体関連分解の基質であるミスフォールディングした糖タンパク質と結合することで小胞体関連分解を促進すること,OS-9およびYos9pのMRHドメインはそれぞれEDEMおよびHtm1pのα1,2マンノシダーゼ活性により生成したマンノース1残基のトリミングされたMan-α1,6-Man結合を認識するという糖鎖特異性が明らかにされた.
筆者らは,小胞体で正しい立体構造を獲得したタンパク質をゴルジ体以降の分泌経路に運びMan8B-9を認識する輸送レクチンVIP36などの立体構造解析により,輸送レクチンによる糖鎖認識機構の構造的な基盤を明らかにしてきた3).一方,小胞体関連分解にかかわるMRHドメインをもつ細胞内レクチンの構造的な知見は得られておらず,“なぜマンノース残基のトリミングがOS-9の結合に必要なのか”,“どのようにOS-9はトリミングされた糖鎖を特異的に認識するのか”などのくわしい糖鎖認識機構は不明であった.このような研究背景のなか,筆者らは,ヒトOS-9のMRHドメインに注目し異常型の糖鎖との複合体の立体構造解析に挑んだ.
1.OS-9のMRHドメインは特徴的なP型レクチンフォールドをもつ
X線結晶構造解析にはmg量のタンパク質が必要であるが,ジスルフィド結合を多くもつタンパク質を大量に得ることは一般に困難であり構造生物学のボトルネックのひとつになっている.そこで筆者らは,ドメインの最適化,大腸菌で発現させる融合タンパク質の選択など,さまざまな発現系の設計とリフォールディング法の検討を重ね大量発現系を独自に構築した.この発現系を用いてジスルフィド結合を3個もつヒトOS-9のMRHドメインを1リットルの培養あたり4 mgも得ることができた(従来法と比較して約5倍量).OS-9のMRHドメインの結晶化は,α6マンノビオース,α6マンノトリオースなど種々のリガンドを用い,最終的にα3,α6マンノペンタオースを用いることにより良質な結晶を得ることに成功した.位相決定はセレノシステイン置換体結晶を用いた単波長異常分散法(single-wavelength anomalous dispersion:SAD)法により行い,2.85Åでの分解能のX線回折強度データを高エネルギー加速器研究機構フォトンファクトリーのAR NE3Aビームライン(波長は0.9790Å)で収集した.ネイティブ結晶のデータは台湾のNSRRC(National Synchrotron Radiation Research Center)の13B1ビームライン(波長は1.0000Å)で収集し,最終的に2.10Åでの分解能での構造解析に成功した(PDB ID:3AIH).
OS-9のMRHドメインは扁平でコンパクトなβバレル構造を形成しており,マンノース6-リン酸受容体と似た全体構造をとっていた4)(図2).このように基本的なフォールドは共通している一方で,OS-9のMRHドメインは特徴的なジスルフィド結合様式とβストランド構造をもっていた.すなわち,N末端から2番目と3番目のジスルフィド結合はマンノース6-リン酸受容体のものとほぼ一致していたが,1番目のものは大きく異なっていた.また,マンノース6-リン酸受容体ではβ4とβ5は別々に存在しているが,OS-9では長く連続したβ4/5によってマンノース6-リン酸受容体よりもさらにコンパクトなβバレル構造を形成していた.以上,OS-9のMRHドメインは特徴的なP型レクチンフォールドをもっているといえた.このP型レクチンとは,リン酸基を認識するマンノース6-リン酸受容体がもつドメインと相同なドメインMRHをもつレクチンファミリーの総称である.
2.OS-9のMRHドメインはWWモチーフを介して高マンノース型糖鎖を認識する
OS-9の糖鎖結合部位はマンノース6-リン酸受容体のマンノース6-リン酸との結合部位に相同なマンノース結合部位と,特徴的な連続する2つのトリプトファン残基から構成されたα6オリゴマンノース認識部位から構成されていた(図3a).この糖鎖認識部位を“WWモチーフ”と名づけた.また,NMRを用いてX線回折実験では明瞭に観測できなかったMan(3) 残基との相互作用を解析しその結合様式の全貌を明らかにした.すなわち,非還元末端のMan(B) はおもに水素結合により認識され,それにつづくMan(4’)-Man(3) はおもに疎水性相互作用により認識されていた.Trp117とMan(3) はタンパク質-糖鎖間の相互作用で頻繁にみられる疎水的なスタッキング5) を介して結合していた.一方,Man(4’) ではTrp118の側鎖がほぼ垂直に相互作用しており非常に珍しいものであった.非還元末端のMan(B) の2位の水酸基はOS-9に強く認識されていることから,EDEMによる高マンノース型糖鎖のα1,2結合したMan(D3) 残基のトリミングはOS-9のWWモチーフを介した相互作用に必須であることが明らかになった(図3b).このように,X線結晶構造解析とNMR解析を相補的に行うことでOS-9のMRHドメインがWWモチーフを介して高マンノース型糖鎖のManα1,6Manα1,6Man部分と特異的に結合することを示した.また,マンノース残基がトリミングされた高マンノース型糖鎖をOS-9が特異的に認識する構造基盤をはじめて明らかにした.
3.OS-9はWWモチーフを介して小胞体関連分解の基質と結合する
WWモチーフのそれぞれのトリプトファン残基の貢献度を調べるため部位特異的変異実験を行った.in vitroでの解析はレクチンテトラマー法にしたがい6),MRHドメインのC末端にビオチン化配列をコードしたcDNAを設計し大腸菌で発現させた組換えタンパク質を用いた.精製したタンパク質を酵素的にビオチン化しフィコエリスリン標識ストレプトアビジンと混合することにより蛍光標識した4量体プローブを作製し,細胞表面に発現する糖鎖に対してレクチンの結合をフローサイトメトリーにより定量化した.その結果,野生型OS-9はMan5GlcNAc2糖鎖をおもに発現したLec1細胞と強く結合したが,WWモチーフの1番目のトリプトファン残基をアラニン残基に置換した変異体では25倍ほど結合が減少し,2番目のトリプトファン残基をアラニン残基に置換した変異体では完全に結合能を失い,これらは構造解析の結果とよく合致していた.
小胞体関連分解の基質となる糖タンパク質とOS-9との相互作用解析は,HEK293T細胞を用い小胞体関連分解の基質のひとつである糖タンパク質ヒトα1アンチトリプシン変異体ヌルホンコンとFLAG標識したOS-9全長との免疫沈降法により行った.その結果,WWモチーフのトリプトファン残基それぞれの変異体は野生型と比較して顕著に結合が減少し,OS-9はMRHドメインのWWモチーフを介して小胞体関連分解の基質となる糖タンパク質と結合することが示された.その一方で,OS-9変異体とヒトα1アンチトリプシン変異体ヌルホンコンとの相互作用は完全には失われず,また,糖鎖付加しないヒトα1アンチトリプシン変異体ヌルホンコン変異体とOS-9との結合がみられることから,OS-9はミスフォールド糖タンパク質の“糖鎖”と“タンパク質”の両方を認識していることが示唆された.
4.WWモチーフはMRHドメインの糖鎖結合の特異性を決定づける
OS-9のWWモチーフはカビからヒトまで,さらには,OS-9のホモログであり糖タンパク質の小胞体関連分解にかかわるXTP3-BのMRHドメインにおいても保存されている.出芽酵母Yos9pではWWモチーフの1番目のトリプトファン残基が同じ芳香族アミノ酸のフェニルアラニン残基に置換されているが,OS-9と同様に疎水的なスタッキングを介してα6マンノトリオース糖鎖と相互作用することが推測される.OS-9ファミリーにおいて高度に保存されたWWモチーフはOS-9のα6マンノトリオース構造を認識する基質結合の特異性を決定づけているといえる.一方,小胞体品質管理機構でカルネキシンサイクルにかかわるグルコシダーゼIIのβサブユニットや,リソソーム酵素の輸送などで機能するマンノース6-リン酸受容体のMRHドメインではWWモチーフはXxx-Tyrに置換されている.これら糖タンパク質の成熟経路および分泌経路にかかわるMRHドメインは,OS-9と異なりα1,2結合したオリゴマンノース(6-リン酸)構造を認識する結合特異性をもつことが知られている7-9).このように,この研究で明らかにしたMRHドメインの配列あるいは構造と機能との相関はMRHドメインをもつタンパク質の構造および機能解析を推進するうえで重要な知見であると考えられる.
おわりに
この研究では,OS-9の異常型の糖鎖との複合体の結晶構造解析およびNMR解析から,OS-9による異常糖タンパク質の糖鎖認識機構をはじめて明らかにした.OS-9が結合する基質はミスフォールディングした糖タンパク質であり,OS-9による変性タンパク質の認識機構を解明することは今後の課題のひとつである.OS-9は小胞体シャペロンであるBiPやGRP94と複合体を形成することが知られており10),タンパク質部分の認識は間接的にBiPなどが担っている可能性もある.今後,これらのタンパク質の立体構造解析を進めることで,どのようにして異常なタンパク質をみわけて分解シグナルをOS-9に提示するのかなど,小胞体関連分解における分解シグナルの提示にまつわる疑問を立体構造的な観点から明らかにしていきたい.
文 献
- Helenius, A., Aebi, M.: Roles of N-linked glycans in the endoplasmic reticulum. Annu. Rev. Biochem., 73, 1019-1049 (2004)[PubMed]
- Hosokawa, N., Kamiya, Y., Kato, K.: The role of MRH domain-containing lectins in ERAD. Glycobiology, 20, 651-660 (2010)[PubMed]
- Satoh, T., Cowieson, N. P., Hakamata, W. et al.: Structural basis for recognition of high mannose type glycoproteins by mammalian transport lectin VIP36. J. Biol. Chem., 282, 28246-28255 (2007)[PubMed]
- Kim, J. -J. P., Olson, L. J., Dahms, N. M.: Carbohydrate recognition by the mannose-6-phosphate receptors. Curr. Opin. Struct. Biol., 19, 534-542 (2009)[PubMed]
- Weis, W. I., Drickamer, K.: Structural basis of lectin-carbohydrate recognition. Annu. Rev. Biochem., 65, 441-473 (1996)[PubMed]
- Yamamoto, K., Kawasaki, N.: Detection of weak-binding sugar activity using membrane-based carbohydrates. Methods Enzymol., 478, 233-240 (2010)[PubMed]
- Song, X., Lasanajak, Y., Olson, L. J. et al.: Glycan microarray analysis of P-type lectins reveals distinct phosphomannose glycan recognition. J. Biol. Chem., 284, 35201-35214 (2009)[PubMed]
- Bohnsack, R. N., Song, X., Olson, L. J. et al.: Cation-independent mannose 6-phosphate receptor: a composite of distinct phosphomannosyl binding sites. J. Biol. Chem., 284, 35215-35226 (2009)[PubMed]
- Hu, D., Kamiya, Y., Totani, K. et al.: Sugar-binding activity of the MRH domain in ER α-glucosidase II βsubunit is important for efficient glucose trimming. Glycobiology, 19, 1127-1135 (2009)[PubMed]
- Christianson, J. C., Shaler, T. A., Tyler, R. E. et al.: OS-9 and GRP94 deliver mutant α1-antitrypsin to the Hrd1-SEL1L ubiquitin ligase complex for ERAD. Nat. Cell Biol., 10, 272-282 (2008)[PubMed]
著者プロフィール
略歴:2003年 日本大学大学院生物資源科学研究科博士課程 修了,同年 高エネルギー加速器研究機構物質構造科学研究所 博士研究員,2007年 米国National Cancer Institute博士研究員を経て,2008年より理化学研究所基幹研究所 研究員.
研究テーマ:ユビキチンおよび糖鎖修飾系の構造生物学.
抱負:立体構造解析から糖タンパク質の輸送機構および分解機構の新しい概念を打ち出したい.
山口 芳樹(Yoshiki Yamaguchi)
略歴:1998年 東京大学大学院薬学研究科博士課程 修了,2001年 同 助手,同年 名古屋市立大学大学院薬学研究科 講師を経て,2007年より理化学研究所基幹研究所 チームリーダー.
研究テーマ:糖タンパク質およびレクチンの構造生物学.
抱負:未解明な難問に挑み法則性を見い出したい.
研究室URL:http://www.riken.jp/r-world/research/lab/frontier-rs/sys-gly/stru/index.html
© 2011 佐藤匡史・山口芳樹 Licensed under CC 表示 2.1 日本
