コケ植物に特有の光化学系I超複合体
岩井 優和
(米国Lawrence Berkeley National Laboratory,Molecular Biophysics and Integrated Bioimaging Division)
email:岩井優和
DOI: 10.7875/first.author.2018.114
A unique supramolecular organization of photosystem I in the moss Physcomitrella patens.
Masakazu Iwai, Patricia Grob, Anthony T. Iavarone, Eva Nogales, Krishna K. Niyogi
Nature Plants, 4, 904-909 (2018)
植物の陸上への進出は地球史においてもっとも重要なイベントのひとつである.植物がどのようにその壮大な環境の変化に適応してきたかを知ることにより,将来に起こりうる環境の変化に対応する糸口をさぐることができる.光合成によるエネルギーの生産は植物の生存に不可欠であるが,これまでの研究により,水中に生息する藻類と陸上植物とのあいだで光合成の機能は細部にわたり修整されていることが明らかにされており,個々の環境の変化に適応してきたことが推察される.コケ植物は植物の進化の過程において藻類と陸上植物の中間に位置しており,陸上への進出における光環境への適応の機構を知るうえで重要と考えられるが,コケ植物における光合成の機能については不明な点が多い.この研究において,筆者らは,コケ植物ヒメツリガネゴケの光合成の機能に注目し,光合成にかかわるタンパク質の構造解析の結果,光化学系Iと集光タンパク質Iとの結合のしくみが藻類や陸上植物とは異なることを見い出した.また,コケ植物に特有の集光タンパク質IIであるLhcb9がその鍵となることが明らかにされた.
光合成の初期の反応過程は光エネルギーの吸収からはじまる.葉緑体のチラコイド膜に存在する光エネルギーを吸収する集光タンパク質はクロロフィルやカロテノイドなど複数の色素を保持する.それらの色素は吸収した光エネルギーを効率よく光合成反応にみちびくために緻密に配置されており,複数の集光タンパク質が協働することによりチラコイド膜における光エネルギーの伝達を制御する.
集光タンパク質により捕捉された光エネルギーは,光化学系タンパク質の複合体である光化学系Iおよび光化学系IIへと伝達される1).光エネルギーに起因する励起エネルギーにより光化学系タンパク質において電荷の分離反応がひき起こされ,チラコイド膜において電子伝達反応が駆動し化学エネルギーが生産される.しかし,光化学系タンパク質複合体へと伝達される光エネルギーが過剰になると活性酸素が発生し,葉緑体に悪影響をおよぼすだけでなくその影響は細胞の全体へと広がる.したがって,活性酸素の発生を抑制するため,チラコイド膜には過剰な光エネルギーを熱として発散する非光化学的な消光の機構が備わる.光エネルギーの伝達や非光化学的な消光の制御には集光タンパク質のもつ機能が大きく関与すると考えられているが,その詳細については不明な点が多い.
これまでの研究により,光化学系Iおよび光化学系IIにはそれぞれの構造に適した集光タンパク質が結合することがわかっている.一般的に,光化学系Iと結合する集光タンパク質を集光タンパク質I,光化学系IIと結合する集光タンパク質を集光タンパク質IIとよぶ.基本となる光合成における電子伝達反応は,光化学系IIが集光タンパク質IIから伝達された光エネルギーを消費し水分子から電子をうばうことによりひき起こされる.一方,光化学系Iは電子伝達反応により伝達されてきた電子および集光タンパク質Iから伝達された光エネルギーを消費し二酸化炭素の還元に要する化学エネルギーを生産する.そのときどきの葉緑体や細胞の状態に応じ,光化学系Iはその下流の電子の流れを適切に分配することも知られている2).この研究においては,光化学系Iに着目した.
光化学系Iの構造について,これまで,藻類および陸上植物を用いた研究がなされていた.どの生物種においても光化学系Iの基盤となるタンパク質の構造は似ているが,最近の研究により,集光タンパク質Iの結合の形態が生物種により異なることがわかってきた.集光タンパク質は光エネルギーの伝達や非光化学的な消光の制御を担うことから,異なる結合の形態は異なる自然環境に適した結果であることが示唆される.
この研究においては,コケ植物ヒメツリガネゴケ(Physcomitrella patens)の光化学系Iについて解析した.これまでの筆者らの研究により,ヒメツリガネゴケには光化学系Iと集光タンパク質Iとが結合したタンパク質超複合体として,大きさの異なる2つの光化学系I超複合体が存在することが明らかにされている3).密度勾配を利用した超遠心解析により,小型の光化学系I超複合体は陸上植物シロイヌナズナ(Arabidopsis thaliana)のもつ光化学系I超複合体と同じ大きさ,大型の光化学系I超複合体は緑藻クラミドモナス(Chlamydomonas reinhardtii)のもつ光化学系I超複合体とよく似た大きさであることがわかった.1種類の光化学系I超複合体のみをもつのが基本であるが,ヒメツリガネゴケは2種類の光化学系I超複合体をもつことから,それぞれの構造の解析を試みた.
Lhcb9はヒメツリガネゴケのゲノム解析によりはじめて見い出されたコケに特有の集光タンパク質IIである4).ヒメツリガネゴケを用いた研究の最大の利点は,遺伝子の相同組換え効率が高いため目的の遺伝子をねらった変異体の作製がほかの植物に比べ容易であることであり,遺伝子相同組換えによりLhcb9を欠損したヒメツリガネゴケ変異株を作製した3).超遠心解析の結果,Lhcb9欠損株には大型の光化学系I超複合体がなく,小型の光化学系I超複合体のみが存在することがわかった.このことから,分子系統学的に集光タンパク質IIに属するLhcb9が,大型の光化学系I超複合体に関与することが示唆された.
2種類の光化学系I超複合体の構造を明らかにするため,電子顕微鏡を用いた単粒子解析を行った.単粒子解析とは,高純度に精製した数万個の複合体を電子顕微鏡により個々に撮影し,画像解析プログラムにより高解像度の構造を再構築する手法である.この研究においては,シロイヌナズナ,クラミドモナス,ヒメツリガネゴケから精製した光化学系I超複合体を低解像度にて単粒子解析した(図1).その結果,シロイヌナズナの光化学系I超複合体は,これまでの報告のとおり,4つの集光タンパク質Iが光化学系Iの片側に連なって結合した特徴的な扇型をしており,この研究における単粒子解析法の有用性が確認された.クラミドモナスの光化学系I超複合体は少なくとも8つの集光タンパク質Iが光化学系Iの片側に結合したより大きな扇型をしていた.クラミドモナスの光化学系I超複合体について高解像度の構造はまだ報告されていないが,光化学系I複合体の逆側にも複数の集光タンパク質が結合することが確認された.ヒメツリガネゴケの小型の光化学系I超複合体の単粒子解析の結果,シロイヌナズナの光化学系I超複合体とよく似た構造であることが確認された一方,大型の光化学系I超複合体はクラミドモナスの光化学系I超複合体とはまったく異なる構造であった(図1).
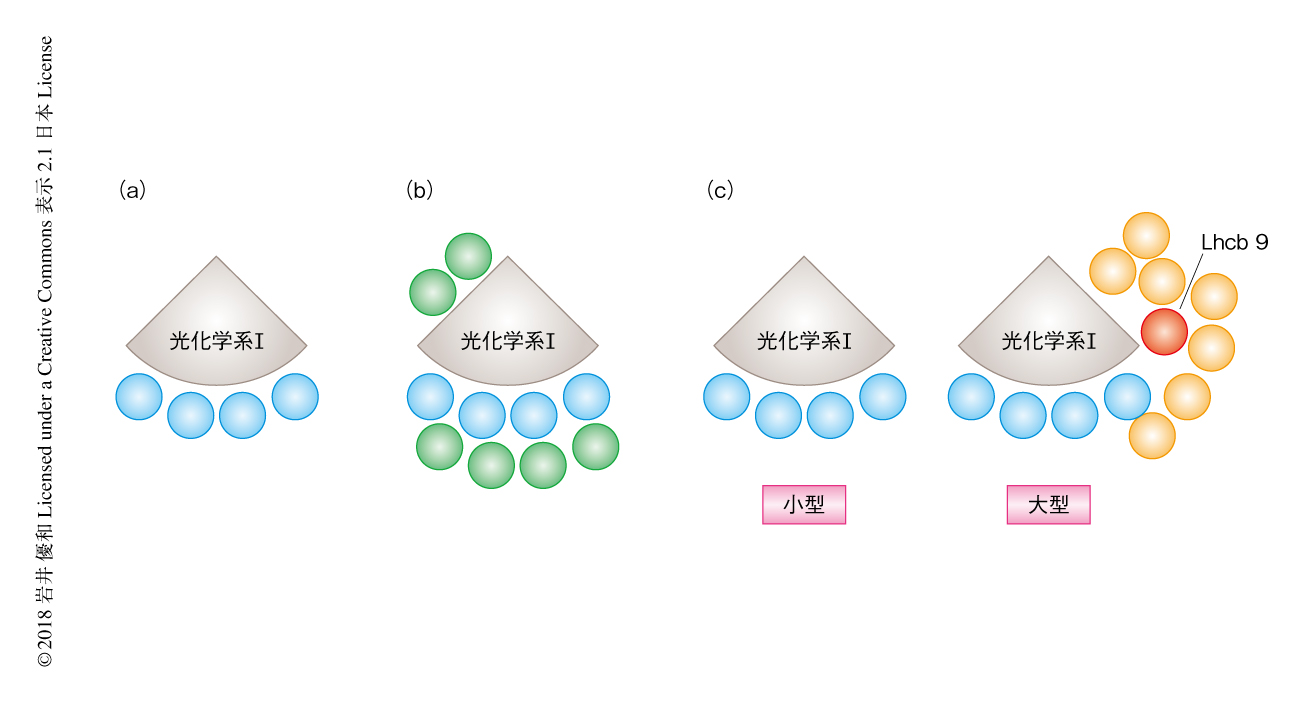
より鮮明な画像を得るため,クライオ電子顕微鏡を用いた単粒子解析を行った結果,ヒメツリガネゴケの光化学系I超複合体は光化学系Iの片側に大きく連なった集光タンパク質が結合した,これまでに例のない結合の形態をとることが明らかにされた.Lhcb9欠損株にくわえ,界面活性剤を利用した単粒子解析の結果,Lhcb9が直接に光化学系Iと結合し,さらに,光化学系Iには1つの三量体型の集光タンパク質および4つの単量体型の集光タンパク質が結合することが示唆された(図1).
これまでの研究により,光化学系Iおよび光化学系IIに伝達される光エネルギーの量の均衡を維持するため,光化学系IIと結合した集光タンパク質IIが光化学系Iとも直接に結合し光化学系Iの光エネルギーの量を増加させるステート遷移とよばれる機構が知られている.Lhcb9は分子系統学的に集光タンパク質IIに属するため,ステート遷移との関連が示唆された.ステート遷移は可逆的かつ短期的な光環境への適応の機構として観測されるが,Lhcb9欠損株を解析した結果,野生型と同等のステート遷移が観測されたことから,ヒメツリガネゴケにおけるLhcb9の結合による大型光化学系I超複合体の形成はステート遷移によるものではなく,より長期的な環境への適応の機構であると考えられた.
ヒメツリガネゴケの生活環において,細胞が原糸体とよばれる糸状の構造を形成する周期があり,この研究においては,おもに原糸体の葉緑体に存在する光化学系Iを解析した.原糸体は単純な構造の形態をしており生育環境の影響をうけやすい.原糸体にはクロロネマおよびカウロネマとよばれる2種類の細胞があり,クロロネマには葉緑体がより多く含まれ,カウロネマには葉緑体が少ないが成長が速い.これまでの研究により,グルコースを含む生育環境ではカウロネマの形成が促進されることがわかっている5,6).生育環境に含まれるグルコースの有無による光化学系I超複合体の形成の変化について調べた結果,グルコースを含まない環境においては大型の光化学系I超複合体は形成されないことがわかった.すなわち,カウロネマの形成が促進されると,大型の光化学系I超複合体の形成が促進されることが示唆された.カウロネマの形成が促進される環境においてはLhcb9の発現が上昇することも報告されている7).生育環境にグルコースが含まれることにより細胞質における解糖系によるエネルギーの生産が促進され,光合成によるエネルギーの生産との均衡を維持する必要が生じる.大型の光化学系I超複合体は小型の光化学系I超複合体と比べ光エネルギーの伝達の効率が40%ほど高いことがわかっている3).また,光化学系Iは状態に応じて電子の流れを制御することが知られている.その詳細はまだ不明だが,カウロネマの形成を促進する環境あるいは細胞の状態における,大型の光化学系I超複合体の形成の制御と電子伝達の制御との関連は興味深い.
この研究において明らかにされたコケ植物ヒメツリガネゴケに特有の光化学系I超複合体の形成は,植物の進化および陸上への進出において無数に起こったと考えられる適応のひとつにすぎないのかもしれない.また,陸上植物のゲノムにLhcb9をコードする遺伝子は残っていない.しかし,現存するLhcb9とそれに起因する特異な光化学系I超複合体の構造は,ヒメツリガネゴケの生育環境においてLhcb9が有利にはたらくことを示す.
植物の陸上への進出とその進化の研究には,コケ植物だけでなくそのほか多くの現生する植物の網羅的な解析が必要となる.植物の進化の機構は壮大な地球環境の変化に対する生存戦略の証跡であり,その解析はこれから起こりうる環境の変化への対応策を見い出すことにもなる.それに,いまや想像するほかすべはない地球史の一大イベントをこれらの研究をとおして垣間みることにより,植物が未踏の環境への進出を実現する鍵をも見い出せるかもしれない.
略歴:2009年 北海道大学大学院生命科学院 修了,同年 理化学研究所 基礎科学特別研究員,2012年 科学技術振興機構 さきがけ研究員,2015年 米国Lawrence Berkeley National Laboratoryポスドクを経て,2018年より同 研究員.
研究テーマ:光化学系タンパク質の適応のダイナミクスおよびタンパク質超複合体の形成の制御機構の生化学的な解析,生細胞イメージングによる解析,再構成による解析.
抱負:一日一善.
© 2018 岩井 優和 Licensed under CC 表示 2.1 日本
(米国Lawrence Berkeley National Laboratory,Molecular Biophysics and Integrated Bioimaging Division)
email:岩井優和
DOI: 10.7875/first.author.2018.114
A unique supramolecular organization of photosystem I in the moss Physcomitrella patens.
Masakazu Iwai, Patricia Grob, Anthony T. Iavarone, Eva Nogales, Krishna K. Niyogi
Nature Plants, 4, 904-909 (2018)
要 約
植物の陸上への進出は地球史においてもっとも重要なイベントのひとつである.植物がどのようにその壮大な環境の変化に適応してきたかを知ることにより,将来に起こりうる環境の変化に対応する糸口をさぐることができる.光合成によるエネルギーの生産は植物の生存に不可欠であるが,これまでの研究により,水中に生息する藻類と陸上植物とのあいだで光合成の機能は細部にわたり修整されていることが明らかにされており,個々の環境の変化に適応してきたことが推察される.コケ植物は植物の進化の過程において藻類と陸上植物の中間に位置しており,陸上への進出における光環境への適応の機構を知るうえで重要と考えられるが,コケ植物における光合成の機能については不明な点が多い.この研究において,筆者らは,コケ植物ヒメツリガネゴケの光合成の機能に注目し,光合成にかかわるタンパク質の構造解析の結果,光化学系Iと集光タンパク質Iとの結合のしくみが藻類や陸上植物とは異なることを見い出した.また,コケ植物に特有の集光タンパク質IIであるLhcb9がその鍵となることが明らかにされた.
はじめに
光合成の初期の反応過程は光エネルギーの吸収からはじまる.葉緑体のチラコイド膜に存在する光エネルギーを吸収する集光タンパク質はクロロフィルやカロテノイドなど複数の色素を保持する.それらの色素は吸収した光エネルギーを効率よく光合成反応にみちびくために緻密に配置されており,複数の集光タンパク質が協働することによりチラコイド膜における光エネルギーの伝達を制御する.
集光タンパク質により捕捉された光エネルギーは,光化学系タンパク質の複合体である光化学系Iおよび光化学系IIへと伝達される1).光エネルギーに起因する励起エネルギーにより光化学系タンパク質において電荷の分離反応がひき起こされ,チラコイド膜において電子伝達反応が駆動し化学エネルギーが生産される.しかし,光化学系タンパク質複合体へと伝達される光エネルギーが過剰になると活性酸素が発生し,葉緑体に悪影響をおよぼすだけでなくその影響は細胞の全体へと広がる.したがって,活性酸素の発生を抑制するため,チラコイド膜には過剰な光エネルギーを熱として発散する非光化学的な消光の機構が備わる.光エネルギーの伝達や非光化学的な消光の制御には集光タンパク質のもつ機能が大きく関与すると考えられているが,その詳細については不明な点が多い.
1.光化学系Iと集光タンパク質Iから構成されるタンパク質超複合体
これまでの研究により,光化学系Iおよび光化学系IIにはそれぞれの構造に適した集光タンパク質が結合することがわかっている.一般的に,光化学系Iと結合する集光タンパク質を集光タンパク質I,光化学系IIと結合する集光タンパク質を集光タンパク質IIとよぶ.基本となる光合成における電子伝達反応は,光化学系IIが集光タンパク質IIから伝達された光エネルギーを消費し水分子から電子をうばうことによりひき起こされる.一方,光化学系Iは電子伝達反応により伝達されてきた電子および集光タンパク質Iから伝達された光エネルギーを消費し二酸化炭素の還元に要する化学エネルギーを生産する.そのときどきの葉緑体や細胞の状態に応じ,光化学系Iはその下流の電子の流れを適切に分配することも知られている2).この研究においては,光化学系Iに着目した.
光化学系Iの構造について,これまで,藻類および陸上植物を用いた研究がなされていた.どの生物種においても光化学系Iの基盤となるタンパク質の構造は似ているが,最近の研究により,集光タンパク質Iの結合の形態が生物種により異なることがわかってきた.集光タンパク質は光エネルギーの伝達や非光化学的な消光の制御を担うことから,異なる結合の形態は異なる自然環境に適した結果であることが示唆される.
この研究においては,コケ植物ヒメツリガネゴケ(Physcomitrella patens)の光化学系Iについて解析した.これまでの筆者らの研究により,ヒメツリガネゴケには光化学系Iと集光タンパク質Iとが結合したタンパク質超複合体として,大きさの異なる2つの光化学系I超複合体が存在することが明らかにされている3).密度勾配を利用した超遠心解析により,小型の光化学系I超複合体は陸上植物シロイヌナズナ(Arabidopsis thaliana)のもつ光化学系I超複合体と同じ大きさ,大型の光化学系I超複合体は緑藻クラミドモナス(Chlamydomonas reinhardtii)のもつ光化学系I超複合体とよく似た大きさであることがわかった.1種類の光化学系I超複合体のみをもつのが基本であるが,ヒメツリガネゴケは2種類の光化学系I超複合体をもつことから,それぞれの構造の解析を試みた.
2.Lhcb9は大型の光化学系I超複合体の形成にかかわる
Lhcb9はヒメツリガネゴケのゲノム解析によりはじめて見い出されたコケに特有の集光タンパク質IIである4).ヒメツリガネゴケを用いた研究の最大の利点は,遺伝子の相同組換え効率が高いため目的の遺伝子をねらった変異体の作製がほかの植物に比べ容易であることであり,遺伝子相同組換えによりLhcb9を欠損したヒメツリガネゴケ変異株を作製した3).超遠心解析の結果,Lhcb9欠損株には大型の光化学系I超複合体がなく,小型の光化学系I超複合体のみが存在することがわかった.このことから,分子系統学的に集光タンパク質IIに属するLhcb9が,大型の光化学系I超複合体に関与することが示唆された.
2種類の光化学系I超複合体の構造を明らかにするため,電子顕微鏡を用いた単粒子解析を行った.単粒子解析とは,高純度に精製した数万個の複合体を電子顕微鏡により個々に撮影し,画像解析プログラムにより高解像度の構造を再構築する手法である.この研究においては,シロイヌナズナ,クラミドモナス,ヒメツリガネゴケから精製した光化学系I超複合体を低解像度にて単粒子解析した(図1).その結果,シロイヌナズナの光化学系I超複合体は,これまでの報告のとおり,4つの集光タンパク質Iが光化学系Iの片側に連なって結合した特徴的な扇型をしており,この研究における単粒子解析法の有用性が確認された.クラミドモナスの光化学系I超複合体は少なくとも8つの集光タンパク質Iが光化学系Iの片側に結合したより大きな扇型をしていた.クラミドモナスの光化学系I超複合体について高解像度の構造はまだ報告されていないが,光化学系I複合体の逆側にも複数の集光タンパク質が結合することが確認された.ヒメツリガネゴケの小型の光化学系I超複合体の単粒子解析の結果,シロイヌナズナの光化学系I超複合体とよく似た構造であることが確認された一方,大型の光化学系I超複合体はクラミドモナスの光化学系I超複合体とはまったく異なる構造であった(図1).
より鮮明な画像を得るため,クライオ電子顕微鏡を用いた単粒子解析を行った結果,ヒメツリガネゴケの光化学系I超複合体は光化学系Iの片側に大きく連なった集光タンパク質が結合した,これまでに例のない結合の形態をとることが明らかにされた.Lhcb9欠損株にくわえ,界面活性剤を利用した単粒子解析の結果,Lhcb9が直接に光化学系Iと結合し,さらに,光化学系Iには1つの三量体型の集光タンパク質および4つの単量体型の集光タンパク質が結合することが示唆された(図1).
3.生育環境に含まれる糖類と光化学系I超複合体の形成との関連
これまでの研究により,光化学系Iおよび光化学系IIに伝達される光エネルギーの量の均衡を維持するため,光化学系IIと結合した集光タンパク質IIが光化学系Iとも直接に結合し光化学系Iの光エネルギーの量を増加させるステート遷移とよばれる機構が知られている.Lhcb9は分子系統学的に集光タンパク質IIに属するため,ステート遷移との関連が示唆された.ステート遷移は可逆的かつ短期的な光環境への適応の機構として観測されるが,Lhcb9欠損株を解析した結果,野生型と同等のステート遷移が観測されたことから,ヒメツリガネゴケにおけるLhcb9の結合による大型光化学系I超複合体の形成はステート遷移によるものではなく,より長期的な環境への適応の機構であると考えられた.
ヒメツリガネゴケの生活環において,細胞が原糸体とよばれる糸状の構造を形成する周期があり,この研究においては,おもに原糸体の葉緑体に存在する光化学系Iを解析した.原糸体は単純な構造の形態をしており生育環境の影響をうけやすい.原糸体にはクロロネマおよびカウロネマとよばれる2種類の細胞があり,クロロネマには葉緑体がより多く含まれ,カウロネマには葉緑体が少ないが成長が速い.これまでの研究により,グルコースを含む生育環境ではカウロネマの形成が促進されることがわかっている5,6).生育環境に含まれるグルコースの有無による光化学系I超複合体の形成の変化について調べた結果,グルコースを含まない環境においては大型の光化学系I超複合体は形成されないことがわかった.すなわち,カウロネマの形成が促進されると,大型の光化学系I超複合体の形成が促進されることが示唆された.カウロネマの形成が促進される環境においてはLhcb9の発現が上昇することも報告されている7).生育環境にグルコースが含まれることにより細胞質における解糖系によるエネルギーの生産が促進され,光合成によるエネルギーの生産との均衡を維持する必要が生じる.大型の光化学系I超複合体は小型の光化学系I超複合体と比べ光エネルギーの伝達の効率が40%ほど高いことがわかっている3).また,光化学系Iは状態に応じて電子の流れを制御することが知られている.その詳細はまだ不明だが,カウロネマの形成を促進する環境あるいは細胞の状態における,大型の光化学系I超複合体の形成の制御と電子伝達の制御との関連は興味深い.
おわりに
この研究において明らかにされたコケ植物ヒメツリガネゴケに特有の光化学系I超複合体の形成は,植物の進化および陸上への進出において無数に起こったと考えられる適応のひとつにすぎないのかもしれない.また,陸上植物のゲノムにLhcb9をコードする遺伝子は残っていない.しかし,現存するLhcb9とそれに起因する特異な光化学系I超複合体の構造は,ヒメツリガネゴケの生育環境においてLhcb9が有利にはたらくことを示す.
植物の陸上への進出とその進化の研究には,コケ植物だけでなくそのほか多くの現生する植物の網羅的な解析が必要となる.植物の進化の機構は壮大な地球環境の変化に対する生存戦略の証跡であり,その解析はこれから起こりうる環境の変化への対応策を見い出すことにもなる.それに,いまや想像するほかすべはない地球史の一大イベントをこれらの研究をとおして垣間みることにより,植物が未踏の環境への進出を実現する鍵をも見い出せるかもしれない.
文 献
- Nelson, N. & Yocum, C. F.: Structure and function of photosystems I and II. Annu. Rev. Plant Biol., 57, 521-565 (2006)[PubMed]
- Shikanai, T. & Yamamoto, H.: Contribution of cyclic and pseudo-cyclic electron transport to the formation of proton motive force in chloroplasts. Mol. Plant, 10, 20-29 (2017)[PubMed]
- Iwai, M., Yokono, M., Kono, M. et al.: Light-harvesting complex Lhcb9 confers a green alga-type photosystem I supercomplex to the moss Physcomitrella patens. Nat. Plants, 1, 14008 (2015)[PubMed]
- Rensing, S. A., Lang, D., Zimmer, A. D. et al.: The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science, 319, 64-69 (2008)[PubMed]
- Olsson, T., Thelander, M. & Ronne, H.: A novel type of chloroplast stromal hexokinase is the major glucose-phosphorylating enzyme in the moss Physcomitrella patens. J. Biol. Chem., 278, 44439-44447 (2003)[PubMed]
- Thelander, M., Olsson, T. & Ronne, H.: Effect of the energy supply on filamentous growth and development in Physcomitrella patens. J. Exp. Bot., 56, 653-662 (2005)[PubMed]
- Sugiyama, T., Ishida, T., Tabei, N. et al.: Involvement of PpDof1 transcriptional repressor in the nutrient condition-dependent growth control of protonemal filaments in Physcomitrella patens. J. Exp. Bot., 63, 3185-3197 (2012)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2009年 北海道大学大学院生命科学院 修了,同年 理化学研究所 基礎科学特別研究員,2012年 科学技術振興機構 さきがけ研究員,2015年 米国Lawrence Berkeley National Laboratoryポスドクを経て,2018年より同 研究員.
研究テーマ:光化学系タンパク質の適応のダイナミクスおよびタンパク質超複合体の形成の制御機構の生化学的な解析,生細胞イメージングによる解析,再構成による解析.
抱負:一日一善.
© 2018 岩井 優和 Licensed under CC 表示 2.1 日本
