運動学習をへて現われる視床から運動野へのシグナルは入力層に依存して異なるダイナミクスを示す
田中康代・田中康裕・松崎政紀
(東京大学大学院医学系研究科 細胞分子生理学分野)
email:田中康代,田中康裕,松崎政紀
DOI: 10.7875/first.author.2018.093
Thalamocortical axonal activity in motor cortex exhibits layer-specific dynamics during motor learning.
Yasuyo H. Tanaka, Yasuhiro R. Tanaka, Masashi Kondo, Shin-Ichiro Terada, Yasuo Kawaguchi, Masanori Matsuzaki
Neuron, 100, 244-258.e12 (2018)
大脳基底核および小脳は運動学習において非常に重要な役割をはたすが,それらが運動学習の際にどのようなシグナルを大脳皮質に送るのかはわかっていない.大脳基底核あるいは小脳からのシグナルは視床をとおり大脳皮質へと送られる.この研究において,筆者らは,マウスが自発的にレバー引き課題を学習する際に,視床から1次運動野へと投射する軸索のブトンにおける神経活動を2光子顕微鏡によるCa2+イメージング法により測定した.その結果,視床から運動野へのシグナルは入力層により異なることが明らかにされた.第1層に入力するブトンにおける神経活動はレバー引きの開始時および終了時に強くなり,第3層に入力するブトンにおける神経活動はレバー引きの開始時にのみ強くなった.また,レバーを引くときのブトンにおける神経活動はシークエンス構造を示し,その持続する時間は第1層に入力するブトンにおいては長く,第3層に入力するブトンにおいては短かった.さらに,第1層に入力するブトンにおける神経活動はレバーを引く速度,レバーを引く時間の長さ,レバーの軌道の安定性と関係し,第3層に入力するブトンにおける神経活動はレバーを引く成功率と関係した.また,第1層に入力するブトンにおける神経活動および第3層に入力するブトンにおける神経活動は大脳基底核あるいは小脳に強い影響をうけ,一方,それらの神経活動のダイナミクスには大脳基底核および小脳の神経活動が必要であった.
ピアノや手術といった高度な熟練の必要なものから,文字の入力や食器洗いなどの日常の動作にいたるまで,何度も何度もくり返して訓練することで目的の達成のための効率のよい運動スキルが習得されることを,われわれは日常的に経験している.これまで,1次運動野のニューロンは学習によりダイナミックな神経活動の変化を示すことが明らかにされているが1,2),運動学習中には脳のさまざまな領域において大きな変化が起こるため,運動学習の機構を解明するためには脳の領域のあいだの情報のやりとりを調べることが非常に重要である.大脳基底核は脳の深部に存在する複数の神経核からなり,運動の制御,認知機能,動機づけ,報酬などさまざまな機能を担い,また,小脳は知覚と運動機能を統合し細かい運動の制御にかかわる3).大脳基底核および小脳の障害は運動失調および運動障害疾患をもたらす.大脳基底核あるいは小脳からのシグナルは視床を介して運動野などの大脳皮質に送られ4,5),最終的に1次運動野の皮質脊髄投射ニューロンが脊髄へと出力を送り運動を遂行する.この運動系の経路において,視床は非常に重要なハブととらえられる.筆者らは,視床から運動野へと送られるシグナルの時間的なパターンを調べることは運動学習の機構の解明において大きな足がかりになると考え,視床から1次運動野へとシグナルを送る経路に着目した.
視床には核とよばれる細胞体の集団が多く存在するが,運動野へと投射する視床核のうち,前腹側核や内側腹側核から運動野への投射は第1層に多く分布し,外側腹側核からの投射は第3層や第5層に分布する6,7).また,前腹側核や内側腹側核は大脳基底核からの投射を多くうけ,外側腹側核は小脳からの投射を多くうける8).投射パターンの異なる視床皮質軸索の神経活動は,それぞれ,運動学習中にどのような活動パターンを示すのであろうか.この研究において,筆者らは,視床皮質軸索からの投射パターンに解剖学的な違いのあることを確認したうえで,マウスにレバーを自発的に一定の時間だけ引くと報酬として水がもらえるという運動課題を実施させ,どのようなシグナルが視床から運動野へと送られるかを調べた.
光遺伝学的な手法を用いて1次運動野へ投射する視床皮質軸索の神経活動を活性化させた.視床皮質軸索にアデノ随伴ウイルスを用いて光駆動性の非選択的なカチオンチャネルであるチャネルロドプシン2を発現させ,頭上から1次運動野に光を照射し視床皮質軸索における神経活動を活性化したところ,それにともない手の運動が誘発された.また,光遺伝学的な手法を用いて視床の神経活動を抑制した.運動野に投射する視床核のニューロンにアデノ随伴ウイルスを用いて光駆動性のプロトンポンプであるアーキロドプシンTを発現させ,レバー引き課題を学習させたのち,視床へ光を照射し視床核のニューロンにおける神経活動を抑制したところ,レバーを引く成功率が有意に低下した.このことから,視床から運動野へと投射する経路に手の動きそのものやレバーを一定の時間だけ引くことに関連する経路が存在することがわかった.
レバー引き課題を実行しているときの視床皮質軸索のブトンにおける神経活動を運動野において観察するため,2光子顕微鏡を用いたCa2+イメージング法を実施した.従来の電気生理学的な手法ではブトンだけのシグナルを分離することは不可能であったが,アデノ随伴ウイルスを用いて運動野に投射する視床皮質軸索にCa2+応答タンパク質であるGCaMP6f 9) を発現させ,ブトンにおけるCa2+濃度の上昇を蛍光輝度の変化により検出した.レバーを引くと脳はゆれるため,わずか数μmのブトンにおける神経活動をとらえることは困難であったが,運動野に赤色蛍光タンパク質を同時に発現させ,さらに,深さ方向に4μmごとの3つの平面でイメージング画像を取得し,赤色の蛍光輝度をもとにオフラインで3次元のゆれを補正することにより,ゆれによるアーティファクトを最小限に抑えてイメージング画像を解析することに成功した.このようにして,視床皮質軸索のブトンにおける神経活動を運動野の第1層および第3層において測定し,レバー引き課題を実行しているときの神経活動を学習の前後で比較した.
レバーを引くときの第1層および第3層の視床軸索のブトンにおける神経活動からレバーを引く軌道を予測しその精度を比較すると,学習をへてどちらも予測の精度が高くなった.また,セッション内の神経活動の再現性も学習をへて高くなった.このことは,学習により視床皮質軸索の神経活動がレバーを引く情報をもつようになったことを意味した.さらに,レバーを引くときの神経活動のタイミングについて解析した.その結果,学習の後期には第1層の視床皮質軸索においてはレバー引きの開始時と終了時に顕著な神経活動を示すブトンが多くなり,第3層においてはレバー引きの開始時に顕著な神経活動を示すブトンが多くなった.さらに,レバーを引くときのブトンの集団の神経活動のシークエンス構造に着目した.視野から2つのブトンをとりだしたとき,レバーを引くときに平均の神経活動が先行していたブトンにおける神経活動からもう一方のブトンにおける神経活動を予測できるかを調べ,有意な予測の精度をもつブトンのペアを定めた.レバーを引く期間のなかで,このようなブトンのペアがランダムよりも多く検出される連続した期間をシークエンス長とした.その結果,第1層のブトンは第3層のブトンよりも長いシークエンス長をもつ神経活動の構造を示すことがわかった.これらの結果より,レバー引き課題の学習に関係する第1層および第3層のブトンは,レバーを引くときに異なるダイナミクスを示すことが明らかにされた.
ここまでに示された第1層および第3層のブトンにおける神経活動のダイナミクスは個体の行動において何を表象しているのであろうか.この問題に答えるため,ブトンにおける神経活動とレバーを引く行動との関係について調べた.ブトンにおける神経活動のダイナミクスを特徴づける量として,レバー引き開始時および終了時の神経活動,および,シークエンス長を用い,レバーを引く成功率,レバーを引く速度およびレバーを戻す速度,レバーを引く時間の長さ,おのおののセッションにおけるレバーの軌道の再現性といった行動を特徴づける量が統計的に説明されるかどうか検討した.行動は学習の初期から後期にかけて変化する可能性があるため,この効果についてはすべてのモデルに組み込んだうえでおのおのの量の効果を判定した.統計モデルとしては一般化線形モデルを用い,最適なモデルは赤池情報量基準を用いて選んだ.その結果,第1層のブトンにおける神経活動はレバーを引く速度,レバーを引く時間,レバーの軌道の再現性,つまり,レバーを引くキネマティクスに関係し,第3層のブトンにおける神経活動はレバーを引く成功率に関係することが明らかにされた.
第1層および第3層の視床皮質軸索のブトンにおける神経活動のダイナミクスが生じる原因は何であろうか.視床は大脳基底核および小脳からの入力を強くうけることがわかっていたことから,大脳基底核の出力を担う黒質網様部あるいは小脳核のニューロンを光遺伝学的な手法により活性化したときの視床皮質軸索のブトンの応答性をみた.その結果,黒質網様部を活性化したときは第1層のブトンが第3層のブトンよりも高率に応答し,小脳核を活性化したときは第3層のブトンが第1層のブトンよりも高率に反応した.これは,解剖学的な結果とも矛盾しなかった.さらに,線条体あるいは小脳核を破壊したマウスにレバー引き課題を実施させ視床皮質軸索のブトンにおける神経活動を測定したところ,いずれもレバー引き課題は正常に学習されず,ブトンにおける神経活動のダイナミクスについても第1層および第3層においてきわだった特徴は現われなかった.このことから,レバー引き課題の学習をとおして視床皮質軸索のブトンにおいて神経活動のダイナミクスが現われるには,大脳基底核および小脳の両方の神経活動が必要とされることが明らかにされた.
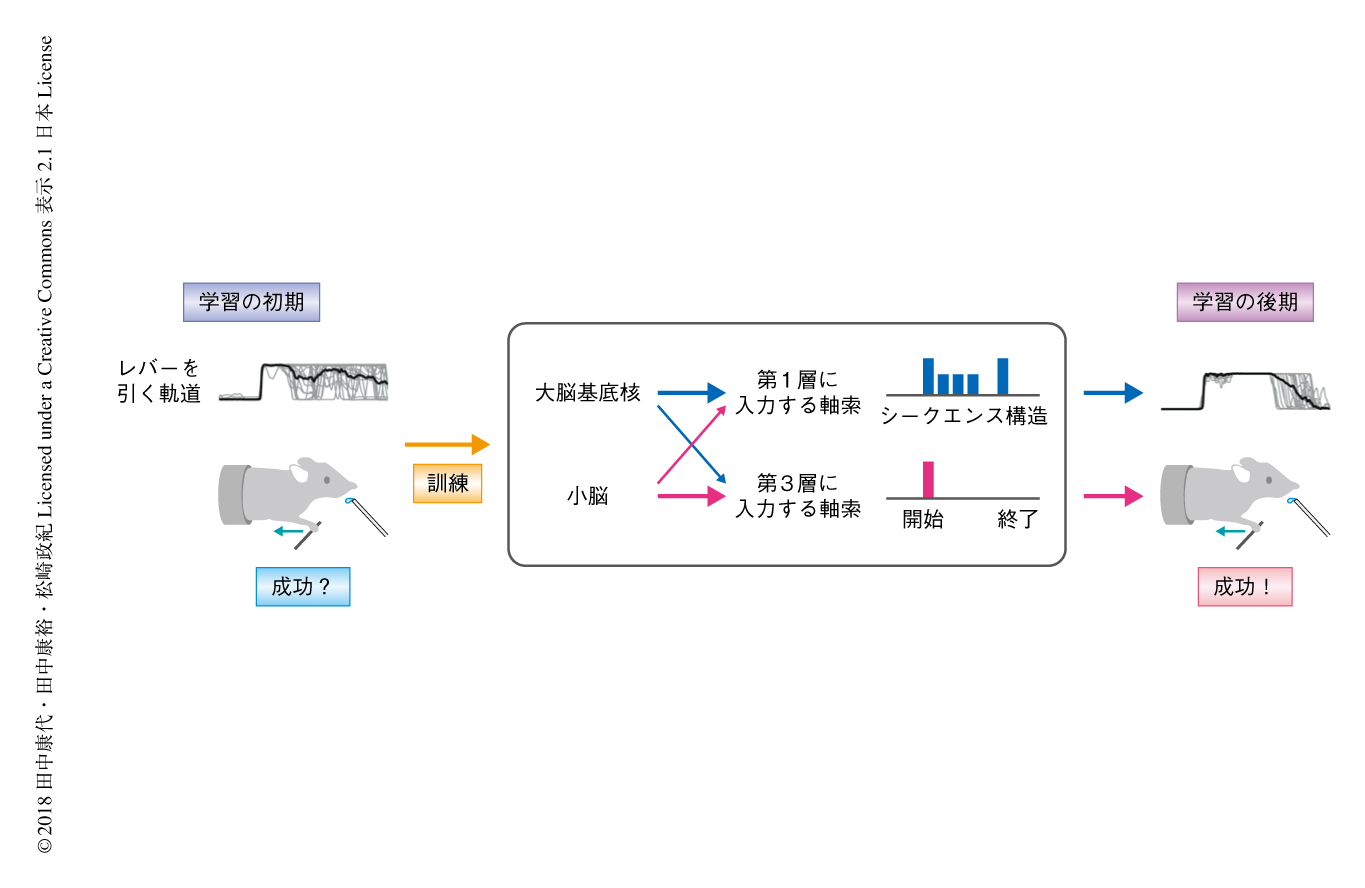
この研究において,筆者らは,これまでの解剖学的な視床皮質からの投射経路の知見から,運動学習における層に依存的な機能の違いを予測し,その実証を試みた.その結果,運動学習をへて視床から運動野へは少なくとも2種類のシグナルが層に依存的に入力されることが明らかにされた.それらは大脳基底核あるいは小脳からのシグナルの影響を強くうけるものであり,行動と神経活動との関係も明らかにされた(図1).
今回の研究を発展させるためには,以下の問題に取り組む必要があるだろう.レバー引き課題を実行しているときに,視床皮質の投射経路に選択的に神経活動を抑制すると個体の行動はどのように変化するのか.第1層のブトンにおける神経活動はレバーを引くキネマティクス,第3層のブトンにおける神経活動はレバーを引く成功率と関係するという結果であったが,その因果関係は得られるかどうか.また,大脳基底核あるいは小脳のどちらかが破壊されると両方のシグナルが消失したことから,レバー引き課題の学習中には互いに影響をうけると考えられるが,どのようにしてこれらのシグナルは運動野にて統合されるのか,という点である.
運動学習中に大脳基底核あるいは小脳からそれぞれ視床を介して入力するシグナルが運動野において統合され,その結果が脊髄へと出力されると同時に,ふたたび大脳基底核および小脳へと送られる.そして,またそれぞれのシグナルが視床をとおり運動野へと送られる.このようにして形成されるループ構造はくり返しの訓練により外界から修飾をうけ,さらに,脳における相互作用をとおししだいにきわだった特徴をもつシグナルを形成していくと考えられる.そして,それらのシグナルが最終的にそれぞれの役割を担い運動スキルは習得されると考えられる.大脳基底核系と小脳系との相互作用は神経科学における最近のトピックでもあり,近年,ますますその重要性が注目されている10).これらのことをひとつひとつ明らかにすることは,運動の機構の解明や病態の理解にも貢献すると考えらえる.
略歴:2012年 京都大学大学院医学研究科にて博士号取得,同年 基礎生物学研究所 博士研究員,2016年 東京大学大学院医学系研究科 博士研究員を経て,2017年より同 特任助教.
研究テーマ:脳のなかの基盤とそれにより実現される機能との関係.
関心事:主観的な時間の長さ.
田中 康裕(Yasuhiro R. Tanaka)
略歴:2012年 京都大学大学院医学研究科にて博士号取得,同年 基礎生物学研究所 博士研究員を経て,2016年より東京大学大学院医学系研究科 助教.
松崎政紀(Masanori Matsuzaki)
東京大学大学院医学系研究科 教授.
研究室URL:http://plaza.umin.ac.jp/~Matsuzaki-Lab/index.html
© 2018 田中康代・田中康裕・松崎政紀 Licensed under CC 表示 2.1 日本
(東京大学大学院医学系研究科 細胞分子生理学分野)
email:田中康代,田中康裕,松崎政紀
DOI: 10.7875/first.author.2018.093
Thalamocortical axonal activity in motor cortex exhibits layer-specific dynamics during motor learning.
Yasuyo H. Tanaka, Yasuhiro R. Tanaka, Masashi Kondo, Shin-Ichiro Terada, Yasuo Kawaguchi, Masanori Matsuzaki
Neuron, 100, 244-258.e12 (2018)
要 約
大脳基底核および小脳は運動学習において非常に重要な役割をはたすが,それらが運動学習の際にどのようなシグナルを大脳皮質に送るのかはわかっていない.大脳基底核あるいは小脳からのシグナルは視床をとおり大脳皮質へと送られる.この研究において,筆者らは,マウスが自発的にレバー引き課題を学習する際に,視床から1次運動野へと投射する軸索のブトンにおける神経活動を2光子顕微鏡によるCa2+イメージング法により測定した.その結果,視床から運動野へのシグナルは入力層により異なることが明らかにされた.第1層に入力するブトンにおける神経活動はレバー引きの開始時および終了時に強くなり,第3層に入力するブトンにおける神経活動はレバー引きの開始時にのみ強くなった.また,レバーを引くときのブトンにおける神経活動はシークエンス構造を示し,その持続する時間は第1層に入力するブトンにおいては長く,第3層に入力するブトンにおいては短かった.さらに,第1層に入力するブトンにおける神経活動はレバーを引く速度,レバーを引く時間の長さ,レバーの軌道の安定性と関係し,第3層に入力するブトンにおける神経活動はレバーを引く成功率と関係した.また,第1層に入力するブトンにおける神経活動および第3層に入力するブトンにおける神経活動は大脳基底核あるいは小脳に強い影響をうけ,一方,それらの神経活動のダイナミクスには大脳基底核および小脳の神経活動が必要であった.
はじめに
ピアノや手術といった高度な熟練の必要なものから,文字の入力や食器洗いなどの日常の動作にいたるまで,何度も何度もくり返して訓練することで目的の達成のための効率のよい運動スキルが習得されることを,われわれは日常的に経験している.これまで,1次運動野のニューロンは学習によりダイナミックな神経活動の変化を示すことが明らかにされているが1,2),運動学習中には脳のさまざまな領域において大きな変化が起こるため,運動学習の機構を解明するためには脳の領域のあいだの情報のやりとりを調べることが非常に重要である.大脳基底核は脳の深部に存在する複数の神経核からなり,運動の制御,認知機能,動機づけ,報酬などさまざまな機能を担い,また,小脳は知覚と運動機能を統合し細かい運動の制御にかかわる3).大脳基底核および小脳の障害は運動失調および運動障害疾患をもたらす.大脳基底核あるいは小脳からのシグナルは視床を介して運動野などの大脳皮質に送られ4,5),最終的に1次運動野の皮質脊髄投射ニューロンが脊髄へと出力を送り運動を遂行する.この運動系の経路において,視床は非常に重要なハブととらえられる.筆者らは,視床から運動野へと送られるシグナルの時間的なパターンを調べることは運動学習の機構の解明において大きな足がかりになると考え,視床から1次運動野へとシグナルを送る経路に着目した.
視床には核とよばれる細胞体の集団が多く存在するが,運動野へと投射する視床核のうち,前腹側核や内側腹側核から運動野への投射は第1層に多く分布し,外側腹側核からの投射は第3層や第5層に分布する6,7).また,前腹側核や内側腹側核は大脳基底核からの投射を多くうけ,外側腹側核は小脳からの投射を多くうける8).投射パターンの異なる視床皮質軸索の神経活動は,それぞれ,運動学習中にどのような活動パターンを示すのであろうか.この研究において,筆者らは,視床皮質軸索からの投射パターンに解剖学的な違いのあることを確認したうえで,マウスにレバーを自発的に一定の時間だけ引くと報酬として水がもらえるという運動課題を実施させ,どのようなシグナルが視床から運動野へと送られるかを調べた.
1.光遺伝学的な手法を用いた視床皮質軸索の活性化による運動の誘発および視床の抑制によるレバーを引く成功率の低下
光遺伝学的な手法を用いて1次運動野へ投射する視床皮質軸索の神経活動を活性化させた.視床皮質軸索にアデノ随伴ウイルスを用いて光駆動性の非選択的なカチオンチャネルであるチャネルロドプシン2を発現させ,頭上から1次運動野に光を照射し視床皮質軸索における神経活動を活性化したところ,それにともない手の運動が誘発された.また,光遺伝学的な手法を用いて視床の神経活動を抑制した.運動野に投射する視床核のニューロンにアデノ随伴ウイルスを用いて光駆動性のプロトンポンプであるアーキロドプシンTを発現させ,レバー引き課題を学習させたのち,視床へ光を照射し視床核のニューロンにおける神経活動を抑制したところ,レバーを引く成功率が有意に低下した.このことから,視床から運動野へと投射する経路に手の動きそのものやレバーを一定の時間だけ引くことに関連する経路が存在することがわかった.
2.レバー引き課題を実行しているときの1次運動野の視床皮質軸索における神経活動のダイナミクス
レバー引き課題を実行しているときの視床皮質軸索のブトンにおける神経活動を運動野において観察するため,2光子顕微鏡を用いたCa2+イメージング法を実施した.従来の電気生理学的な手法ではブトンだけのシグナルを分離することは不可能であったが,アデノ随伴ウイルスを用いて運動野に投射する視床皮質軸索にCa2+応答タンパク質であるGCaMP6f 9) を発現させ,ブトンにおけるCa2+濃度の上昇を蛍光輝度の変化により検出した.レバーを引くと脳はゆれるため,わずか数μmのブトンにおける神経活動をとらえることは困難であったが,運動野に赤色蛍光タンパク質を同時に発現させ,さらに,深さ方向に4μmごとの3つの平面でイメージング画像を取得し,赤色の蛍光輝度をもとにオフラインで3次元のゆれを補正することにより,ゆれによるアーティファクトを最小限に抑えてイメージング画像を解析することに成功した.このようにして,視床皮質軸索のブトンにおける神経活動を運動野の第1層および第3層において測定し,レバー引き課題を実行しているときの神経活動を学習の前後で比較した.
レバーを引くときの第1層および第3層の視床軸索のブトンにおける神経活動からレバーを引く軌道を予測しその精度を比較すると,学習をへてどちらも予測の精度が高くなった.また,セッション内の神経活動の再現性も学習をへて高くなった.このことは,学習により視床皮質軸索の神経活動がレバーを引く情報をもつようになったことを意味した.さらに,レバーを引くときの神経活動のタイミングについて解析した.その結果,学習の後期には第1層の視床皮質軸索においてはレバー引きの開始時と終了時に顕著な神経活動を示すブトンが多くなり,第3層においてはレバー引きの開始時に顕著な神経活動を示すブトンが多くなった.さらに,レバーを引くときのブトンの集団の神経活動のシークエンス構造に着目した.視野から2つのブトンをとりだしたとき,レバーを引くときに平均の神経活動が先行していたブトンにおける神経活動からもう一方のブトンにおける神経活動を予測できるかを調べ,有意な予測の精度をもつブトンのペアを定めた.レバーを引く期間のなかで,このようなブトンのペアがランダムよりも多く検出される連続した期間をシークエンス長とした.その結果,第1層のブトンは第3層のブトンよりも長いシークエンス長をもつ神経活動の構造を示すことがわかった.これらの結果より,レバー引き課題の学習に関係する第1層および第3層のブトンは,レバーを引くときに異なるダイナミクスを示すことが明らかにされた.
3.行動と神経活動との関係
ここまでに示された第1層および第3層のブトンにおける神経活動のダイナミクスは個体の行動において何を表象しているのであろうか.この問題に答えるため,ブトンにおける神経活動とレバーを引く行動との関係について調べた.ブトンにおける神経活動のダイナミクスを特徴づける量として,レバー引き開始時および終了時の神経活動,および,シークエンス長を用い,レバーを引く成功率,レバーを引く速度およびレバーを戻す速度,レバーを引く時間の長さ,おのおののセッションにおけるレバーの軌道の再現性といった行動を特徴づける量が統計的に説明されるかどうか検討した.行動は学習の初期から後期にかけて変化する可能性があるため,この効果についてはすべてのモデルに組み込んだうえでおのおのの量の効果を判定した.統計モデルとしては一般化線形モデルを用い,最適なモデルは赤池情報量基準を用いて選んだ.その結果,第1層のブトンにおける神経活動はレバーを引く速度,レバーを引く時間,レバーの軌道の再現性,つまり,レバーを引くキネマティクスに関係し,第3層のブトンにおける神経活動はレバーを引く成功率に関係することが明らかにされた.
4.大脳基底核および小脳が視床皮質軸索の神経活動におよぼす影響
第1層および第3層の視床皮質軸索のブトンにおける神経活動のダイナミクスが生じる原因は何であろうか.視床は大脳基底核および小脳からの入力を強くうけることがわかっていたことから,大脳基底核の出力を担う黒質網様部あるいは小脳核のニューロンを光遺伝学的な手法により活性化したときの視床皮質軸索のブトンの応答性をみた.その結果,黒質網様部を活性化したときは第1層のブトンが第3層のブトンよりも高率に応答し,小脳核を活性化したときは第3層のブトンが第1層のブトンよりも高率に反応した.これは,解剖学的な結果とも矛盾しなかった.さらに,線条体あるいは小脳核を破壊したマウスにレバー引き課題を実施させ視床皮質軸索のブトンにおける神経活動を測定したところ,いずれもレバー引き課題は正常に学習されず,ブトンにおける神経活動のダイナミクスについても第1層および第3層においてきわだった特徴は現われなかった.このことから,レバー引き課題の学習をとおして視床皮質軸索のブトンにおいて神経活動のダイナミクスが現われるには,大脳基底核および小脳の両方の神経活動が必要とされることが明らかにされた.
おわりに
この研究において,筆者らは,これまでの解剖学的な視床皮質からの投射経路の知見から,運動学習における層に依存的な機能の違いを予測し,その実証を試みた.その結果,運動学習をへて視床から運動野へは少なくとも2種類のシグナルが層に依存的に入力されることが明らかにされた.それらは大脳基底核あるいは小脳からのシグナルの影響を強くうけるものであり,行動と神経活動との関係も明らかにされた(図1).
今回の研究を発展させるためには,以下の問題に取り組む必要があるだろう.レバー引き課題を実行しているときに,視床皮質の投射経路に選択的に神経活動を抑制すると個体の行動はどのように変化するのか.第1層のブトンにおける神経活動はレバーを引くキネマティクス,第3層のブトンにおける神経活動はレバーを引く成功率と関係するという結果であったが,その因果関係は得られるかどうか.また,大脳基底核あるいは小脳のどちらかが破壊されると両方のシグナルが消失したことから,レバー引き課題の学習中には互いに影響をうけると考えられるが,どのようにしてこれらのシグナルは運動野にて統合されるのか,という点である.
運動学習中に大脳基底核あるいは小脳からそれぞれ視床を介して入力するシグナルが運動野において統合され,その結果が脊髄へと出力されると同時に,ふたたび大脳基底核および小脳へと送られる.そして,またそれぞれのシグナルが視床をとおり運動野へと送られる.このようにして形成されるループ構造はくり返しの訓練により外界から修飾をうけ,さらに,脳における相互作用をとおししだいにきわだった特徴をもつシグナルを形成していくと考えられる.そして,それらのシグナルが最終的にそれぞれの役割を担い運動スキルは習得されると考えられる.大脳基底核系と小脳系との相互作用は神経科学における最近のトピックでもあり,近年,ますますその重要性が注目されている10).これらのことをひとつひとつ明らかにすることは,運動の機構の解明や病態の理解にも貢献すると考えらえる.
文 献
- Masamizu, Y., Tanaka, Y. R., Tanaka Y. H. et al.: Two distinct layer specific dynamics of cortical ensembles during learning of a motor task. Nat. Neurosci., 17, 987-994 (2014)[PubMed]
- Peters, A. J., Lee, J., Hedrick, N. G. et al.: Reorganization of corticospinal output during motor learning. Nat. Neurosci., 20, 1133-1141 (2017)[PubMed]
- Hikosaka, O., Nakahara, H., Rand, M. K. et al.: Parallel neural networks for learning sequential procedures. Trends Neurosci., 22, 464-471 (1999)[PubMed]
- Houk, J. C. & Wise, S. P.: Distributed modular architectures linking basal ganglia, cerebellum, and cerebral cortex: their role in planning and controlling action. Cereb. Cortex, 5, 95-110 (1995)[PubMed]
- Middleton, F. A. & Strick, P. L.: Basal ganglia and cerebellar loops: motor and cognitive circuits. Brain Res. Brain Res. Rev., 31, 236-250 (2000)[PubMed]
- Kuramoto, E., Furuta, T., Nakamura, K. C. et al.: Two types of thalamocortical projections from the motor thalamic nuclei of the rat: a single neuron-tracing study using viral vectors. Cereb. Cortex, 19, 2065-2077 (2009)[PubMed]
- Kuramoto, E., Ohno, S., Furuta, T. et al.: Ventral medial nucleus neurons send thalamocortical afferents more widely and more preferentially to layer 1 than neurons of the ventral anterior-ventral lateral nuclear complex in the rat. Cereb. Cortex, 25, 221-235 (2015)[PubMed]
- Kuramoto, E., Fujiyama, F., Nakamura, K. C. et al.: Complementary distribution of glutamatergic cerebellar and GABAergic basal ganglia afferents to the rat motor thalamic nuclei. Eur. J. Neurosci., 33, 95-109 (2011)[PubMed]
- Chen, T. W., Wardill, T. J., Sun, Y. et al.: Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature, 499, 295-300 (2013)[PubMed]
- Bostan, A. C. & Strick, P. L.: The basal ganglia and the cerebellum: nodes in an integrated network. Nat. Rev. Neurosci., 19, 338-350 (2018)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2012年 京都大学大学院医学研究科にて博士号取得,同年 基礎生物学研究所 博士研究員,2016年 東京大学大学院医学系研究科 博士研究員を経て,2017年より同 特任助教.
研究テーマ:脳のなかの基盤とそれにより実現される機能との関係.
関心事:主観的な時間の長さ.
田中 康裕(Yasuhiro R. Tanaka)
略歴:2012年 京都大学大学院医学研究科にて博士号取得,同年 基礎生物学研究所 博士研究員を経て,2016年より東京大学大学院医学系研究科 助教.
松崎政紀(Masanori Matsuzaki)
東京大学大学院医学系研究科 教授.
研究室URL:http://plaza.umin.ac.jp/~Matsuzaki-Lab/index.html
© 2018 田中康代・田中康裕・松崎政紀 Licensed under CC 表示 2.1 日本
