シャジクモのゲノムの解読が明かす陸上植物への道および独自の進化
西山智明1・坂山英俊2
(1金沢大学学際科学実験センター ゲノム機能解析分野,2神戸大学大学院理学研究科 生物学専攻生物多様性講座)
email:西山智明,坂山英俊
DOI: 10.7875/first.author.2018.077
The Chara genome: secondary complexity and implications for plant terrestrialization.
Tomoaki Nishiyama, Hidetoshi Sakayama, Jan de Vries, Henrik Buschmann, Denis Saint-Marcoux, Kristian K. Ullrich, Fabian B. Haas, Lisa Vanderstraeten, Dirk Becker, Daniel Lang, Stanislav Vosolsobě, Stephane Rombauts, Per K.I. Wilhelmsson, Philipp Janitza, Ramona Kern, Alexander Heyl, Florian Rümpler, Luz Irina A. Calderón Villalobos, John M. Clay, Roman Skokan, Atsushi Toyoda, Yutaka Suzuki, Hiroshi Kagoshima, Elio Schijlen, Navindra Tajeshwar, Bruno Catarino, Alexander J. Hetherington, Assia Saltykova, Clemence Bonnot, Holger Breuninger, Aikaterini Symeonidi, Guru V. Radhakrishnan, Filip Van Nieuwerburgh, Dieter Deforce, Caren Chang, Kenneth G. Karol, Rainer Hedrich, Peter Ulvskov, Gernot Glöckner, Charles F. Delwiche, Jan Petrášek, Yves Van de Peer, Jiri Friml, Mary Beilby, Liam Dolan, Yuji Kohara, Sumio Sugano, Asao Fujiyama, Pierre-Marc Delaux, Marcel Quint, Günter Theißen, Martin Hagemann, Jesper Harholt, Christophe Dunand, Sabine Zachgo, Jane Langdale, Florian Maumus, Dominique Van Der Straeten, Sven B. Gould, Stefan A. Rensing
Cell, 174, 448-464.e24 (2018)
この研究において,筆者らは,陸上植物に近縁な藻類であるシャジクモのゲノムを解読した.陸上植物およびシャジクモより遠縁のクレブロソルミディウムとの比較から,シャジクモと陸上植物の共通祖先において獲得された遺伝子,シャジクモの系統と陸上植物の系統が分岐してから陸上植物の系統において獲得された遺伝子,および,シャジクモの系統における独自の進化について明らかにされた.
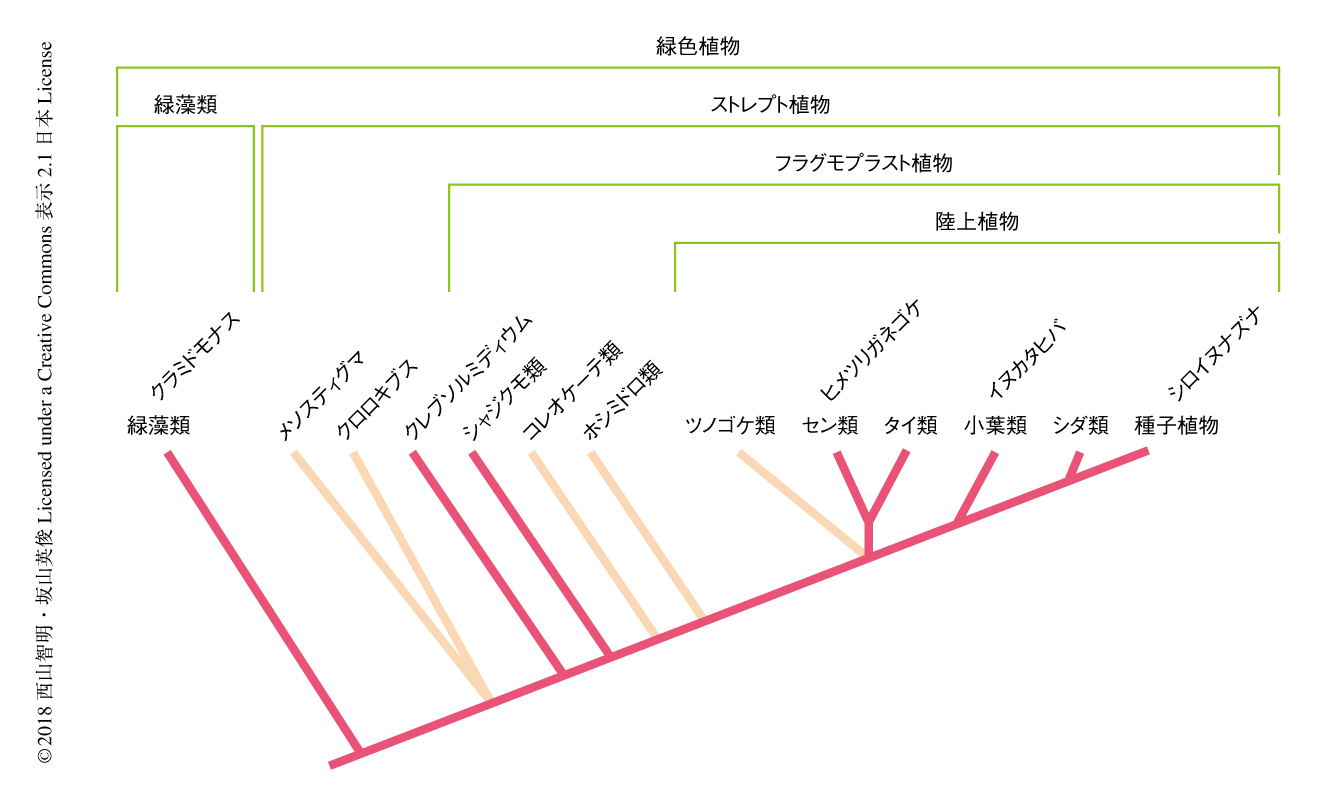
陸上植物とよばれる実際に陸上の植生の大部分をしめる植物は,2倍体からなる多細胞のからだを形成することを特徴とし,4.7億年~5億年まえに成立したと考えられている.陸上植物が成立して初期に分岐したコケ植物のヒメツリガネゴケ1),小葉類のイヌカタヒバ,被子植物のシロイヌナズナ,緑藻類のクラミドモナスのゲノムを比較することにより,被子植物の発生にかかわる遺伝子について多くがヒメツリガネゴケに相同遺伝子が存在すること,また,陸上植物における遺伝子ファミリーの多くは緑藻類と陸上植物が分岐したのちに陸上植物へとつながる系統において獲得されたことが示されている2).緑色植物は初期にクラミドモナスを含む緑藻類と陸上植物を含むストレプト植物とに分岐し,ストレプト植物には陸上植物への系統から分岐した複数の藻類の系統がある.そのような藻類のゲノムを解読し比較することにより,陸上植物が進化する過程において,いつ,どの遺伝子ファミリーが獲得されたかをより明瞭に知ることができる.この研究において,筆者らは,そうした系統のなかでも複雑な体制をもつシャジクモ(Chara braunii)のゲノムを解読した.シャジクモは19世紀半ばより陸上植物との近縁性が指摘されており,近年まで陸上植物にもっとも近い藻類と考えられていたが,最近の系統解析によりホシミドロ類,コレオケーテ類についで3番目に近い系統に属すると考えられている3)(図1).シャジクモのゲノムを解読しているあいだ,シャジクモよりまえに分岐したクレブソルミディウムのゲノムが解読されたことから4),緑藻類,クレブソルミディウム,シャジクモ,陸上植物をゲノムレベルで比較できるようになった.
シャジクモのゲノムを解読するにあたり,シャジクモ類のうちどの種を対象とするかを検討した.望ましい条件は,ゲノムのサイズが小さく,実験室において生活環のすべてが進行することである.また,純粋培養になっているとよいが,せめて,ほかの藻類が入らない条件で培養しないと解読された配列がどの生物に由来するのかわからなくなる.この問題を解決し,野外から採集した卵胞子を実験室において発芽させることによりシャジクモの単藻培養系を確立した.ただし,発芽および培養にはオートクレーブした土を用いており,無菌条件にはならなかった.土+水あるいは寒天ゲル+液相の2層培地で培養し1.5~3カ月で次世代の卵胞子が成熟した.
シャジクモ類のゲノムのサイズの推定値はたいてい大きいが,シャジクモは比較的小さく約2 Gbと推定された.とはいっても,ヒメツリガネゴケの4倍,シロイヌナズナの20倍である.インサート長を約250 bpに調製したペアエンドライブラリーを作製して塩基配列を決定した.
大きいゲノムの解読においては重複配列が問題となり,よいアセンブリーを得るためには重複配列をこえるインサート長のメイトペアライブラリーを作製することが肝になる.結果として,約17 kbまでのインサート長のライブラリーを19個ほど作製し塩基配列を決定した.得られたアセンブリーは28,091スカフォールド,合計1.99 Gbであった.ゲノムの解読に用いたS276株は無菌条件ではなく,したがって,このスカフォールドには共存する細菌に由来する配列も含まれていた.これらを区別するため,べつのS277株から得られたペアエンドリードのデータと比較した.S277株は同種であり,別の産地から得られた株ではあるがゲノム配列は共通で基本的に同じ割合で含まれるはずである.一方,細菌の配列は培養株の確立からDNAの調製にいたる過程において異なる割合で混入される.おのおののスカフォールドにマッピングされるS276株に由来するリード数およびS277株に由来するリード数をプロットしたところ,シャジクモの核のゲノムに相当すると考えられる対角線の付近に分布するスカフォールドと,S277株に由来するリード数の少ないスカフォールドとに分かれたことから,S277株に由来するリードの少ないスカフォールドは細菌に由来すると推定した.シャジクモに由来するとした配列と細菌に由来するとした配列はGC含量の分布も異なっていた.
アノテーションにおいては,シャジクモの完全長cDNAライブラリーのESTおよび生活環の多様なステージにおいて実施したRNA-seq法のデータを用いた.それにくわえ,陸上植物のヒメツリガネゴケおよびシロイヌナズナのタンパク質データセットが類似性を示す領域をヒント情報として遺伝子構造予測プログラムAugustusにあたえ,その結果,さきに細菌に由来するとみなされたスカフォールドを除くスカフォールドに予想された35,883遺伝子座,36,877モデルが得られた.なお,36,877モデルのうち13,331モデルについては転移因子との重なりがあったことから,タンパク質をコードしないかもしれないとして除いた数が23,546モデルである.のちの解析において細菌に由来すると判定したものを除き,一部は手作業で修正して,35,422遺伝子を国際塩基配列データベースに登録した.
シャジクモの遺伝子構造の特徴として,イントロンが非常に大きいという点があった.シャジクモの平均のイントロン長は約5.9 kbであり,ゲノムのサイズが4.8 Gbのオオムギにおける平均のイントロン長は約775 bp,ゲノムのサイズが2.1 Gbのトウモロコシにおける平均のイントロン長は746 bpであるのに比べ,1桁も大きな値であった.
シャジクモがクレブソルミディウムより陸上植物に近い形態的な特徴として,細胞分裂の際にフラグモプラストを形成するという点がある.フラグモプラストとは,新しく形成される細胞壁に対し垂直な微小管の束である.クレブソルミディウムなどにおいては細胞膜がくびれて新しい細胞壁に平行な微小管の束が観察されるが,新しい細胞壁に垂直な微小管の束はみられない.陸上植物において知られている細胞分裂にかかわる221個の遺伝子について調べたところ,TANGLED1遺伝子を除くすべての遺伝子がシャジクモよりまえに存在した.TANGLED1は前期前微小管束に局在し,新しく形成される細胞壁の方向の制御にかかわるタンパク質である.
陸上植物においてはさまざまな植物ホルモンにより発生および環境応答が制御されることが知られている.そこで,植物ホルモンの生合成,受容,シグナル伝達にかかわる遺伝子について調べた.その結果,シャジクモのゲノムにはエチレンのシグナル伝達にかかわるEIN2,オーキシンの応答にかかわるIAA,ARFファミリー転写因子をコードする遺伝子が存在し,これらはシャジクモが分岐するよりまえに獲得されたことがわかった.一方,オーキシン受容体TIR,アブシジン酸の生合成にかかわるNCED,アブシジン酸受容体PYR,ストリゴラクトンおよびカリキンの応答に関与するFボックスタンパク質MAX2をコードする遺伝子はシャジクモのゲノムにはみつからず,シャジクモと陸上植物が分岐してから陸上植物にいたる系統において獲得されたものと推定された.陸上植物において多様な機能をはたす転写因子の含まれるGRASファミリー転写因子をコードする遺伝子についても,シャジクモと陸上植物が分岐したのちに獲得されたと推定された.
植物は葉緑体において光合成の明反応を行うが,葉緑体において機能するタンパク質の多くは核のゲノムにコードされる.このため,葉緑体の状態に応じ核のゲノムにコードされる遺伝子の発現を制御するしくみが存在し,このシグナル伝達系のことをレトログレードシグナル伝達系とよぶ.レトログレードシグナル伝達系はGUN遺伝子として同定された複数の遺伝子により担われるが,GUN2遺伝子およびGUN3遺伝子はクレブソルミディウムにもある一方,中心的な役割を担うGUN1遺伝子はクレブソルミディウムにはないがシャジクモゲノムに存在し,したがって,シャジクモと陸上植物の共通祖先においてクレブソルミディウムと分岐したのちに獲得されたものと考えられた.
シャジクモの接合子はコケ植物およびシダ植物の胞子や種子植物の種子のように長期間にわたり休眠し乾燥環境に耐えることができる.これは,生活環における位置が異なり非相同であるが,LEA,Cupin,Oleosinなどの特徴的なタンパク質の発現は類似することがわかった.
シャジクモの節間細胞はひとつの細胞の長さが数cmにもおよび,細胞における原形質流動がはっきり観察できることで知られている.この原形質流動はアクチン-ミオシン系が担っており,シャジクモのゲノムには16個のアクチン遺伝子が検出された.クラミドモナス,クレブソルミディウム,ゼニゴケ,シロイヌナズナにおいては7~9個しかないのに比べ多く,大きい細胞あるいは原形質流動に関連して独自に増加したのかもしれない.
シャジクモの転写因子は,ファミリーごとにみるとおおむね陸上植物より少ない数のメンバーからなっていたが,Trihelix転写因子にかぎっては陸上植物で30個前後のところ302個もみつかった.なぜこれだけ増加したのかはわからないが,増加したのはシャジクモの系統に分岐してからであり,シャジクモの独自の進化にかかわる可能性がある.
陸上植物はキチンを検出して病原体や有益な微生物を認識するLysM型受容体キナーゼを多くもつ.LysM型受容体キナーゼはトランスクリプトーム解析ではひとつだけ見い出されており5),多様な相手を識別することはできないか,あるいは,識別はほかのシステムに依存すると考えられていたが,シャジクモのゲノムには7つのLysM型受容体キナーゼがコードされていた.これらは,いずれもシャジクモの系統において遺伝子重複により増加したと推定され,シャジクモに独自の微生物相互作用の系が存在するのかもしれない.LysM型受容体キナーゼのひとつは接合子においてとくに発現が高く,発芽するかどうかの制御に関与するのかもしれない.
シャジクモは生殖器官として造精器および生卵器という複雑な構造を形成する.この構造が陸上植物の造精器および造卵器と相同であるかどうかは必ずしも明らかではないが,シャジクモと陸上植物の共通祖先の段階において精子および卵という異形配偶子が形成されており,ホシミドロ類にいたる系統において,あとからほぼ同じ大きさおよびかたちの配偶子が接合するように退化したと考えられている.造精器のトランスクリプトーム解析においては,ダイニンをコードする遺伝子など鞭毛運動にかかわる遺伝子の発現が高くなっており鞭毛をもつ精子が分化する器官らしさがみられた.また,クラミドモナスにおいて配偶子の分化に関与するMID,および,ゼニゴケにおいて卵細胞が受精のまえに分裂するのを抑制するのに必要なMpRKD 6,7) を含むRWP-RK転写因子ファミリーに属する遺伝子のひとつの発現が造精器において高いことがわかった.生卵器においてはとくに活性酸素種にかかわる遺伝子の発現がめだち,ペルオキシダーゼであるクラスIII Prxの発現が高くなっていた.Prxについては遺伝子の数もクレブソルミディウムより多くなっており,とくにシャジクモの系統における遺伝子重複により増加していた.Prxは被子植物においては細胞壁の成分の重合に関与することが知られており,接合子の特別に厚い細胞壁の形成に関与するのかもしれない.また,生卵器においてとくに発現の高いPrxが多く,有性生殖の制御にかかわるのかもしれない.
この研究において,筆者らは,シャジクモの概要ゲノム配列を解読し,そこからわかることの一端を解析した.直接には扱っていない遺伝子ファミリーの解析においても,シャジクモを含め考察することにより陸上植物の進化の理解を助けることが期待される.ゲノムの進化を探求するうえでは,さらに,ストレプト植物の基部において分岐したメソスティグマ,クロロキブス,および,シャジクモより陸上植物に近縁なコレオケーテ類,ホシミドロ類のゲノムの解読を進めることがつぎのステップとなろう.
シャジクモのゲノム配列については,さらに精度を高め,染色体スケールの仮想染色体を組み立てることがひとつの課題である.今回,ゲノムが解読された茨城県霞ヶ浦産のS276株と愛媛県西条産のS277株の交配から遺伝学的な連鎖地図を作成して,これにもとづき構築が可能であろうと期待される.また,アセンブリーギャップを埋めていくことが望ましい.
シャジクモのリファレンス配列が得られたことにより,多様なシャジクモのゲノムにおける遺伝的なバリエーションを調べることが容易になった.シャジクモは世界中に分布し,それぞれ多様な環境に適応している.たとえば,国内においても,浅い水系と深い水系とで異なる遺伝型に分化していることが報告されている8).この遺伝的な分化のみられる集団について,形態的な違いはあまり明瞭でないとされるが,浅水型であるS276株と深水型であるS277株を同じ光環境で生育させると,S276株の節間が長くなる光強度においてS277株は節間がより短いなど明瞭に異なる形態になり,おそらく,深水型のほうが光感受性は高いと考えられる.今後は,こういった適応進化にかかわる遺伝的なバリエーションを探索できるようになると期待される.
また,シャジクモにおいて遺伝子の機能を解析する系の確立が重要である.ゲノム配列はわかっていても,そこにある遺伝子の機能を調べられないのは隔靴掻痒である.ヒメツリガネゴケなどに比べてもイントロンが非常に大きく機能解析はむずかしそうではあるが,形質転換さえ実現すれば,CRISPR-Cas9系を利用したゲノム編集法によりある程度の解析はできるのではないかと期待される.
略歴:2000年 東京大学大学院理学系研究科博士課程 修了,同年 基礎生物学研究所 研究員を経て,2005年より金沢大学学際科学実験センター 助手(現 助教).
研究テーマ:植物の進化.
坂山 英俊(Hidetoshi Sakayama)
神戸大学大学院理学研究科 准教授.
© 2018 西山智明・坂山英俊 Licensed under CC 表示 2.1 日本
(1金沢大学学際科学実験センター ゲノム機能解析分野,2神戸大学大学院理学研究科 生物学専攻生物多様性講座)
email:西山智明,坂山英俊
DOI: 10.7875/first.author.2018.077
The Chara genome: secondary complexity and implications for plant terrestrialization.
Tomoaki Nishiyama, Hidetoshi Sakayama, Jan de Vries, Henrik Buschmann, Denis Saint-Marcoux, Kristian K. Ullrich, Fabian B. Haas, Lisa Vanderstraeten, Dirk Becker, Daniel Lang, Stanislav Vosolsobě, Stephane Rombauts, Per K.I. Wilhelmsson, Philipp Janitza, Ramona Kern, Alexander Heyl, Florian Rümpler, Luz Irina A. Calderón Villalobos, John M. Clay, Roman Skokan, Atsushi Toyoda, Yutaka Suzuki, Hiroshi Kagoshima, Elio Schijlen, Navindra Tajeshwar, Bruno Catarino, Alexander J. Hetherington, Assia Saltykova, Clemence Bonnot, Holger Breuninger, Aikaterini Symeonidi, Guru V. Radhakrishnan, Filip Van Nieuwerburgh, Dieter Deforce, Caren Chang, Kenneth G. Karol, Rainer Hedrich, Peter Ulvskov, Gernot Glöckner, Charles F. Delwiche, Jan Petrášek, Yves Van de Peer, Jiri Friml, Mary Beilby, Liam Dolan, Yuji Kohara, Sumio Sugano, Asao Fujiyama, Pierre-Marc Delaux, Marcel Quint, Günter Theißen, Martin Hagemann, Jesper Harholt, Christophe Dunand, Sabine Zachgo, Jane Langdale, Florian Maumus, Dominique Van Der Straeten, Sven B. Gould, Stefan A. Rensing
Cell, 174, 448-464.e24 (2018)
要 約
この研究において,筆者らは,陸上植物に近縁な藻類であるシャジクモのゲノムを解読した.陸上植物およびシャジクモより遠縁のクレブロソルミディウムとの比較から,シャジクモと陸上植物の共通祖先において獲得された遺伝子,シャジクモの系統と陸上植物の系統が分岐してから陸上植物の系統において獲得された遺伝子,および,シャジクモの系統における独自の進化について明らかにされた.
はじめに
陸上植物とよばれる実際に陸上の植生の大部分をしめる植物は,2倍体からなる多細胞のからだを形成することを特徴とし,4.7億年~5億年まえに成立したと考えられている.陸上植物が成立して初期に分岐したコケ植物のヒメツリガネゴケ1),小葉類のイヌカタヒバ,被子植物のシロイヌナズナ,緑藻類のクラミドモナスのゲノムを比較することにより,被子植物の発生にかかわる遺伝子について多くがヒメツリガネゴケに相同遺伝子が存在すること,また,陸上植物における遺伝子ファミリーの多くは緑藻類と陸上植物が分岐したのちに陸上植物へとつながる系統において獲得されたことが示されている2).緑色植物は初期にクラミドモナスを含む緑藻類と陸上植物を含むストレプト植物とに分岐し,ストレプト植物には陸上植物への系統から分岐した複数の藻類の系統がある.そのような藻類のゲノムを解読し比較することにより,陸上植物が進化する過程において,いつ,どの遺伝子ファミリーが獲得されたかをより明瞭に知ることができる.この研究において,筆者らは,そうした系統のなかでも複雑な体制をもつシャジクモ(Chara braunii)のゲノムを解読した.シャジクモは19世紀半ばより陸上植物との近縁性が指摘されており,近年まで陸上植物にもっとも近い藻類と考えられていたが,最近の系統解析によりホシミドロ類,コレオケーテ類についで3番目に近い系統に属すると考えられている3)(図1).シャジクモのゲノムを解読しているあいだ,シャジクモよりまえに分岐したクレブソルミディウムのゲノムが解読されたことから4),緑藻類,クレブソルミディウム,シャジクモ,陸上植物をゲノムレベルで比較できるようになった.
1.シャジクモの単藻培養系の確立
シャジクモのゲノムを解読するにあたり,シャジクモ類のうちどの種を対象とするかを検討した.望ましい条件は,ゲノムのサイズが小さく,実験室において生活環のすべてが進行することである.また,純粋培養になっているとよいが,せめて,ほかの藻類が入らない条件で培養しないと解読された配列がどの生物に由来するのかわからなくなる.この問題を解決し,野外から採集した卵胞子を実験室において発芽させることによりシャジクモの単藻培養系を確立した.ただし,発芽および培養にはオートクレーブした土を用いており,無菌条件にはならなかった.土+水あるいは寒天ゲル+液相の2層培地で培養し1.5~3カ月で次世代の卵胞子が成熟した.
2.ゲノムの解読,アセンブリー,アノテーション
シャジクモ類のゲノムのサイズの推定値はたいてい大きいが,シャジクモは比較的小さく約2 Gbと推定された.とはいっても,ヒメツリガネゴケの4倍,シロイヌナズナの20倍である.インサート長を約250 bpに調製したペアエンドライブラリーを作製して塩基配列を決定した.
大きいゲノムの解読においては重複配列が問題となり,よいアセンブリーを得るためには重複配列をこえるインサート長のメイトペアライブラリーを作製することが肝になる.結果として,約17 kbまでのインサート長のライブラリーを19個ほど作製し塩基配列を決定した.得られたアセンブリーは28,091スカフォールド,合計1.99 Gbであった.ゲノムの解読に用いたS276株は無菌条件ではなく,したがって,このスカフォールドには共存する細菌に由来する配列も含まれていた.これらを区別するため,べつのS277株から得られたペアエンドリードのデータと比較した.S277株は同種であり,別の産地から得られた株ではあるがゲノム配列は共通で基本的に同じ割合で含まれるはずである.一方,細菌の配列は培養株の確立からDNAの調製にいたる過程において異なる割合で混入される.おのおののスカフォールドにマッピングされるS276株に由来するリード数およびS277株に由来するリード数をプロットしたところ,シャジクモの核のゲノムに相当すると考えられる対角線の付近に分布するスカフォールドと,S277株に由来するリード数の少ないスカフォールドとに分かれたことから,S277株に由来するリードの少ないスカフォールドは細菌に由来すると推定した.シャジクモに由来するとした配列と細菌に由来するとした配列はGC含量の分布も異なっていた.
アノテーションにおいては,シャジクモの完全長cDNAライブラリーのESTおよび生活環の多様なステージにおいて実施したRNA-seq法のデータを用いた.それにくわえ,陸上植物のヒメツリガネゴケおよびシロイヌナズナのタンパク質データセットが類似性を示す領域をヒント情報として遺伝子構造予測プログラムAugustusにあたえ,その結果,さきに細菌に由来するとみなされたスカフォールドを除くスカフォールドに予想された35,883遺伝子座,36,877モデルが得られた.なお,36,877モデルのうち13,331モデルについては転移因子との重なりがあったことから,タンパク質をコードしないかもしれないとして除いた数が23,546モデルである.のちの解析において細菌に由来すると判定したものを除き,一部は手作業で修正して,35,422遺伝子を国際塩基配列データベースに登録した.
シャジクモの遺伝子構造の特徴として,イントロンが非常に大きいという点があった.シャジクモの平均のイントロン長は約5.9 kbであり,ゲノムのサイズが4.8 Gbのオオムギにおける平均のイントロン長は約775 bp,ゲノムのサイズが2.1 Gbのトウモロコシにおける平均のイントロン長は746 bpであるのに比べ,1桁も大きな値であった.
3.陸上への進出につながる特徴
シャジクモがクレブソルミディウムより陸上植物に近い形態的な特徴として,細胞分裂の際にフラグモプラストを形成するという点がある.フラグモプラストとは,新しく形成される細胞壁に対し垂直な微小管の束である.クレブソルミディウムなどにおいては細胞膜がくびれて新しい細胞壁に平行な微小管の束が観察されるが,新しい細胞壁に垂直な微小管の束はみられない.陸上植物において知られている細胞分裂にかかわる221個の遺伝子について調べたところ,TANGLED1遺伝子を除くすべての遺伝子がシャジクモよりまえに存在した.TANGLED1は前期前微小管束に局在し,新しく形成される細胞壁の方向の制御にかかわるタンパク質である.
陸上植物においてはさまざまな植物ホルモンにより発生および環境応答が制御されることが知られている.そこで,植物ホルモンの生合成,受容,シグナル伝達にかかわる遺伝子について調べた.その結果,シャジクモのゲノムにはエチレンのシグナル伝達にかかわるEIN2,オーキシンの応答にかかわるIAA,ARFファミリー転写因子をコードする遺伝子が存在し,これらはシャジクモが分岐するよりまえに獲得されたことがわかった.一方,オーキシン受容体TIR,アブシジン酸の生合成にかかわるNCED,アブシジン酸受容体PYR,ストリゴラクトンおよびカリキンの応答に関与するFボックスタンパク質MAX2をコードする遺伝子はシャジクモのゲノムにはみつからず,シャジクモと陸上植物が分岐してから陸上植物にいたる系統において獲得されたものと推定された.陸上植物において多様な機能をはたす転写因子の含まれるGRASファミリー転写因子をコードする遺伝子についても,シャジクモと陸上植物が分岐したのちに獲得されたと推定された.
植物は葉緑体において光合成の明反応を行うが,葉緑体において機能するタンパク質の多くは核のゲノムにコードされる.このため,葉緑体の状態に応じ核のゲノムにコードされる遺伝子の発現を制御するしくみが存在し,このシグナル伝達系のことをレトログレードシグナル伝達系とよぶ.レトログレードシグナル伝達系はGUN遺伝子として同定された複数の遺伝子により担われるが,GUN2遺伝子およびGUN3遺伝子はクレブソルミディウムにもある一方,中心的な役割を担うGUN1遺伝子はクレブソルミディウムにはないがシャジクモゲノムに存在し,したがって,シャジクモと陸上植物の共通祖先においてクレブソルミディウムと分岐したのちに獲得されたものと考えられた.
シャジクモの接合子はコケ植物およびシダ植物の胞子や種子植物の種子のように長期間にわたり休眠し乾燥環境に耐えることができる.これは,生活環における位置が異なり非相同であるが,LEA,Cupin,Oleosinなどの特徴的なタンパク質の発現は類似することがわかった.
4.シャジクモに独自の特徴
シャジクモの節間細胞はひとつの細胞の長さが数cmにもおよび,細胞における原形質流動がはっきり観察できることで知られている.この原形質流動はアクチン-ミオシン系が担っており,シャジクモのゲノムには16個のアクチン遺伝子が検出された.クラミドモナス,クレブソルミディウム,ゼニゴケ,シロイヌナズナにおいては7~9個しかないのに比べ多く,大きい細胞あるいは原形質流動に関連して独自に増加したのかもしれない.
シャジクモの転写因子は,ファミリーごとにみるとおおむね陸上植物より少ない数のメンバーからなっていたが,Trihelix転写因子にかぎっては陸上植物で30個前後のところ302個もみつかった.なぜこれだけ増加したのかはわからないが,増加したのはシャジクモの系統に分岐してからであり,シャジクモの独自の進化にかかわる可能性がある.
陸上植物はキチンを検出して病原体や有益な微生物を認識するLysM型受容体キナーゼを多くもつ.LysM型受容体キナーゼはトランスクリプトーム解析ではひとつだけ見い出されており5),多様な相手を識別することはできないか,あるいは,識別はほかのシステムに依存すると考えられていたが,シャジクモのゲノムには7つのLysM型受容体キナーゼがコードされていた.これらは,いずれもシャジクモの系統において遺伝子重複により増加したと推定され,シャジクモに独自の微生物相互作用の系が存在するのかもしれない.LysM型受容体キナーゼのひとつは接合子においてとくに発現が高く,発芽するかどうかの制御に関与するのかもしれない.
シャジクモは生殖器官として造精器および生卵器という複雑な構造を形成する.この構造が陸上植物の造精器および造卵器と相同であるかどうかは必ずしも明らかではないが,シャジクモと陸上植物の共通祖先の段階において精子および卵という異形配偶子が形成されており,ホシミドロ類にいたる系統において,あとからほぼ同じ大きさおよびかたちの配偶子が接合するように退化したと考えられている.造精器のトランスクリプトーム解析においては,ダイニンをコードする遺伝子など鞭毛運動にかかわる遺伝子の発現が高くなっており鞭毛をもつ精子が分化する器官らしさがみられた.また,クラミドモナスにおいて配偶子の分化に関与するMID,および,ゼニゴケにおいて卵細胞が受精のまえに分裂するのを抑制するのに必要なMpRKD 6,7) を含むRWP-RK転写因子ファミリーに属する遺伝子のひとつの発現が造精器において高いことがわかった.生卵器においてはとくに活性酸素種にかかわる遺伝子の発現がめだち,ペルオキシダーゼであるクラスIII Prxの発現が高くなっていた.Prxについては遺伝子の数もクレブソルミディウムより多くなっており,とくにシャジクモの系統における遺伝子重複により増加していた.Prxは被子植物においては細胞壁の成分の重合に関与することが知られており,接合子の特別に厚い細胞壁の形成に関与するのかもしれない.また,生卵器においてとくに発現の高いPrxが多く,有性生殖の制御にかかわるのかもしれない.
おわりに
この研究において,筆者らは,シャジクモの概要ゲノム配列を解読し,そこからわかることの一端を解析した.直接には扱っていない遺伝子ファミリーの解析においても,シャジクモを含め考察することにより陸上植物の進化の理解を助けることが期待される.ゲノムの進化を探求するうえでは,さらに,ストレプト植物の基部において分岐したメソスティグマ,クロロキブス,および,シャジクモより陸上植物に近縁なコレオケーテ類,ホシミドロ類のゲノムの解読を進めることがつぎのステップとなろう.
シャジクモのゲノム配列については,さらに精度を高め,染色体スケールの仮想染色体を組み立てることがひとつの課題である.今回,ゲノムが解読された茨城県霞ヶ浦産のS276株と愛媛県西条産のS277株の交配から遺伝学的な連鎖地図を作成して,これにもとづき構築が可能であろうと期待される.また,アセンブリーギャップを埋めていくことが望ましい.
シャジクモのリファレンス配列が得られたことにより,多様なシャジクモのゲノムにおける遺伝的なバリエーションを調べることが容易になった.シャジクモは世界中に分布し,それぞれ多様な環境に適応している.たとえば,国内においても,浅い水系と深い水系とで異なる遺伝型に分化していることが報告されている8).この遺伝的な分化のみられる集団について,形態的な違いはあまり明瞭でないとされるが,浅水型であるS276株と深水型であるS277株を同じ光環境で生育させると,S276株の節間が長くなる光強度においてS277株は節間がより短いなど明瞭に異なる形態になり,おそらく,深水型のほうが光感受性は高いと考えられる.今後は,こういった適応進化にかかわる遺伝的なバリエーションを探索できるようになると期待される.
また,シャジクモにおいて遺伝子の機能を解析する系の確立が重要である.ゲノム配列はわかっていても,そこにある遺伝子の機能を調べられないのは隔靴掻痒である.ヒメツリガネゴケなどに比べてもイントロンが非常に大きく機能解析はむずかしそうではあるが,形質転換さえ実現すれば,CRISPR-Cas9系を利用したゲノム編集法によりある程度の解析はできるのではないかと期待される.
文 献
- Rensing, S. A., Lang, D., Zimmer, A. D. et al.: The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science, 319, 64-69 (2008)[PubMed]
- Banks, J. A., Nishiyama, T., Hasebe, M. et al.: The Selaginella genome identifies genetic changes associated with the evolution of vascular plants. Science, 332, 960-963 (2011)[PubMed]
- Wickett, N. J., Mirarab, S., Nguyen, N. et al.: Phylotranscriptomic analysis of the origin and early diversification of land plants. Proc. Natl. Acad. Sci. USA, 111, E4859-E4868 (2014)[PubMed]
- Hori, K., Maruyama, F., Fujisawa, T. et al.: Klebsormidium flaccidum genome reveals primary factors for plant terrestrial adaptation. Nat. Commun., 5, 3978 (2014)[PubMed]
- Delaux, P. M., Radhakrishnan, G. V., Jayaraman, D. et al.: Algal ancestor of land plants was preadapted for symbiosis. Proc. Natl. Acad. Sci. USA, 112, 13390-13395 (2015)[PubMed]
- Rovekamp, M., Bowman, J. L. & Grossniklaus, U.: Marchantia MpRKD regulates the gametophyte-sporophyte transition by keeping egg cells quiescent in the absence of fertilization. Curr. Biol., 26, 1782-1789 (2016)[PubMed]
- Koi, S,, Hisanaga, T., Sato, K. et al.: An evolutionarily conserved plant RKD factor controls germ cell differentiation. Curr Biol., 26, 1775-1781 (2016)[PubMed]
- Kato, S., Sakayama, H., Sano, S. et al.: Morphological variation and intraspecific phylogeny of the ubiquitous species Chara braunii (Charales, Charophyceae) in Japan. Phycologia, 47, 191-202 (2008)
著者プロフィール
略歴:2000年 東京大学大学院理学系研究科博士課程 修了,同年 基礎生物学研究所 研究員を経て,2005年より金沢大学学際科学実験センター 助手(現 助教).
研究テーマ:植物の進化.
坂山 英俊(Hidetoshi Sakayama)
神戸大学大学院理学研究科 准教授.
© 2018 西山智明・坂山英俊 Licensed under CC 表示 2.1 日本
