海馬の記憶エングラムは経験の表象へのインデックスである
田中和正・Thomas J. McHugh
(理化学研究所脳神経科学研究センター 神経回路・行動生理学研究チーム)
email:田中和正
DOI: 10.7875/first.author.2018.076
The hippocampal engram maps experience but not place.
Kazumasa Z. Tanaka, Hongshen He, Anupratap Tomar, Kazue Niisato, Arthur J. Y. Huang, Thomas J. McHugh
Science, 361, 392-397 (2018)
エピソード記憶は海馬のニューロンが担う.マウスにおいて,記憶エングラムの光遺伝学的な操作により海馬のニューロンが記憶の想起に必要かつ誘導的であることが示されてきた.しかしながら,自由行動下のマウスにおける記憶エングラムの活動のパターンや,記憶における具体的な役割についてはまったくの未知である.この研究において,筆者らは,文脈記憶の記銘の際に海馬CA1野における場所細胞の一部が記憶エングラムになることを見い出した.これらの場所細胞は,記銘の際にシータ域の周波数にて特徴的なバースト活動をくり返していた.この活動のパターンはシナプスを増強させるのに効果的であることが知られている.さらに,記憶を想起する際の記憶エングラムの活動を解析したところ,予想とは逆に,記銘の際に表現していた空間の情報はまったく保持していなかった.しかしながら,記憶エングラムの活動量それ自体は,経験している文脈のアイデンティティをすばやく正確に反映していた.これらの結果から,海馬には異なる2つの記憶の痕跡が形成され,一方は正確な空間の情報を,もう一方はより広義な文脈の情報を保存することが示された.
海馬はエピソード記憶を担う.海馬を損傷した患者は日々経験する出来事を記憶すること(記憶の記銘)ができなくなり,また,損傷のまえに経験した事柄の多くを思い出すこと(記憶の想起)ができなくなる.この障害はヒト以外の多くの哺乳類においても起こることが実験的に示されている.
では,海馬はどのようにエピソード記憶を担うのであろうか? この問いは2つの意味をもつ.第1に,記憶の物理的な痕跡とは何かという問い.第2に,海馬が記憶の情報をどのように表現し,その情報が記憶の痕跡によりどのように修飾されることが記憶の記銘に対応するのかという問いである.第1の問いに対しては,シナプス可塑性,とくに,長期増強がその候補であると考えられており,現代の神経科学はこの見方を支持する証拠を積み上げてきた1).たとえば,シナプス可塑性のマーカーである最初期遺伝子の発現により定義されるニューロン,すなわち,記憶エングラムのスパイク活動がある種の記憶の想起に必要かつ誘導的であることが実験的に示されてきた2,3).また,光遺伝学的にスパインの収縮をひき起こすことにより,学習により増強されたスパインが運動の記憶の保持に必要であることも実証された4).しかしながら,それにつづく問い,すなわち,経験により形成された記憶の物理的な痕跡が神経活動,とくに,海馬が表現する情報をどのように安定化(あるいは,変化)させることにより記憶の情報の保存にいたるのか,という問題に対し,その理解はほとんど進んでいない.この問題を説明しようとする理論は数多く存在するものの,それら理論のあいだの整合性が実験的に検証されてきたとはいいがたく,ひとつひとつの理論を直接的に証明することも技術的にむずかしい.
海馬の役割について現在もっとも広く受け入れられている考え方は認知地図仮説であろう5).げっ歯類を含む哺乳類の海馬には,空間の特定の位置に対し発火する場所細胞が存在する.ある環境にいる動物が移動するとき,場所細胞は動物がある位置を通過したときのみ発火する.ひとつひとつの場所細胞は異なる場所受容野をもつため,場所細胞の集団は総体としてひとつの空間地図とみなすことができる.この空間地図は環境に特異的であり,同じ動物が別の環境を移動する際には,異なる組合せの場所細胞の集団が異なる場所受容野をもつことにより別の空間地図を表現する.この見方を拡張し,任意の認知領域における枠組みを表現するのが海馬の役割であると考えるのが認知地図仮説である(図1).では,記憶の物理的な痕跡,すなわち,シナプス可塑性はこうした情報の表現をどのように修飾するのであろうか? もっとも一般的な見方は,シナプス可塑性が位置情報の表現の安定化に寄与するというものである.実際に,シナプス可塑性に重要な役割をはたすNMDA型グルタミン酸受容体を薬理学的あるいは遺伝学的に阻害することにより,場所細胞の安定性は損なわれることが報告されている1).

しかしながら,認知地図仮説は海馬の役割を説明する唯一の理論ではない.なかでも,記憶インデックス仮説は認知地図仮説とは大きく性質が異なる6).記憶インデックス仮説は,海馬に保存される記憶の痕跡は記憶をよび起こすためのたんなるインデックス(索引)にすぎず,記憶の本体は皮質に保存されると提唱する.動物が経験する文脈を構成する刺激の組合せは海馬に記憶の痕跡を形成する.この海馬における記憶の痕跡は,皮質に保存されたそれぞれの刺激の表象とリンクしており,記憶の想起の際にはこれら皮質の表象を再活性化する.すなわち,海馬にはそれ単独で解読の可能な情報が保存されているわけではなく,海馬の役割はあくまで皮質の活動のパターンを再構成することであると考える(図1).
場所細胞についての知見は認知地図仮説を支持するが,文脈恐怖条件づけを用いた研究との整合性が高いのは記憶インデックス仮説である.なぜならば,海馬において文脈表象を形成するためには複数のモダリティからなる刺激を同時に経験する必要があり,文脈記憶を想起するために環境を物理的に探索する必要がないためである7).こうした海馬の文脈表象はしばしば最初期遺伝子の産物であるc-Fosの発現により定義されており,これらを標識あるいは操作することにより文脈記憶の想起が抑制あるいは惹起されることも示されている2,3).
以上のように,電気生理学的な手法を用いた場所細胞の研究と,文脈恐怖条件づけを用いた文脈表象の研究は,海馬の記憶の研究における二大パラダイムを構築してきたといえる.しかしながら,両者を統一的に説明する試みはいまだ成功をおさめていない.
このような背景のもと,エピソード記憶に対する海馬の役割をより深く理解するため,海馬の記憶エングラムからテトロード電極を用いた記録を行った.海馬CA1野においてc-Fosを発現するニューロンに光駆動性の陽イオンチャネルであるチャネルロドプシン2および蛍光タンパク質EYFPを発現するマウスを用い,海馬CA1野に光ファイバーおよびテトロード電極を搭載するマイクロドライブを埋め込んだ.このマウスに新規の文脈Aを探索させる(記銘)ことにより,c-Fosを発現する記憶エングラムをチャネルロドプシン2により標識した.翌日,マウスにもういちど文脈Aを探索させ(想起),場所細胞の活動の一貫性から海馬の認知地図の安定性を評価した.さらに12時間後,マイクロドライブに搭載した光ファイバーをとおして海馬CA1野を青色光により刺激し,最初の新規の文脈の探索の結果としてチャネルロドプシン2により標識されたニューロンを同定した.この光遺伝学的な同定により平均19.59%の海馬CA1野のニューロンが光応答性の発火を示し,したがって,c-Fos陽性細胞と同定された.この割合は過去の文献において報告された割合とも一致し8),また,記録ののち確認されたEYFP陽性細胞の割合とも近似していた.最後に,同じマウスに異なる文脈Bを探索させ,文脈に特異的な活動について調べた.
マウスが新規の文脈を探索する際に,記憶エングラムがマウスの位置に応答して発火するかどうか調べた.以前から想定されていたように,記憶エングラムの大部分は記銘した文脈において場所受容野をもっていた.しかしながら,記憶エングラムである場所細胞は全体のごく一部であり,記銘した文脈に応答した場所細胞の大半はチャネルロドプシン2により標識されていなかった.記憶エングラムとそれ以外の場所細胞は生理学的な性質の多くがよく似ており,たとえば,最大の発火率には有意な差はみられなかった.しかしながら,平均の発火率は記憶エングラムのほうが高かった.したがって,記憶エングラムの場所受容野はそれ以外の場所細胞よりも有意に大きく,その結果として,記憶エングラムによる発火ひとつひとつのもつ空間の情報量は小さかった.
文脈記憶を記銘する際の発火の時間的な構造を解析した.記憶エングラムの発火時間の間隔について解析したところ,記憶エングラムの発火はそれ以外の場所細胞よりもバーストして起こりやすいことがわかった.記憶エングラムはそれ以外の場所細胞よりもバースト発火率が有意に高く,バーストのあいだの時間間隔が有意に短かった.記憶エングラムのバースト活動の多くは約125ミリ秒の間隔で起こった.これは,海馬において顕著に観察される8 Hzのシータリズムに対応した.結果として,記憶エングラムのバースト活動はそれ以外の場所細胞よりも強くシータ変調されていた.この活動パターンをより定量的に扱うため,83~167ミリ秒(6~12 Hz)の間隔で連続して起こるバースト活動をシータバーストイベントと定義し,記憶エングラムとそれ以外の場所細胞の活動のパターンをさらにくわしく調べた.その結果,新規の文脈の探索において,記憶エングラムにおいてはそれ以外の場所細胞よりも有意に高い率でシータバーストイベントが起こっており,1回のシータバーストイベントにおいて連続して起こるバースト活動の数も有意に多かった.文脈Aの記銘において形成された記憶エングラムAにみられたこれら特徴的な活動のパターンは,マウスが文脈Bを探索する際には記憶エングラムAにおいてみられなかった.このことから,シータ域の波長にて反復して起こるバースト活動が文脈記憶の記銘に関与することが示唆された.
局所フィールド電位から検出されるニューロンの集団の活動がこれらの発火とどのように関連するのかを検証した.海馬CA1野のニューロンは,シータ波(6~12 Hz),遅いガンマ波(30~50 Hz),速いガンマ波(55~85 Hz)の3つのリズムの影響をうけて発火しており,ガンマ波の強度は海馬CA3野(遅いガンマ波)および嗅内皮質(速いガンマ波)からの入力の強度と相関する9).マウスが新規の文脈を探索する際,記憶エングラムを含むすべての場所細胞の発火はシータ波と同じ程度に同期しており,また,これまでに報告されていたように,シータ波の上昇相において優先的に発火していた.しかしながら,単独の発火ではなくシータバーストイベントと振動との関連を解析したところ,記憶エングラム以外の場所細胞におけるシータバーストイベントはシータ波の上昇相にとどまったままであったが,記憶エングラムのシータバーストイベントは下降相へとシフトしていた.海馬CA1野への入力がこれらの場所細胞におよぼす影響について調べるため,ガンマイベントの際の発火について解析した.記憶エングラムにおいては,より多くの発火が速いガンマイベントにおいて起こっており,有意に大きな割合の場所細胞が速いガンマ波と位相固定されていた.これらのガンマイベントにおける発火や位相固定における記憶エングラムとそれ以外の場所細胞とでの違いは,遅いガンマ波においては検出されなかった.これらの結果から,記憶エングラムもそれ以外の場所細胞も海馬CA3野からの入力を同じ程度にうけてはいるが,記憶エングラムは嗅内皮質からの入力に応答性が高く,それによりシータ波にともない起こるシータバーストイベントの位相がシフトする可能性が示唆された.
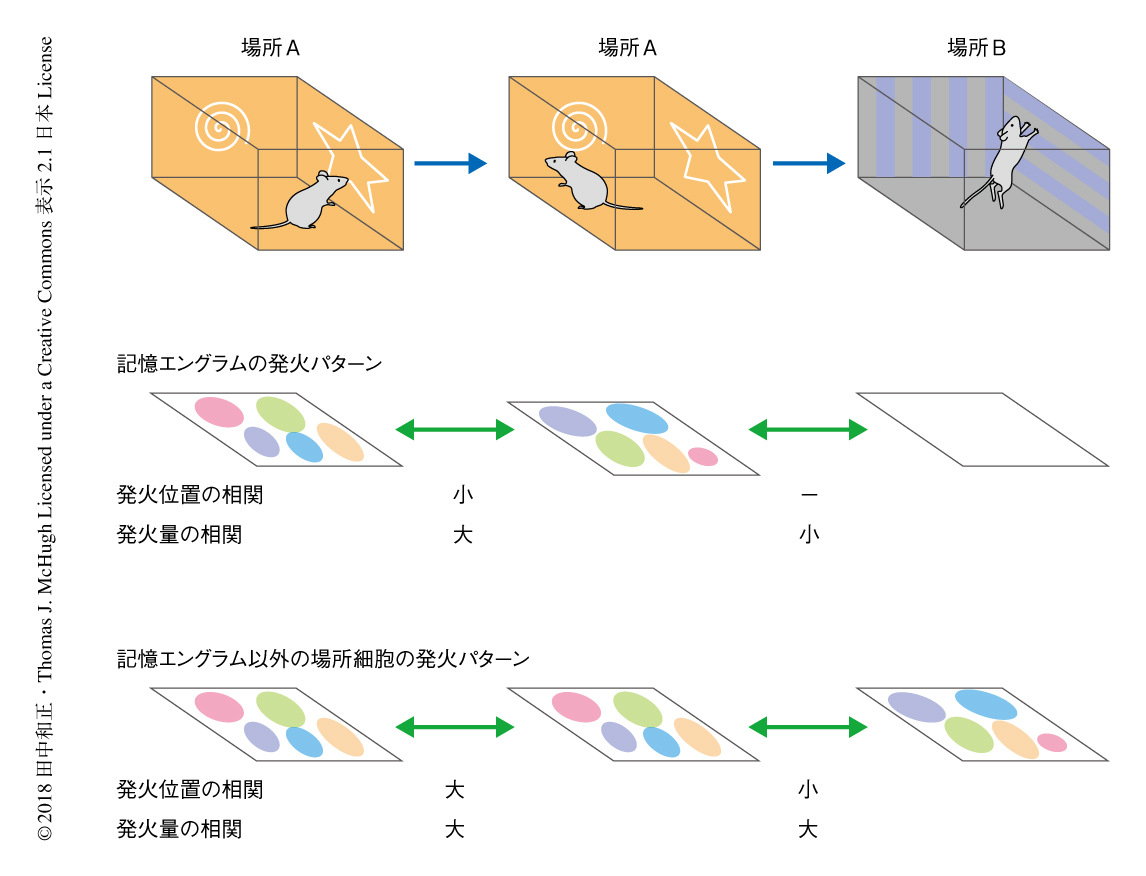
マウスが文脈記憶を想起している際の海馬の記憶エングラムの活動について調べた.標識に用いた文脈をマウスがもういちど探索しているときの記憶エングラムによる空間地図の安定性について評価したところ,空間地図の記憶の痕跡として予想された結果とは真逆に,同じ文脈をもういちど探索したときの記憶エングラムの発火受容野は新しいものへと変化していた.記憶エングラムにおける発火率のマップについて,記銘の際と想起の際の相関は平均してほぼゼロであり,記憶エングラム以外の場所細胞における相関よりも有意に低かった.しかしながら,発火の位置を考慮にいれず平均の発火率だけをみると,記憶エングラムとそれ以外の場所細胞において,記銘の際と想起の際とは同じ程度に相関していた.このことから,記憶エングラムがマウスの空間的な位置以外の文脈の情報を表現することが示唆された.
別の文脈Bを探索する際の記憶エングラムの活動について調べた.マウスが文脈Bを探索しているとき,多くの記憶エングラムはほとんど発火しなかった.結果として,文脈Bの探索の際に場所細胞として位置に特異的な発火を示す記憶エングラムAの割合は有意に低く,また,記憶エングラムAが文脈Bで示す空間の情報量は文脈Aで示す空間の情報量よりも有意に小さかった.それに対し,記憶エングラム以外の場所細胞が示す空間の情報量は文脈Aと文脈Bとで有意差はなかった.ただし,これらの記憶エングラム以外の場所細胞は,発火受容野をシフトさせることにより別の文脈を表現していた.これらの結果から,海馬の記憶エングラムは必ずしも外部の世界についての信頼度の高い空間情報を表現する必要はなく,むしろ,その活動量が海馬以外の別の部位に保存されたエピソード記憶を想起するためのインデックスとして機能するという予測が支持された(図2).
この見方をさらに検証するため,記憶エングラムの部分集合による発火率の相関だけから弁別の指標を定義し,マウスが探索している文脈の弁別を試みた.その結果,ニューロンの集団のレベルでは,記憶エングラムの平均の発火率から高い信頼度をもって文脈Aと文脈Bとが弁別された.それに対し,記憶エングラム以外の場所細胞の平均の発火率により文脈Aと文脈Bは弁別されなかった.さらに,記憶エングラムの平均の発火率による弁別能の高さは,文脈記憶を想起する初期,とくに,最初の数分間に顕著であった.この傾向は,記憶エングラム以外の場所細胞ではみられなかった.このことから,記憶エングラムによる文脈表象はきわめてすばやく再活性化されることが示唆された.これらの結果から,記憶エングラムは場所細胞としては不安定かつ不正確ではあるが,その活動をとおしすばやくかつ高い信頼度をもって文脈のアイデンティティを表現するといえた.
この研究の結果により,海馬の記憶エングラムが空間記憶を保存する記憶の痕跡であるとする見方は否定された.なぜならば,これらのニューロンは同じ環境という同一の入力に対し同じ発火受容野という同一の出力を維持していないからである.筆者らは,記憶エングラムとはその活動量をとおして文脈記憶のインデックスとして機能するニューロンであると提唱する.これは,海馬には2種類の異なる記憶の痕跡が混在することを意味する.すなわち,空間を記憶する素子としてすぐれた記憶エングラム以外の場所細胞が認知地図として外部の世界の枠組みを提供し,空間情報が不安定な記憶エングラムが文脈記憶のインデックスとして皮質の表象へのリンクになるという見方である.
今後は,c-Fosの発現により定義される記憶エングラムにより統一的にエピソード記憶を説明しようとするのではなく,さまざまな様式およびレイヤーにおける可塑的な変化が神経活動をどのように修飾し,結果として何の記憶をどのように担うのかを包括的に明らかにする必要がある.
略歴:2015年 米国California大学Davis校Ph.D.プログラム 修了,同年より理化学研究所脳科学総合研究センター(現 理化学研究所脳神経科学研究センター) 研究員.
研究テーマ:記憶そのものおよび記憶の機構にみられる動的なふるまい.
抱負:Do not work hard, work smart.
Thomas J. McHugh
理化学研究所脳神経科学研究センター チームリーダー.
研究室URL:https://cbs.riken.jp/jp/faculty/t.mchugh/
© 2018 田中和正・Thomas J. McHugh Licensed under CC 表示 2.1 日本
(理化学研究所脳神経科学研究センター 神経回路・行動生理学研究チーム)
email:田中和正
DOI: 10.7875/first.author.2018.076
The hippocampal engram maps experience but not place.
Kazumasa Z. Tanaka, Hongshen He, Anupratap Tomar, Kazue Niisato, Arthur J. Y. Huang, Thomas J. McHugh
Science, 361, 392-397 (2018)
要 約
エピソード記憶は海馬のニューロンが担う.マウスにおいて,記憶エングラムの光遺伝学的な操作により海馬のニューロンが記憶の想起に必要かつ誘導的であることが示されてきた.しかしながら,自由行動下のマウスにおける記憶エングラムの活動のパターンや,記憶における具体的な役割についてはまったくの未知である.この研究において,筆者らは,文脈記憶の記銘の際に海馬CA1野における場所細胞の一部が記憶エングラムになることを見い出した.これらの場所細胞は,記銘の際にシータ域の周波数にて特徴的なバースト活動をくり返していた.この活動のパターンはシナプスを増強させるのに効果的であることが知られている.さらに,記憶を想起する際の記憶エングラムの活動を解析したところ,予想とは逆に,記銘の際に表現していた空間の情報はまったく保持していなかった.しかしながら,記憶エングラムの活動量それ自体は,経験している文脈のアイデンティティをすばやく正確に反映していた.これらの結果から,海馬には異なる2つの記憶の痕跡が形成され,一方は正確な空間の情報を,もう一方はより広義な文脈の情報を保存することが示された.
はじめに
海馬はエピソード記憶を担う.海馬を損傷した患者は日々経験する出来事を記憶すること(記憶の記銘)ができなくなり,また,損傷のまえに経験した事柄の多くを思い出すこと(記憶の想起)ができなくなる.この障害はヒト以外の多くの哺乳類においても起こることが実験的に示されている.
では,海馬はどのようにエピソード記憶を担うのであろうか? この問いは2つの意味をもつ.第1に,記憶の物理的な痕跡とは何かという問い.第2に,海馬が記憶の情報をどのように表現し,その情報が記憶の痕跡によりどのように修飾されることが記憶の記銘に対応するのかという問いである.第1の問いに対しては,シナプス可塑性,とくに,長期増強がその候補であると考えられており,現代の神経科学はこの見方を支持する証拠を積み上げてきた1).たとえば,シナプス可塑性のマーカーである最初期遺伝子の発現により定義されるニューロン,すなわち,記憶エングラムのスパイク活動がある種の記憶の想起に必要かつ誘導的であることが実験的に示されてきた2,3).また,光遺伝学的にスパインの収縮をひき起こすことにより,学習により増強されたスパインが運動の記憶の保持に必要であることも実証された4).しかしながら,それにつづく問い,すなわち,経験により形成された記憶の物理的な痕跡が神経活動,とくに,海馬が表現する情報をどのように安定化(あるいは,変化)させることにより記憶の情報の保存にいたるのか,という問題に対し,その理解はほとんど進んでいない.この問題を説明しようとする理論は数多く存在するものの,それら理論のあいだの整合性が実験的に検証されてきたとはいいがたく,ひとつひとつの理論を直接的に証明することも技術的にむずかしい.
海馬の役割について現在もっとも広く受け入れられている考え方は認知地図仮説であろう5).げっ歯類を含む哺乳類の海馬には,空間の特定の位置に対し発火する場所細胞が存在する.ある環境にいる動物が移動するとき,場所細胞は動物がある位置を通過したときのみ発火する.ひとつひとつの場所細胞は異なる場所受容野をもつため,場所細胞の集団は総体としてひとつの空間地図とみなすことができる.この空間地図は環境に特異的であり,同じ動物が別の環境を移動する際には,異なる組合せの場所細胞の集団が異なる場所受容野をもつことにより別の空間地図を表現する.この見方を拡張し,任意の認知領域における枠組みを表現するのが海馬の役割であると考えるのが認知地図仮説である(図1).では,記憶の物理的な痕跡,すなわち,シナプス可塑性はこうした情報の表現をどのように修飾するのであろうか? もっとも一般的な見方は,シナプス可塑性が位置情報の表現の安定化に寄与するというものである.実際に,シナプス可塑性に重要な役割をはたすNMDA型グルタミン酸受容体を薬理学的あるいは遺伝学的に阻害することにより,場所細胞の安定性は損なわれることが報告されている1).
しかしながら,認知地図仮説は海馬の役割を説明する唯一の理論ではない.なかでも,記憶インデックス仮説は認知地図仮説とは大きく性質が異なる6).記憶インデックス仮説は,海馬に保存される記憶の痕跡は記憶をよび起こすためのたんなるインデックス(索引)にすぎず,記憶の本体は皮質に保存されると提唱する.動物が経験する文脈を構成する刺激の組合せは海馬に記憶の痕跡を形成する.この海馬における記憶の痕跡は,皮質に保存されたそれぞれの刺激の表象とリンクしており,記憶の想起の際にはこれら皮質の表象を再活性化する.すなわち,海馬にはそれ単独で解読の可能な情報が保存されているわけではなく,海馬の役割はあくまで皮質の活動のパターンを再構成することであると考える(図1).
場所細胞についての知見は認知地図仮説を支持するが,文脈恐怖条件づけを用いた研究との整合性が高いのは記憶インデックス仮説である.なぜならば,海馬において文脈表象を形成するためには複数のモダリティからなる刺激を同時に経験する必要があり,文脈記憶を想起するために環境を物理的に探索する必要がないためである7).こうした海馬の文脈表象はしばしば最初期遺伝子の産物であるc-Fosの発現により定義されており,これらを標識あるいは操作することにより文脈記憶の想起が抑制あるいは惹起されることも示されている2,3).
以上のように,電気生理学的な手法を用いた場所細胞の研究と,文脈恐怖条件づけを用いた文脈表象の研究は,海馬の記憶の研究における二大パラダイムを構築してきたといえる.しかしながら,両者を統一的に説明する試みはいまだ成功をおさめていない.
1.海馬CA1野において場所細胞の一部がc-Fosを発現する
このような背景のもと,エピソード記憶に対する海馬の役割をより深く理解するため,海馬の記憶エングラムからテトロード電極を用いた記録を行った.海馬CA1野においてc-Fosを発現するニューロンに光駆動性の陽イオンチャネルであるチャネルロドプシン2および蛍光タンパク質EYFPを発現するマウスを用い,海馬CA1野に光ファイバーおよびテトロード電極を搭載するマイクロドライブを埋め込んだ.このマウスに新規の文脈Aを探索させる(記銘)ことにより,c-Fosを発現する記憶エングラムをチャネルロドプシン2により標識した.翌日,マウスにもういちど文脈Aを探索させ(想起),場所細胞の活動の一貫性から海馬の認知地図の安定性を評価した.さらに12時間後,マイクロドライブに搭載した光ファイバーをとおして海馬CA1野を青色光により刺激し,最初の新規の文脈の探索の結果としてチャネルロドプシン2により標識されたニューロンを同定した.この光遺伝学的な同定により平均19.59%の海馬CA1野のニューロンが光応答性の発火を示し,したがって,c-Fos陽性細胞と同定された.この割合は過去の文献において報告された割合とも一致し8),また,記録ののち確認されたEYFP陽性細胞の割合とも近似していた.最後に,同じマウスに異なる文脈Bを探索させ,文脈に特異的な活動について調べた.
マウスが新規の文脈を探索する際に,記憶エングラムがマウスの位置に応答して発火するかどうか調べた.以前から想定されていたように,記憶エングラムの大部分は記銘した文脈において場所受容野をもっていた.しかしながら,記憶エングラムである場所細胞は全体のごく一部であり,記銘した文脈に応答した場所細胞の大半はチャネルロドプシン2により標識されていなかった.記憶エングラムとそれ以外の場所細胞は生理学的な性質の多くがよく似ており,たとえば,最大の発火率には有意な差はみられなかった.しかしながら,平均の発火率は記憶エングラムのほうが高かった.したがって,記憶エングラムの場所受容野はそれ以外の場所細胞よりも有意に大きく,その結果として,記憶エングラムによる発火ひとつひとつのもつ空間の情報量は小さかった.
2.記憶エングラムの形成の際にみられる特徴的なバースト活動
文脈記憶を記銘する際の発火の時間的な構造を解析した.記憶エングラムの発火時間の間隔について解析したところ,記憶エングラムの発火はそれ以外の場所細胞よりもバーストして起こりやすいことがわかった.記憶エングラムはそれ以外の場所細胞よりもバースト発火率が有意に高く,バーストのあいだの時間間隔が有意に短かった.記憶エングラムのバースト活動の多くは約125ミリ秒の間隔で起こった.これは,海馬において顕著に観察される8 Hzのシータリズムに対応した.結果として,記憶エングラムのバースト活動はそれ以外の場所細胞よりも強くシータ変調されていた.この活動パターンをより定量的に扱うため,83~167ミリ秒(6~12 Hz)の間隔で連続して起こるバースト活動をシータバーストイベントと定義し,記憶エングラムとそれ以外の場所細胞の活動のパターンをさらにくわしく調べた.その結果,新規の文脈の探索において,記憶エングラムにおいてはそれ以外の場所細胞よりも有意に高い率でシータバーストイベントが起こっており,1回のシータバーストイベントにおいて連続して起こるバースト活動の数も有意に多かった.文脈Aの記銘において形成された記憶エングラムAにみられたこれら特徴的な活動のパターンは,マウスが文脈Bを探索する際には記憶エングラムAにおいてみられなかった.このことから,シータ域の波長にて反復して起こるバースト活動が文脈記憶の記銘に関与することが示唆された.
局所フィールド電位から検出されるニューロンの集団の活動がこれらの発火とどのように関連するのかを検証した.海馬CA1野のニューロンは,シータ波(6~12 Hz),遅いガンマ波(30~50 Hz),速いガンマ波(55~85 Hz)の3つのリズムの影響をうけて発火しており,ガンマ波の強度は海馬CA3野(遅いガンマ波)および嗅内皮質(速いガンマ波)からの入力の強度と相関する9).マウスが新規の文脈を探索する際,記憶エングラムを含むすべての場所細胞の発火はシータ波と同じ程度に同期しており,また,これまでに報告されていたように,シータ波の上昇相において優先的に発火していた.しかしながら,単独の発火ではなくシータバーストイベントと振動との関連を解析したところ,記憶エングラム以外の場所細胞におけるシータバーストイベントはシータ波の上昇相にとどまったままであったが,記憶エングラムのシータバーストイベントは下降相へとシフトしていた.海馬CA1野への入力がこれらの場所細胞におよぼす影響について調べるため,ガンマイベントの際の発火について解析した.記憶エングラムにおいては,より多くの発火が速いガンマイベントにおいて起こっており,有意に大きな割合の場所細胞が速いガンマ波と位相固定されていた.これらのガンマイベントにおける発火や位相固定における記憶エングラムとそれ以外の場所細胞とでの違いは,遅いガンマ波においては検出されなかった.これらの結果から,記憶エングラムもそれ以外の場所細胞も海馬CA3野からの入力を同じ程度にうけてはいるが,記憶エングラムは嗅内皮質からの入力に応答性が高く,それによりシータ波にともない起こるシータバーストイベントの位相がシフトする可能性が示唆された.
3.記憶エングラムによる空間表象および文脈表象
マウスが文脈記憶を想起している際の海馬の記憶エングラムの活動について調べた.標識に用いた文脈をマウスがもういちど探索しているときの記憶エングラムによる空間地図の安定性について評価したところ,空間地図の記憶の痕跡として予想された結果とは真逆に,同じ文脈をもういちど探索したときの記憶エングラムの発火受容野は新しいものへと変化していた.記憶エングラムにおける発火率のマップについて,記銘の際と想起の際の相関は平均してほぼゼロであり,記憶エングラム以外の場所細胞における相関よりも有意に低かった.しかしながら,発火の位置を考慮にいれず平均の発火率だけをみると,記憶エングラムとそれ以外の場所細胞において,記銘の際と想起の際とは同じ程度に相関していた.このことから,記憶エングラムがマウスの空間的な位置以外の文脈の情報を表現することが示唆された.
別の文脈Bを探索する際の記憶エングラムの活動について調べた.マウスが文脈Bを探索しているとき,多くの記憶エングラムはほとんど発火しなかった.結果として,文脈Bの探索の際に場所細胞として位置に特異的な発火を示す記憶エングラムAの割合は有意に低く,また,記憶エングラムAが文脈Bで示す空間の情報量は文脈Aで示す空間の情報量よりも有意に小さかった.それに対し,記憶エングラム以外の場所細胞が示す空間の情報量は文脈Aと文脈Bとで有意差はなかった.ただし,これらの記憶エングラム以外の場所細胞は,発火受容野をシフトさせることにより別の文脈を表現していた.これらの結果から,海馬の記憶エングラムは必ずしも外部の世界についての信頼度の高い空間情報を表現する必要はなく,むしろ,その活動量が海馬以外の別の部位に保存されたエピソード記憶を想起するためのインデックスとして機能するという予測が支持された(図2).
この見方をさらに検証するため,記憶エングラムの部分集合による発火率の相関だけから弁別の指標を定義し,マウスが探索している文脈の弁別を試みた.その結果,ニューロンの集団のレベルでは,記憶エングラムの平均の発火率から高い信頼度をもって文脈Aと文脈Bとが弁別された.それに対し,記憶エングラム以外の場所細胞の平均の発火率により文脈Aと文脈Bは弁別されなかった.さらに,記憶エングラムの平均の発火率による弁別能の高さは,文脈記憶を想起する初期,とくに,最初の数分間に顕著であった.この傾向は,記憶エングラム以外の場所細胞ではみられなかった.このことから,記憶エングラムによる文脈表象はきわめてすばやく再活性化されることが示唆された.これらの結果から,記憶エングラムは場所細胞としては不安定かつ不正確ではあるが,その活動をとおしすばやくかつ高い信頼度をもって文脈のアイデンティティを表現するといえた.
おわりに
この研究の結果により,海馬の記憶エングラムが空間記憶を保存する記憶の痕跡であるとする見方は否定された.なぜならば,これらのニューロンは同じ環境という同一の入力に対し同じ発火受容野という同一の出力を維持していないからである.筆者らは,記憶エングラムとはその活動量をとおして文脈記憶のインデックスとして機能するニューロンであると提唱する.これは,海馬には2種類の異なる記憶の痕跡が混在することを意味する.すなわち,空間を記憶する素子としてすぐれた記憶エングラム以外の場所細胞が認知地図として外部の世界の枠組みを提供し,空間情報が不安定な記憶エングラムが文脈記憶のインデックスとして皮質の表象へのリンクになるという見方である.
今後は,c-Fosの発現により定義される記憶エングラムにより統一的にエピソード記憶を説明しようとするのではなく,さまざまな様式およびレイヤーにおける可塑的な変化が神経活動をどのように修飾し,結果として何の記憶をどのように担うのかを包括的に明らかにする必要がある.
文 献
- Neves, G., Cooke, S. F. & Bliss, T. V. P.: Synaptic plasticity, memory and the hippocampus: a neural network approach to causality. Nat. Rev. Neurosci., 9, 65-75 (2008)[PubMed]
- Liu, X., Ramirez, S., Pang, P. T. et al.: Optogenetic stimulation of a hippocampal engram activates fear memory recall. Nature, 484, 381-385 (2012)[PubMed]
- Tanaka, K. Z., Pevzner A., Hamidi A. B. et al.: Cortical representations are reinstated by the hippocampus during memory retrieval. Neuron, 84, 347-354 (2014)[PubMed]
- Hayashi-Takagi, A., Yagishita S., Nakamura M. et al.: Labelling and optical erasure of synaptic memory traces in the motor cortex. Nature, 525, 333-338 (2015)[PubMed]
- O'Keefe, J. & Nadel, L .: The Hippocampus as a Cognitive Map. Oxford University Press, Oxford (1978)
- Teyler, T. J. & DiScenna, P.: The hippocampal memory indexing theory. Behav. Neurosci., 100, 147-154 (1986)[PubMed]
- Rudy, J. W. & O'Reilly, R. C.: Conjunctive representations, the hippocampus, and contextual fear conditioning. Cogn. Affect. Behav. Neurosci., 1, 66-82 (2001)[PubMed]
- Tayler, K. K., Tanaka, K. Z., Reijmers, L. G. et al.: Reactivation of neural ensembles during the retrieval of recent and remote memory. Curr. Biol., 23, 99-106 (2013)[PubMed]
- Lisman, J.: The theta/gamma discrete phase code occuring during the hippocampal phase precession may be a more general brain coding scheme. Hippocampus, 15, 913-922 (2005)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2015年 米国California大学Davis校Ph.D.プログラム 修了,同年より理化学研究所脳科学総合研究センター(現 理化学研究所脳神経科学研究センター) 研究員.
研究テーマ:記憶そのものおよび記憶の機構にみられる動的なふるまい.
抱負:Do not work hard, work smart.
Thomas J. McHugh
理化学研究所脳神経科学研究センター チームリーダー.
研究室URL:https://cbs.riken.jp/jp/faculty/t.mchugh/
© 2018 田中和正・Thomas J. McHugh Licensed under CC 表示 2.1 日本
